



Cytotoxicity, Mitochondrial Functionality, and Redox Status of Human Conjunctival Cells after Short and Chronic Exposure to Preservative-Free Bimatoprost 0.03% and 0.01%: An In Vitro Comparative Study
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
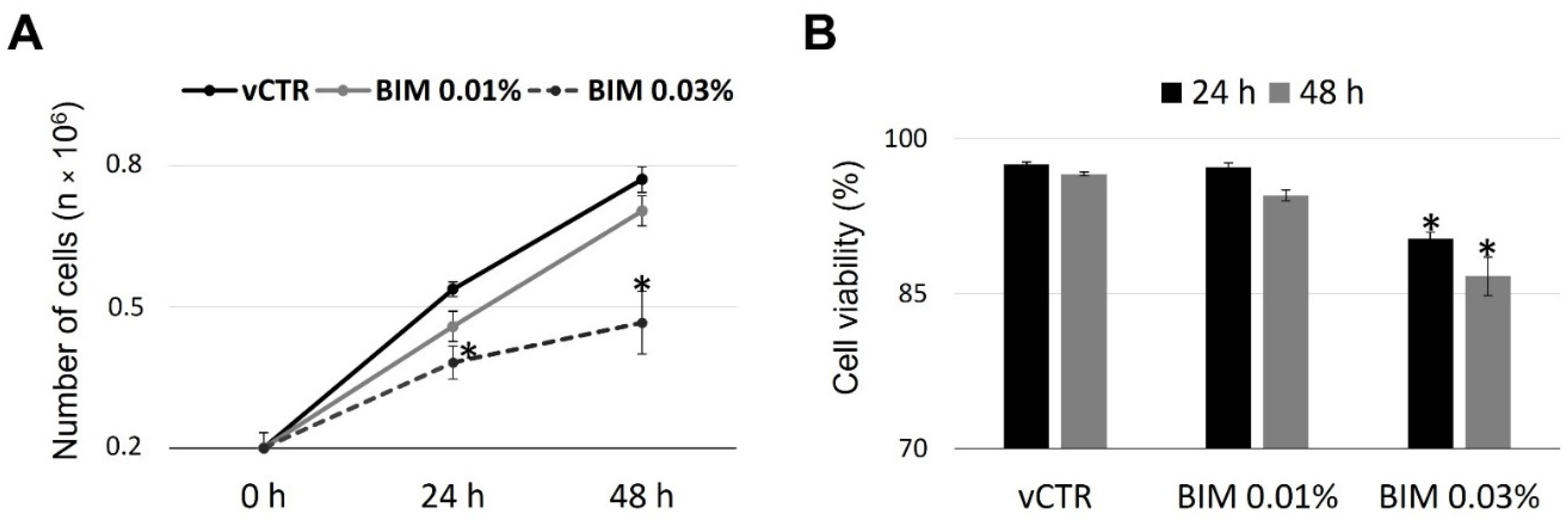
2.1. Cell Growth and Viability of HCE Cells Are Not Affected by BIM 0.01%
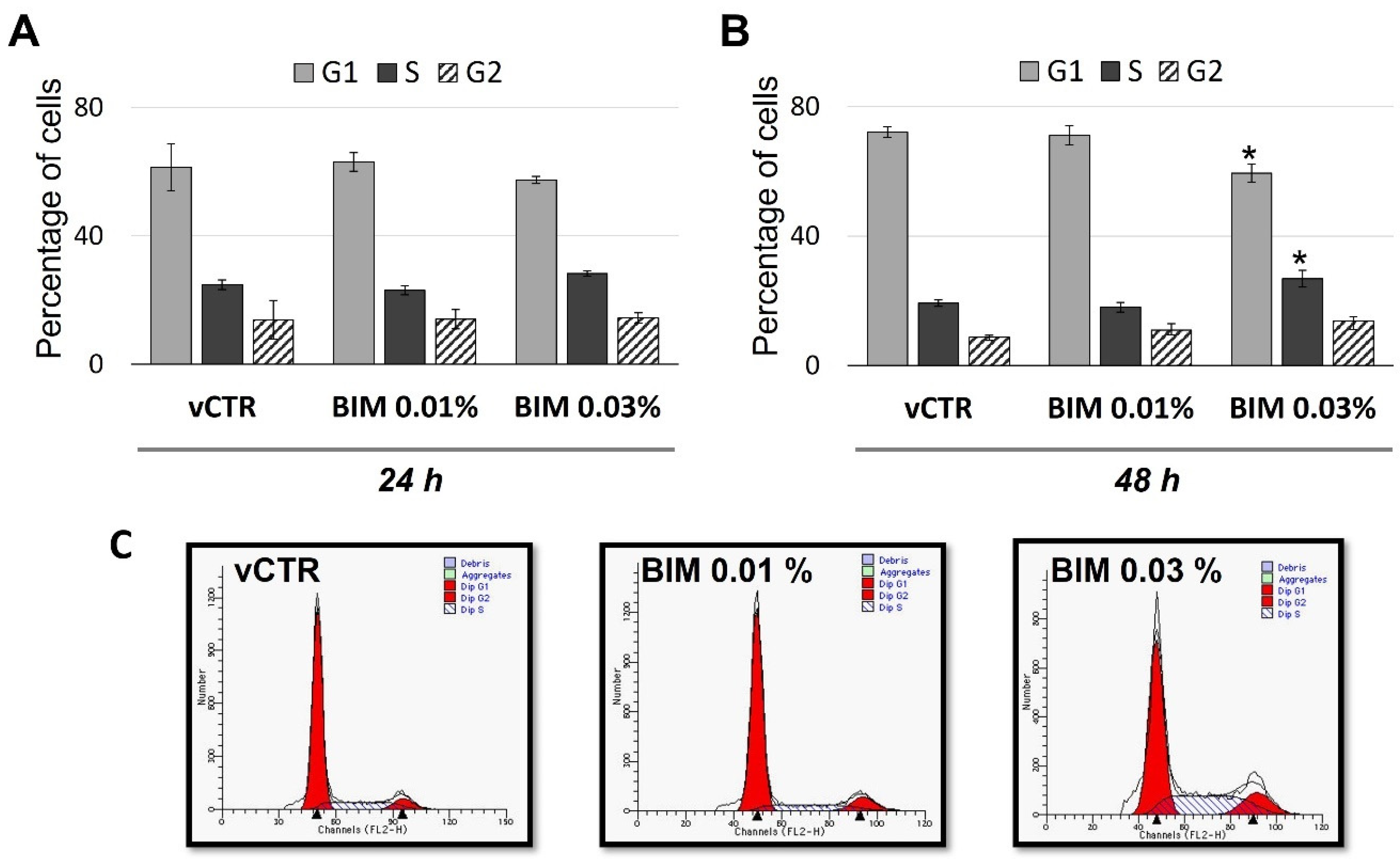
2.2. BIM 0.03% Affects Cell Cycle Phase Distribution, Inducing S/G2 Transition Arrest
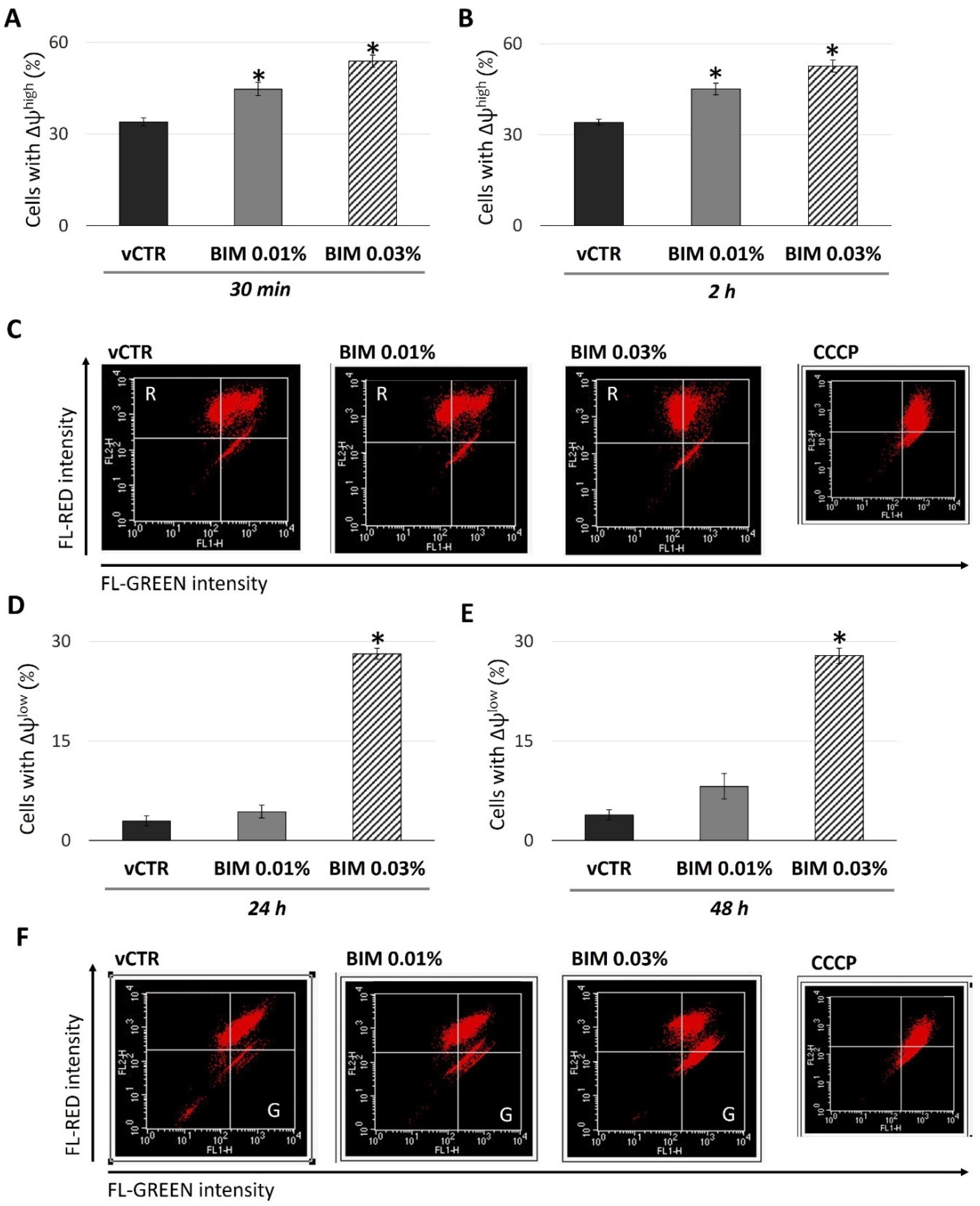
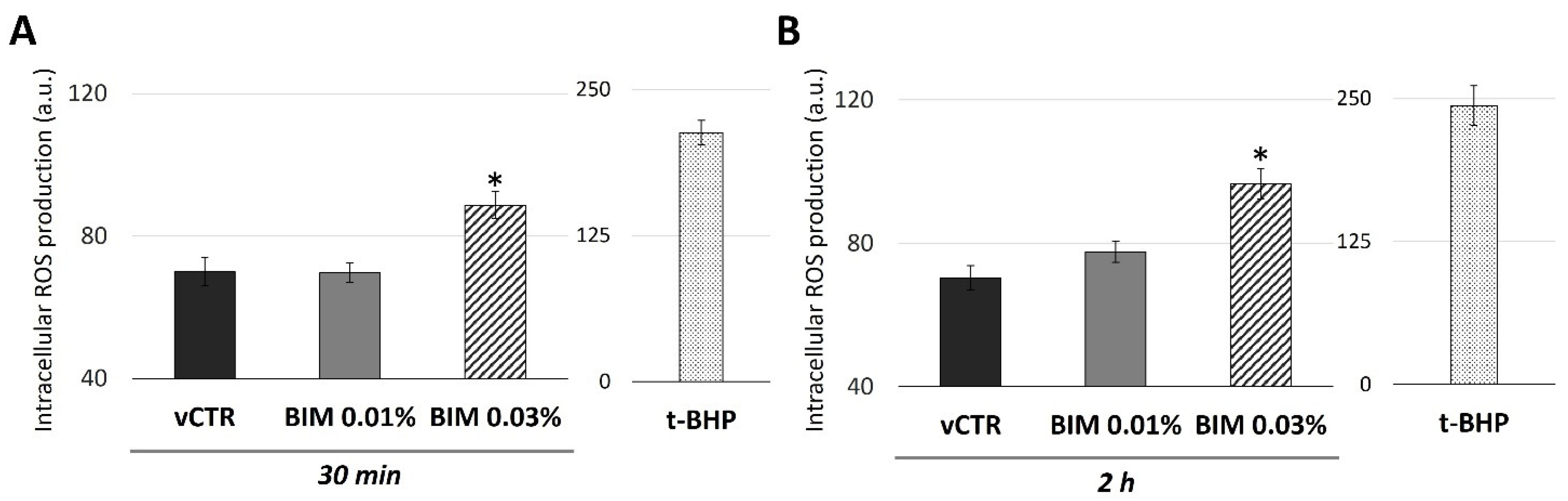
2.3. BIM 0.03% Induces Biphasic Modulation in Mitochondrial Activity and a Short-Term Increase of Intracellular ROS in HCE Cells
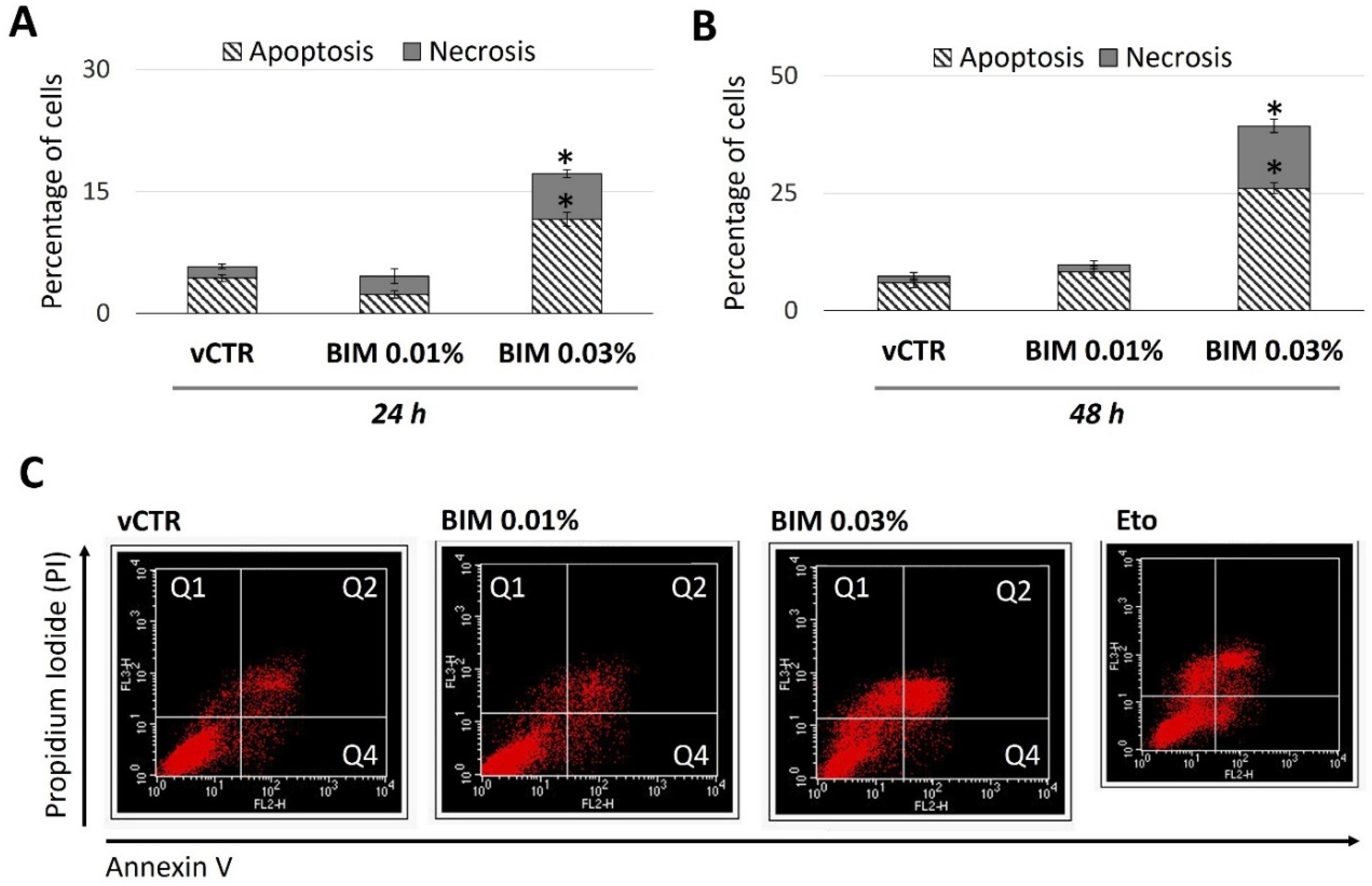
2.4. BIM 0.03% Promotes HCE Cell Death as Both Apoptosis and Necrosis
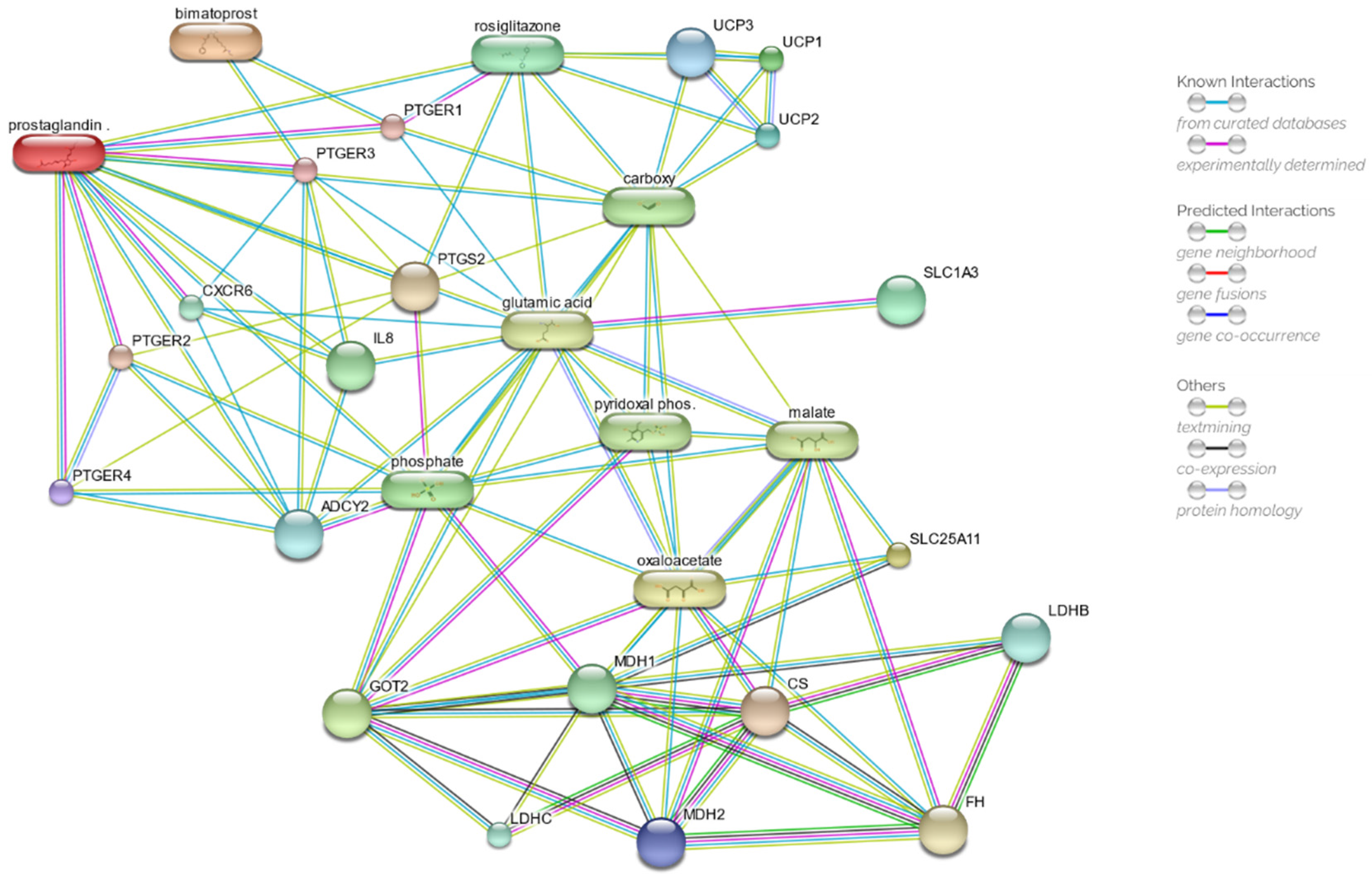
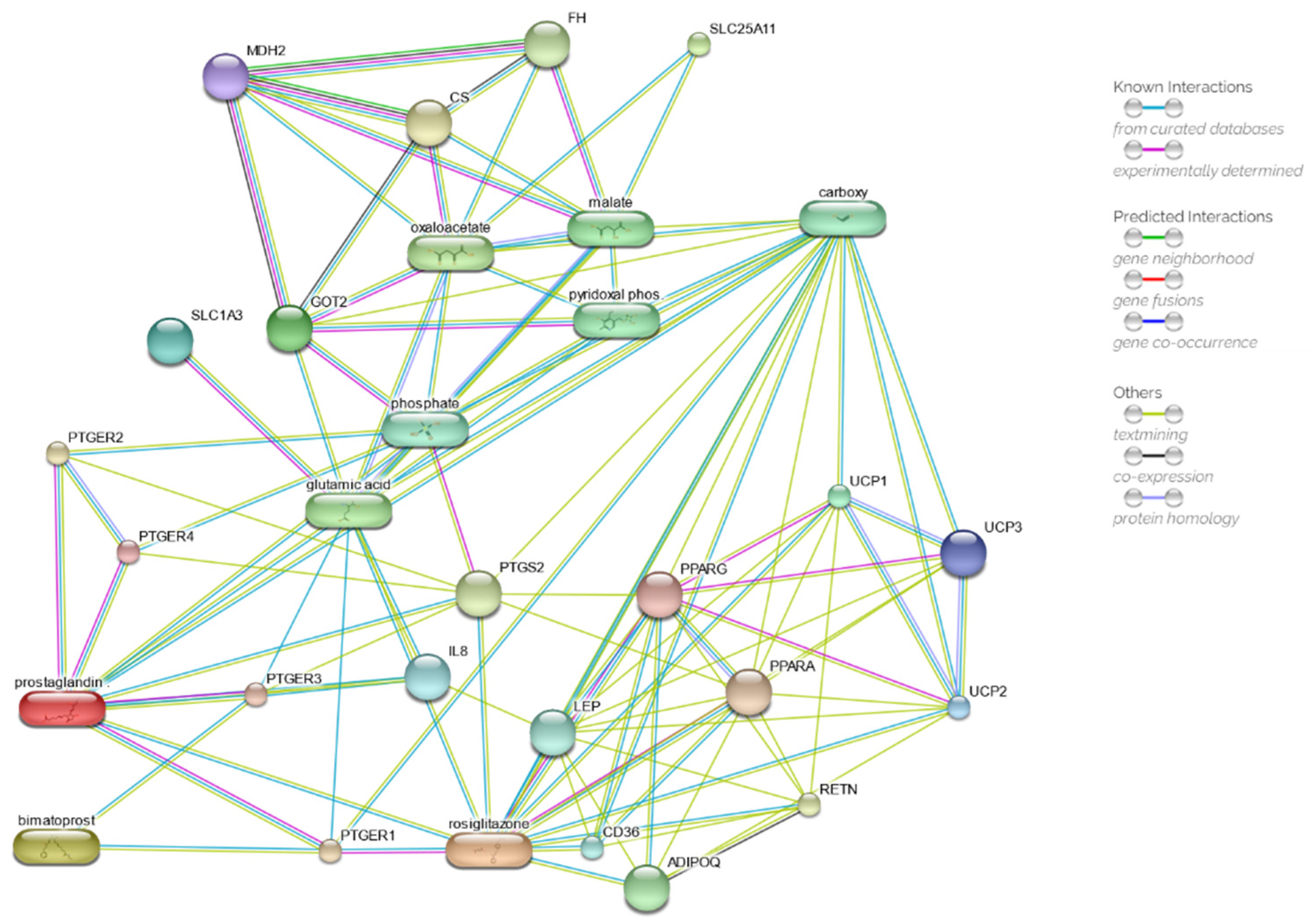
2.5. Drug Network Prediction
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatments
4.2. Cell Growth and Viability
4.3. Flow Cytometry Analyses of Cell Cycle
4.4. Detection of Intracellular Reactive Oxygen Species (ROS)
4.5. Assessment of ΔΨm by Flow Cytometry
4.6. Annexin V/FITC and Propidium Iodide Assay
4.7. Drug Network Prediction
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wu, X.; Yang, X.; Liang, Q.; Xue, X.; Huang, J.; Wang, J.; Xu, Y.; Tong, R.; Liu, M.; Zhou, Q.; et al. Drugs for the treatment of glaucoma: Targets, structure-activity relationships and clinical research. Eur. J. Med. Chem. 2021, 226, 113842. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G. Molecular regulation of neuroinflammation in glaucoma: Current knowledge and the ongoing search for new treatment targets. Prog. Retin. Eye Res. 2022, 87, 100998. [Google Scholar] [CrossRef] [PubMed]
- Terao, E.; Nakakura, S.; Fujisawa, Y.; Nagata, Y.; Ueda, K.; Kobayashi, Y.; Oogi, S.; Dote, S.; Shiraishi, M.; Tabuchi, H.; et al. Time course of conjunctival hyperemia induced by omidenepag isopropyl ophthalmic solution 0.002%: A pilot, comparative study versus ripasudil 0.4%. BMJ Open Ophthalmol. 2020, 5, e000538. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.M. Conjunctival Hyperemia and the Use of Topical Prostaglandins in Glaucoma and Ocular Hypertension. J. Ocul. Pharmacol. Ther. 2003, 19, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Stewart, W.C.; Stewart, J.A.; Jenkins, J.N.; Jackson, A.L. Corneal Punctate Staining With Latanoprost, Bimatoprost, and Travoprost in Healthy Subjects. J. Glaucoma 2003, 12, 475–479. [Google Scholar] [CrossRef]
- Bolívar, G.; Sánchez-Barahona, C.; Teus, M.; Castejón, M.A.; Paz-Moreno-Arrones, J.; Gutiérrez-Ortiz, C.; Mikropoulos, D.G. Effect of topical prostaglandin analogues on corneal hysteresis. Acta Ophthalmol. 2015, 93, e495–e498. [Google Scholar] [CrossRef]
