
Oxygenated Water Increases Seizure Threshold in Various Rodent Seizure Models

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of OW in Prenatal Betamethasone Priming with Postnatal NMDA-Triggered Spasm
2.2. Effects of OW on Postnatal NMDA-Triggered Spasms without Prenatal Betamethasone Priming
2.3. Effects of OW in the Repetitive Kainic Acid Injection Model
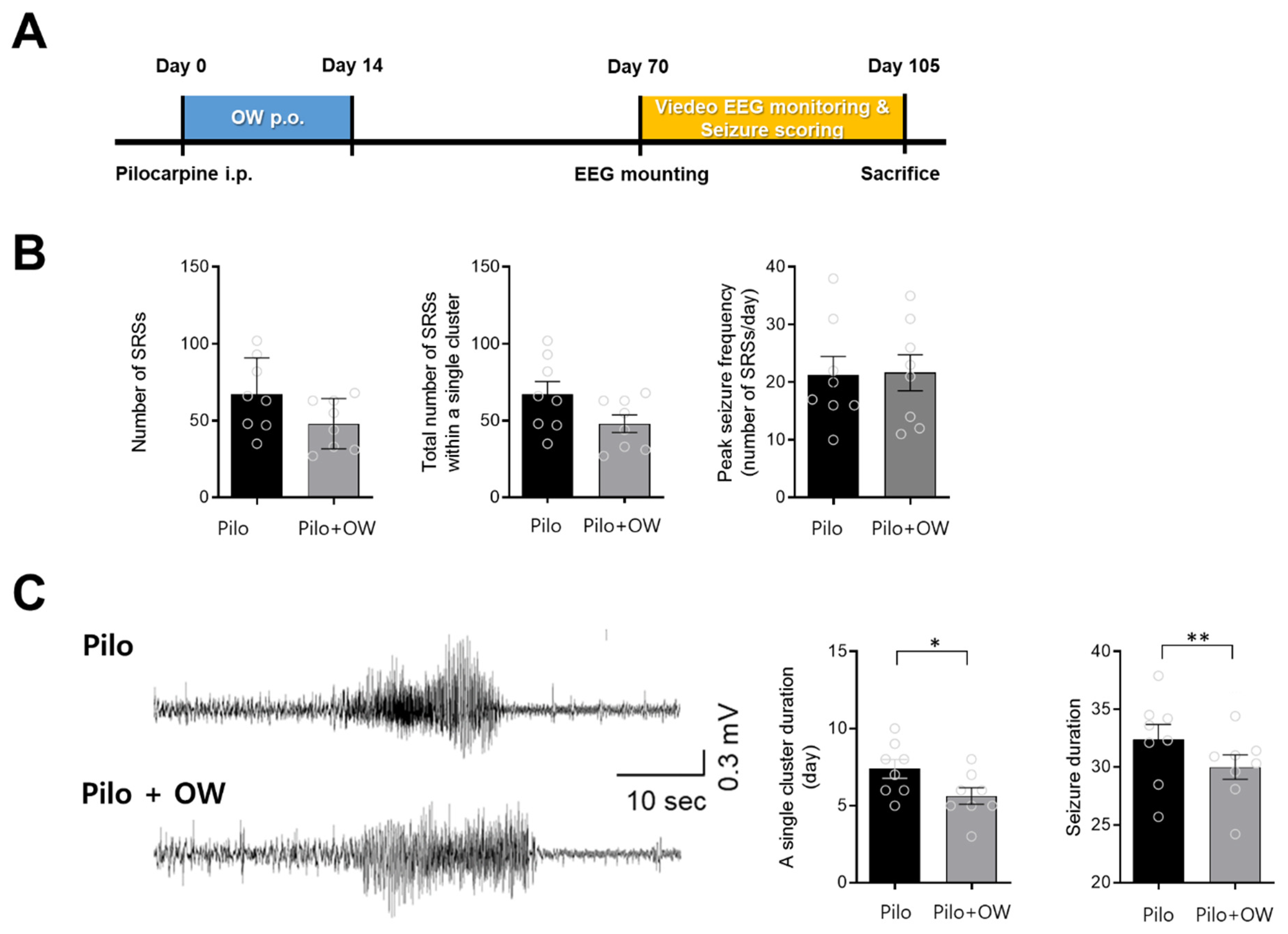
2.4. Effects of OW in the Pilocarpine-Induced Temporal Lobe Epilepsy Model
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.1.1. Prenatal Betamethasone Priming with Postnatal NMDA-Triggered Spasms
4.1.2. Postnatal NMDA-Triggered Spasms without Prenatal Betamethasone Priming
4.1.3. Repetitive Kainate Injection Model
4.1.4. Intraperitoneal Pilocarpine Model
4.2. Cresyl Violet Staining
4.3. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Z.; Brodie, M.J.; Liew, D.; Kwan, P. Treatment outcomes in patients with newly diagnosed epilepsy treated with established and new antiepileptic drugs: A 30-year longitudinal cohort study. JAMA Neurol. 2018, 75, 279–286. [Google Scholar] [CrossRef]
- Pawlik, M.J.; Miziak, B.; Walczak, A.; Konarzewska, A.; Chrościńska-Krawczyk, M.; Albrecht, J.; Czuczwar, S.J. Selected molecular targets for antiepileptogenesis. Int. J. Mol. Sci. 2021, 22, 9737. [Google Scholar] [CrossRef]
- Fang, C.-H.; Tsai, C.-C.; Shyong, Y.-J.; Yang, C.-T.; Li, K.-Y.; Lin, Y.-W.; Chang, K.-C.; Wang, M.-H.; Wu, T.-M.; Lin, F.-H. Effects of highly oxygenated water in a hyperuricemia rat model. J. Healthc. Eng. 2020, 2020, 1323270–1323278. [Google Scholar] [CrossRef]
- Piantadosi, C.A. “Oxygenated” water and athletic performance. Br. J. Sports Med. 2006, 40, 740–741. [Google Scholar] [CrossRef] [Green Version]
- Forth, W.; Adam, O. Uptake of oxygen from the intestine--experiments with rabbits. Eur. J. Med. Res. 2001, 6, 488–492. [Google Scholar]
- Vatnehol, S.A.S.; Hol, P.K.; Bjørnerud, A.; Amiry-Moghaddam, M.; Haglerød, C.; Storås, T.H. Effect of drinking oxygenated water assessed by in vivo MRI relaxometry. J. Magn. Reson. Imaging 2020, 52, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Fuller, P.J. The Effects of Activated Stabilized Oxygen on Aerobic Endurance in Division II Collegiate Male Soccer Players; Humboldt State University: Arcata, CA, USA, 2010. [Google Scholar]