- Kaštelan, S.; Tomić, M.; Metež Soldo, K.; Salopek-Rabatić, J. How ocular surface disease impacts the glaucoma treatment outcome. BioMed Res. Int. 2013, 2013, 696328. [Google Scholar] [CrossRef] [Green Version]
- Baudouin, C. Allergic reaction to topical eyedrops. Curr. Opin. Allergy Clin. Immunol. 2005, 5, 459–463. [Google Scholar] [CrossRef]
- Broadway, D.C.; Bates, A.K.; Lightman, S.L.; Grierson, I.; Hitchings, R.A. The importance of cellular changes in the conjunctiva of patients with uveitic glaucoma undergoing trabeculectomy. Eye 1993, 7, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Netland Nonpenetrating glaucoma surgery. Ophthalmology 2001, 108, 416–421. [CrossRef]
- Arita, R.; Itoh, K.; Maeda, S.; Maeda, K.; Furuta, A.; Tomidokoro, A.; Aihara, M.; Amano, S. Effects of long-term topical anti-glaucoma medications on meibomian glands. Graefes Arch. Clin. Exp. Ophthalmol. 2012, 250, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Arita, R.; Itoh, K.; Maeda, S.; Maeda, K.; Furuta, A.; Tomidokoro, A.; Aihara, M.; Amano, S. Comparison of the long-term effects of various topical antiglaucoma medications on meibomian glands. Cornea 2012, 31, 1229–1234. [Google Scholar] [CrossRef] [PubMed]
- Higginbotham, E.J.; Diestelhorst, M.; Pfeiffer, N.; Rouland, J.F.; Alm, A. The efficacy and safety of unfixed and fixed combinations of latanoprost and other antiglaucoma medications. Surv. Ophthalmol. 2002, 47 (Suppl. S1), S133–S140. [Google Scholar] [CrossRef]
- Cohen, J.S.; Gross, R.L.; Cheetham, J.K.; VanDenburgh, A.M.; Bernstein, P.; Whitcup, S.M. Two-year double-masked comparison of bimatoprost with timolol in patients with glaucoma or ocular hypertension. Surv. Ophthalmol. 2004, 49, S45–S52. [Google Scholar] [CrossRef]
- Williams, R.D.; Cohen, J.S.; Gross, R.L.; Liu, C.-C.; Safyan, E.; Batoosingh, A.L. Bimatoprost Study Group Long-term efficacy and safety of bimatoprost for intraocular pressure lowering in glaucoma and ocular hypertension: Year 4. Br. J. Ophthalmol. 2008, 92, 1387–1392. [Google Scholar] [CrossRef]
- How, A.C.S.; Kumar, R.S.; Chen, Y.-M.; Su, D.H.; Gao, H.; Oen, F.T.; Ho, C.-L.; Seah, S.K.; Aung, T. A randomised crossover study comparing bimatoprost and latanoprost in subjects with primary angle closure glaucoma. Br. J. Ophthalmol. 2009, 93, 782–786. [Google Scholar] [CrossRef]
- Lee, A.J.; McCluskey, P. Clinical utility and differential effects of prostaglandin analogs in the management of raised intraocular pressure and ocular hypertension. Clin. Ophthalmol. 2010, 4, 741–764. [Google Scholar] [CrossRef] [Green Version]
- Jayanetti, V.; Sandhu, S.; Lusthaus, J.A. The Latest Drugs in Development That Reduce Intraocular Pressure in Ocular Hypertension and Glaucoma. J. Exp. Pharmacol. 2020, 12, 539–548. [Google Scholar] [CrossRef]
- Chen, J.; Lu, R.T.; Lai, R.; Dinh, T.; Paul, D.; Venadas, S.; Wheeler, L.A. Bimatoprost-induced calcium signaling in human T-cells does not involve prostanoid FP or TP receptors. Curr. Eye Res. 2009, 34, 184–195. [Google Scholar] [CrossRef]
- Aptel, F.; Cucherat, M.; Denis, P. Efficacy and Tolerability of Prostaglandin Analogs. J. Glaucoma 2008, 17, 667–673. [Google Scholar] [CrossRef]
- Simmons, S.T.; Dirks, M.S.; Noecker, R.J. Bimatoprost versus latanoprost in lowering intraocular pressure in glaucoma and ocular hypertension: Results from parallel-group comparison trials. Adv. Ther. 2004, 21, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Shiokawa, M.; Fujimoto, T.; Tomita, G. Effects of treatment with bimatoprost 0.03% for 3 years in patients with normal-tension glaucoma. Clin. Ophthalmol. 2014, 8, 1179–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsumura, T.; Yoshikawa, K.; Suzumura, H.; Kimura, T.; Sasaki, S.; Kimura, I.; Takeda, R. Bimatoprost ophthalmic solution 0.03% lowered intraocular pressure of normal-tension glaucoma with minimal adverse events. Clin. Ophthalmol. 2012, 6, 1547–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, S.; Hirooka, K.; Baba, T.; Mizote, M.; Fujimura, T.; Tenkumo, K.; Ueda, H.; Shiraga, F. Efficacy and safety of switching from topical latanoprost to bimatoprost in patients with normal-tension glaucoma. J. Ocul. Pharmacol. Ther. 2011, 27, 499–502. [Google Scholar] [CrossRef]
- Chen, M.-J.; Cheng, C.-Y.; Chen, Y.-C.; Chou, C.-K.; Hsu, W.-M. Effects of bimatoprost 0.03% on ocular hemodynamics in normal tension glaucoma. J. Ocul. Pharmacol. Ther. 2006, 22, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Dirks, M.S.; Noecker, R.J.; Earl, M.; Roh, S.; Silverstein, S.M.; Williams, R.D. A 3-month clinical trial comparing the IOP-lowering efficacy of bimatoprost and latanoprost in patients with normal-tension glaucoma. Adv. Ther. 2006, 23, 385–394. [Google Scholar] [CrossRef]
- Servat, J.J.; Bernardino, C.R. Effects of common topical antiglaucoma medications on the ocular surface, eyelids and periorbital tissue. Drugs Aging 2011, 28, 267–282. [Google Scholar] [CrossRef]
- Cate, H.; Bhattacharya, D.; Clark, A.; Fordham, R.; Holland, R.; Broadway, D.C. Improving adherence to glaucoma medication: A randomised controlled trial of a patient-centred intervention (The Norwich Adherence Glaucoma Study). BMC Ophthalmol. 2014, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Filippelli, M.; Campagna, G.; Ciampa, N.; Fioretto, G.; Giannini, R.; Marino, P.F.; dell’Omo, R.; Costagliola, C. Ocular Tolerability of Bimatoprost 0.1 mg/mL Preservative-Free versus Bimatoprost 0.1 mg/mL with Benzalkonium Chloride or Bimatoprost 0.3 mg/mL Preservative-Free in Patients with Primary Open-Angle Glaucoma. J. Clin. Med. 2022, 11, 3518. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamada, M.; Teshima, M.; Nakashima, M.; To, H.; Ichikawa, N.; Sasaki, H. Electrophysiological Characterization of Tight Junctional Pathway of Rabbit Cornea Treated with Ophthalmic Ingredients. Biol. Pharm. Bull. 2007, 30, 2360–2364. [Google Scholar] [CrossRef]