- Jenkins, A.; Moreland, M.R.; Waddell, T.B.; Fernhall, B. Effect of oxygenized water on percent oxygen saturation and performance DURING exercise. Med. Sci. Sports Exer. 2001, 33, S167. [Google Scholar] [CrossRef]
- Fleming, N.; Vaughan, J.; Feeback, M. Ingestion of oxygenated water enhances lactate clearance kinetics in trained runners. J. Int. Soc. Sports Nutr. 2017, 14, 9. [Google Scholar] [CrossRef] [Green Version]
- Wing, S.L.; Askew, E.W.; Luetkemeier, M.J.; Ryujin, D.T.; Kamimori, G.H.; Grissom, C.K. Lack of effect of Rhodiola or oxygenated water supplementation on hypoxemia and oxidative stress. Wilderness Environ. Med. 2003, 14, 9–16. [Google Scholar] [CrossRef]
- Handajani, Y.S.; Tenggara, R.; Suyatna, F.D.; Surjadi, C.; Widjaja, N.T. The effect of oxygenated water in Diabetes Mellitus. Med. J. Indones. 2009, 18, 102–107. [Google Scholar] [CrossRef]
- Huff, J.S.; Fountain, N.B. Pathophysiology and definitions of seizures and status epilepticus. Emerg. Med. Clin. N. Am. 2011, 29, 1–13. [Google Scholar] [CrossRef]
- Ingram, J.; Zhang, C.; Cressman, J.R.; Hazra, A.; Wei, Y.; Koo, Y.E.; Žiburkus, J.; Kopelman, R.; Xu, J.; Schiff, S.J. Oxygen and seizure dynamics: I. Experiments. J. Neurophysiol. 2014, 112, 205–212. [Google Scholar] [CrossRef] [Green Version]
- McGarry, T.; Biniecka, M.; Veale, D.J.; Fearon, U. Hypoxia, oxidative stress and inflammation. Free Radic. Biol. Med. 2018, 125, 15–24. [Google Scholar] [CrossRef]
- Rusina, E.; Bernard, C.; Williamson, A. The Kainic Acid Models of Temporal Lobe Epilepsy Eneuro. Eneuro 2021, 8. [Google Scholar] [CrossRef]
- Tse, K.; Puttachary, S.; Beamer, E.; Sills, G.J.; Thippeswamy, T. Advantages of repeated low dose against single high dose of kainate in C57BL/6J mouse model of status epilepticus: Behavioral and electroencephalographic studies. PLoS ONE 2014, 9, e96622. [Google Scholar] [CrossRef]
- Umpierre, A.D.; Bennett, I.V.; Nebeker, L.D.; Newell, T.G.; Tian, B.B.; Thomson, K.E.; White, H.S.; White, J.A.; Wilcox, K.S. Repeated low-dose kainate administration in C57BL/6J mice produces temporal lobe epilepsy pathology but infrequent spontaneous seizures. Exp. Neurol. 2016, 279, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Houser, C.R. Granule cell dispersion in the dentate gyrus of humans with temporal lobe epilepsy. Brain Res. 1990, 535, 195–204. [Google Scholar] [CrossRef]