- Figus, M.; Nardi, M.; Piaggi, P.; Sartini, M.; Guidi, G.; Martini, L.; Lazzeri, S. Bimatoprost 0.01% vs bimatoprost 0.03%: A 12-month prospective trial of clinical and in vivo confocal microscopy in glaucoma patients. Eye 2014, 28, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Nixon, D.R.; Simonyi, S.; Bhogal, M.; Sigouin, C.S.; Crichton, A.C.; Discepola, M.; Hutnik, C.M.; Yan, D.B. An observational study of bimatoprost 0.01% in treatment-native patients with primary open angle glaucoma or ocular hypertension: The CLEAR trial. Clin. Ophthalmol. 2012, 6, 2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, M.H.; Silva, F.Q.; Blender, N.; Tran, T.; Vantipalli, S. Ocular benzalkonium chloride exposure: Problems and solutions. Eye 2022, 36, 361–368. [Google Scholar] [CrossRef] [PubMed]
- D’andrea, L.; Montemagni, M.; Celenza, G.; Iorio, R.; Costagliola, C. Is it time for a moratorium on the use of benzalkonium chloride in eyedrops? Br. J. Clin. Pharmacol. 2022, 88, 3947–3949. [Google Scholar] [CrossRef]
- Brignole-Baudouin, F.; Desbenoit, N.; Hamm, G.; Liang, H.; Both, J.-P.; Brunelle, A.; Fournier, I.; Guerineau, V.; Legouffe, R.; Stauber, J.; et al. A new safety concern for glaucoma treatment demonstrated by mass spectrometry imaging of benzalkonium chloride distribution in the eye, an experimental study in rabbits. PLoS ONE 2012, 7, e50180. [Google Scholar] [CrossRef]
- Desbenoit, N.; Schmitz-Afonso, I.; Baudouin, C.; Laprévote, O.; Touboul, D.; Brignole-Baudouin, F.; Brunelle, A. Localisation and quantification of benzalkonium chloride in eye tissue by TOF-SIMS imaging and liquid chromatography mass spectrometry. Anal. Bioanal. Chem. 2013, 405, 4039–4049. [Google Scholar] [CrossRef]
- Datta, S.; Baudouin, C.; Brignole-Baudouin, F.; Denoyer, A.; Cortopassi, G.A. The Eye Drop Preservative Benzalkonium Chloride Potently Induces Mitochondrial Dysfunction and Preferentially Affects LHON Mutant Cells. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2406–2412. [Google Scholar] [CrossRef]
- Rogov, A.G.; Goleva, T.N.; Sukhanova, E.I.; Epremyan, K.K.; Trendeleva, T.A.; Ovchenkova, A.P.; Aliverdieva, D.A.; Zvyagilskaya, R.A. Mitochondrial Dysfunctions May Be One of the Major Causative Factors Underlying Detrimental Effects of Benzalkonium Chloride. Oxid. Med. Cell. Longev. 2020, 2020, 8956504. [Google Scholar] [CrossRef] [Green Version]
- Russo, R.; Varano, G.P.; Adornetto, A.; Nucci, C.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Retinal ganglion cell death in glaucoma: Exploring the role of neuroinflammation. Eur. J. Pharmacol. 2016, 787, 134–142. [Google Scholar] [CrossRef]
- Coyle, S.; Khan, M.N.; Chemaly, M.; Callaghan, B.; Doyle, C.; Willoughby, C.E.; Atkinson, S.D.; Gregory-Ksander, M.; McGilligan, V. Targeting the NLRP3 Inflammasome in Glaucoma. Biomolecules 2021, 11, 1239. [Google Scholar] [CrossRef]
- Baudouin, C. Ocular Surface and External Filtration Surgery: Mutual Relationships. Dev. Ophthalmol. 2017, 59, 67–79. [Google Scholar] [CrossRef] [PubMed]
- European Glaucoma Society Terminology and Guidelines for Glaucoma, 4th Edition—Chapter 2: Classification and terminology Supported by the EGS Foundation: Part 1: Foreword; Introduction; Glossary; Chapter 2 Classification and Terminology. Br. J. Ophthalmol. 2017, 101, 73–127. [CrossRef] [Green Version]
- Mullard, A. 2017 FDA drug approvals. Nat. Rev. Drug Discov. 2018, 17, 81–85. [Google Scholar] [CrossRef]
- Iorio, R.; Celenza, G.; Petricca, S. Mitophagy: Molecular Mechanisms, New Concepts on Parkin Activation and the Emerging Role of AMPK/ULK1 Axis. Cells 2021, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Iorio, R.; Celenza, G.; Petricca, S. Multi-Target Effects of ß-Caryophyllene and Carnosic Acid at the Crossroads of Mitochondrial Dysfunction and Neurodegeneration: From Oxidative Stress to Microglia-Mediated Neuroinflammation. Antioxidants 2022, 11, 1199. [Google Scholar] [CrossRef]
- Schrier, S.A.; Falk, M.J. Mitochondrial disorders and the eye. Curr. Opin. Ophthalmol. 2011, 22, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, L.; Kumar Singh, M. Mitochondria and Eye. In Mutagenesis [Working Title]; IntechOpen: London, UK, 2021. [Google Scholar]
- Meyer, J.N.; Leung, M.C.K.; Rooney, J.P.; Sendoel, A.; Hengartner, M.O.; Kisby, G.E.; Bess, A.S. Mitochondria as a target of environmental toxicants. Toxicol. Sci. 2013, 134, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.N.; Hartman, J.H.; Mello, D.F. Mitochondrial Toxicity. Toxicol. Sci. 2018, 162, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Iorio, R.; Petricca, S.; Luzi, C.; Bellio, P.; Cristiano, L.; Festuccia, C.; Amicosante, G.; Celenza, G. Lactobacillus sakei Pro-Bio65 Reduces TNF-α Expression and Upregulates GSH Content and Antioxidant Enzymatic Activities in Human Conjunctival Cells. Transl. Vis. Sci. Technol. 2021, 10, 8. [Google Scholar] [CrossRef]
- Iorio, R.; Castellucci, A.; Rossi, G.; Cinque, B.; Cifone, M.G.; Macchiarelli, G.; Cecconi, S. Mancozeb affects mitochondrial activity, redox status and ATP production in mouse granulosa cells. Toxicol. In Vitro 2015, 30, 438–445. [Google Scholar] [CrossRef]
- Petricca, S.; Flati, V.; Celenza, G.; Di Gregorio, J.; Lizzi, A.R.; Luzi, C.; Cristiano, L.; Cinque, B.; Rossi, G.; Festuccia, C.; et al. Tebuconazole and Econazole Act Synergistically in Mediating Mitochondrial Stress, Energy Imbalance, and Sequential Activation of Autophagy and Apoptosis in Mouse Sertoli TM4 Cells: Possible Role of AMPK/ULK1 Axis. Toxicol. Sci. 2019, 169, 209–223. [Google Scholar] [CrossRef]
- Iorio, R.; Delle Monache, S.; Bennato, F.; Di Bartolomeo, C.; Scrimaglio, R.; Cinque, B.; Colonna, R.C. Involvement of mitochondrial activity in mediating ELF-EMF stimulatory effect on human sperm motility. Bioelectromagnetics 2011, 32, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Petricca, S.; Celenza, G.; Luzi, C.; Cinque, B.; Lizzi, A.R.; Franceschini, N.; Festuccia, C.; Iorio, R. Synergistic Activity of Ketoconazole and Miconazole with Prochloraz in Inducing Oxidative Stress, GSH Depletion, Mitochondrial Dysfunction, and Apoptosis in Mouse Sertoli TM4 Cells. Int. J. Mol. Sci. 2022, 23, 5429. [Google Scholar] [CrossRef] [PubMed]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial membrane potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef]
- Horbay, R.; Bilyy, R. Mitochondrial dynamics during cell cycling. Apoptosis 2016, 21, 1327–1335. [Google Scholar] [CrossRef]
- Martínez-Reyes, I.; Diebold, L.P.; Kong, H.; Schieber, M.; Huang, H.; Hensley, C.T.; Mehta, M.M.; Wang, T.; Santos, J.H.; Woychik, R.; et al. TCA Cycle and Mitochondrial Membrane Potential Are Necessary for Diverse Biological Functions. Mol. Cell 2016, 61, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Sanin, D.E.; Matsushita, M.; Klein Geltink, R.I.; Grzes, K.M.; van Teijlingen Bakker, N.; Corrado, M.; Kabat, A.M.; Buck, M.D.; Qiu, J.; Lawless, S.J.; et al. Mitochondrial Membrane Potential Regulates Nuclear Gene Expression in Macrophages Exposed to Prostaglandin E2. Immunity 2018, 49, 1021–1033.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newmeyer, D.D.; Ferguson-Miller, S. Mitochondria: Releasing power for life and unleashing the machineries of death. Cell 2003, 112, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Perl, A.; Gergely, P.; Nagy, G.; Koncz, A.; Banki, K. Mitochondrial hyperpolarisation: A checkpoint of T-cell life, death and autoimmunity. Trends Immunol. 2004, 25, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Banki, K.; Hutter, E.; Gonchoroff, N.J.; Perl, A. Elevation of mitochondrial transmembrane potential and reactive oxygen intermediate levels are early events and occur independently from activation of caspases in Fas signaling. J. Immunol. 1999, 162, 1466–1479. [Google Scholar]
- Leal, A.M.d.S.; de Queiroz, J.D.F.; de Medeiros, S.R.B.; Lima, T.K.D.S.; Agnez-Lima, L.F. Violacein induces cell death by triggering mitochondrial membrane hyperpolarisation in vitro. BMC Microbiol. 2015, 15, 115. [Google Scholar] [CrossRef] [PubMed]
- Bajić, A.; Spasić, M.; Andjus, P.R.; Savić, D.; Parabucki, A.; Nikolić-Kokić, A.; Spasojević, I. Fluctuating vs. Continuous Exposure to H2O2: The Effects on Mitochondrial Membrane Potential, Intracellular Calcium, and NF-κB in Astroglia. PLoS ONE 2013, 8, e76383. [Google Scholar] [CrossRef] [PubMed]
- Kadenbach, B.; Arnold, S.; Lee, I.; Hüttemann, M. The possible role of cytochrome c oxidase in stress-induced apoptosis and degenerative diseases. Biochim. Biophys. Acta 2004, 1655, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forkink, M.; Manjeri, G.R.; Liemburg-Apers, D.C.; Nibbeling, E.; Blanchard, M.; Wojtala, A.; Smeitink, J.A.M.; Wieckowski, M.R.; Willems, P.H.G.M.; Koopman, W.J.H. Mitochondrial hyperpolarisation during chronic complex I inhibition is sustained by low activity of complex II, III, IV and V. Biochim. Biophys. Acta 2014, 1837, 1247–1256. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.J. Eicosanoid receptors: Targets for the treatment of disrupted intestinal epithelial homeostasis. Eur. J. Pharmacol. 2017, 796, 7–19. [Google Scholar] [CrossRef]
- Sharif, N.A.; Kelly, C.R.; Crider, J.Y.; Williams, G.W.; Xu, S.X. Ocular Hypotensive FP Prostaglandin (PG) Analogs: PG Receptor Subtype Binding Affinities and Selectivities, and Agonist Potencies at FP and Other PG Receptors in Cultured Cells. J. Ocul. Pharmacol. Ther. 2003, 19, 501–515. [Google Scholar] [CrossRef]
- Föller, M.; Kasinathan, R.S.; Duranton, C.; Wieder, T.; Huber, S.M.; Lang, F. PGE2-induced Apoptotic Cell Death in K562 Human Leukaemia Cells. Cell. Physiol. Biochem. 2006, 17, 201–210. [Google Scholar] [CrossRef]
- Park, J.; Cho, H.K.; Moon, J.I. Changes to upper eyelid orbital fat from use of topical bimatoprost, travoprost, and latanoprost. Jpn. J. Ophthalmol. 2011, 55, 22–27. [Google Scholar] [CrossRef]
- Aydin, S.; Işıklıgil, I.; Tekşen, Y.A.; Kir, E. Recovery of orbital fat pad prolapsus and deepening of the lid sulcus from topical bimatoprost therapy: 2 case reports and review of the literature. Cutan. Ocul. Toxicol. 2010, 29, 212–216. [Google Scholar] [CrossRef]
- Filippopoulos, T.; Paula, J.S.; Torun, N.; Hatton, M.P.; Pasquale, L.R.; Grosskreutz, C.L. Periorbital changes associated with topical bimatoprost. Ophthal. Plast. Reconstr. Surg. 2008, 24, 302–307. [Google Scholar] [CrossRef]
- Jbara, D.; Eiger-Moscovich, M.; Didkovsky, E.; Keshet, Y.; Avisar, I. In Vivo Effects of Prostaglandin Analogues Application by Topical Drops or Retrobulbar Injections on the Orbital Fat of a Rat Model. Ocul. Immunol. Inflamm. 2022, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Eftekhari, K.; Vagefi, R.R.; Lee, V.; Hui, J.Z.; Zhu, M.; Dine, K.; Anderson, R.L.; Koeberlein, B.; Sulaimankutty, R.; Shindler, K.S. In Vivo Effects of Retrobulbar Bimatoprost Injection on Orbital Fat. Ophthal. Plast. Reconstr. Surg. 2018, 34, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Dixit, G.; Prabhu, A. The pleiotropic peroxisome proliferator activated receptors: Regulation and therapeutics. Exp. Mol. Pathol. 2022, 124, 104723. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Barak, Y.; Hevener, A.; Olson, P.; Liao, D.; Le, J.; Nelson, M.; Ong, E.; Olefsky, J.M.; Evans, R.M. Adipose-specific peroxisome proliferator-activated receptor γ knockout causes insulin resistance in fat and liver but not in muscle. Proc. Natl. Acad. Sci. USA 2003, 100, 15712. [Google Scholar] [CrossRef] [Green Version]
- Clouzeau, C.; Godefroy, D.; Riancho, L.; Rostène, W.; Baudouin, C.; Brignole-Baudouin, F. Hyperosmolarity potentiates toxic effects of benzalkonium chloride on conjunctival epithelial cells in vitro. Mol. Vis. 2012, 18, 851. [Google Scholar]
- Brisdelli, F.; Bennato, F.; Bozzi, A.; Cinque, B.; Mancini, F.; Iorio, R. ELF-MF attenuates quercetin-induced apoptosis in K562 cells through modulating the expression of Bcl-2 family proteins. Mol. Cell. Biochem. 2014, 397, 33–43. [Google Scholar] [CrossRef]
- Luzi, C.; Brisdelli, F.; Iorio, R.; Bozzi, A.; Carnicelli, V.; Di Giulio, A.; Lizzi, A.R. Apoptotic effects of bovine apo-lactoferrin on HeLa tumor cells. Cell Biochem. Funct. 2017, 35, 33–41. [Google Scholar] [CrossRef]
- Fagnani, L.; Nazzicone, L.; Brisdelli, F.; Giansanti, L.; Battista, S.; Iorio, R.; Petricca, S.; Amicosante, G.; Perilli, M.; Celenza, G.; et al. Cyclic and Acyclic Amine Oxide Alkyl Derivatives as Potential Adjuvants in Antimicrobial Chemotherapy against Methicillin-Resistant Staphylococcus aureus with an MDR Profile. Antibiotics 2021, 10, 952. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petricca, S.; Celenza, G.; Costagliola, C.; Tranfa, F.; Iorio, R. Cytotoxicity, Mitochondrial Functionality, and Redox Status of Human Conjunctival Cells after Short and Chronic Exposure to Preservative-Free Bimatoprost 0.03% and 0.01%: An In Vitro Comparative Study. Int. J. Mol. Sci. 2022, 23, 14113. https://doi.org/10.3390/ijms232214113
Petricca S, Celenza G, Costagliola C, Tranfa F, Iorio R. Cytotoxicity, Mitochondrial Functionality, and Redox Status of Human Conjunctival Cells after Short and Chronic Exposure to Preservative-Free Bimatoprost 0.03% and 0.01%: An In Vitro Comparative Study. International Journal of Molecular Sciences. 2022; 23(22):14113. https://doi.org/10.3390/ijms232214113
Chicago/Turabian StylePetricca, Sabrina, Giuseppe Celenza, Ciro Costagliola, Fausto Tranfa, and Roberto Iorio. 2022. "Cytotoxicity, Mitochondrial Functionality, and Redox Status of Human Conjunctival Cells after Short and Chronic Exposure to Preservative-Free Bimatoprost 0.03% and 0.01%: An In Vitro Comparative Study" International Journal of Molecular Sciences 23, no. 22: 14113. https://doi.org/10.3390/ijms232214113
APA StylePetricca, S., Celenza, G., Costagliola, C., Tranfa, F., & Iorio, R. (2022). Cytotoxicity, Mitochondrial Functionality, and Redox Status of Human Conjunctival Cells after Short and Chronic Exposure to Preservative-Free Bimatoprost 0.03% and 0.01%: An In Vitro Comparative Study. International Journal of Molecular Sciences, 23(22), 14113. https://doi.org/10.3390/ijms232214113