- Velíšek, L.; Jehle, K.; Asche, S.; Velíšková, J. Model of infantile spasms induced by N-methyl-D-aspartic acid in prenatally impaired brain. Ann. Neurol. 2007, 61, 109–119. [Google Scholar] [CrossRef]
- Barrett, K.T.; Choudhary, A.; Charkhand, B.; Scantlebury, M.H. Animal models in epileptic spasms and the development of novel treatment options. J. Clin. Neurophysiol. 2022, 10, 1097. [Google Scholar] [CrossRef]
- Shi, X.Y.; Ju, J.; Zou, L.P.; Wang, J.; Shang, N.X.; Zhao, J.B.; Wang, J.; Zhang, J.Y. Increased precipitation of spasms in an animal model of infantile spasms by prenatal stress exposure. Life Sci. 2016, 152, 171–177. [Google Scholar] [CrossRef]
- Song, L.; Zheng, J.; Li, H.; Jia, N.; Suo, Z.; Cai, Q.; Bai, Z.; Cheng, D.; Zhu, Z. Prenatal stress causes oxidative damage to mitochondrial DNA in hippocampus of offspring rats. Neurochem. Res. 2009, 34, 739–745. [Google Scholar] [CrossRef]
- Kwon, H.H.; Lee, J.S.; Park, H.; Shin, J.; Yin, Y.; Shin, N.; Shin, H.J.; Hwang, J.A.; Kim, D.W.; Kang, J.W. Vitamin E reduces spasms caused by prenatal stress by lowering calpain expression. Epilepsy Behav. 2021, 114, 107609. [Google Scholar] [CrossRef]
- Reyes, R.C.; Brennan, A.M.; Shen, Y.; Baldwin, Y.; Swanson, R.A. Activation of neuronal NMDA receptors induces superoxide-mediated oxidative stress in neighboring neurons and astrocytes. J. Neurosci. 2012, 32, 12973–12978. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, C.; Arias-Cavieres, A. Calcium, reactive oxygen species, and synaptic plasticity. Physiology 2016, 31, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Nadler, J.V.; Perry, B.W.; Cotman, C.W. Intraventricular kainic acid preferentially destroys hippocampal pyramidal cells. Nature 1978, 271, 676–677. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, M.; Avoli, M. The kainic acid model of temporal lobe epilepsy. Neurosci. Biobehav. Rev. 2013, 37, 2887–2899. [Google Scholar] [CrossRef] [Green Version]
- Covolan, L.; Mello, L.E. Temporal profile of neuronal injury following pilocarpine or kainic acid-induced status epilepticus. Epilepsy Res. 2000, 39, 133–152. [Google Scholar] [CrossRef]
- Özugur, S.; Kunz, L.; Straka, H. Relationship between oxygen consumption and neuronal activity in a defined neural circuit. BMC Biol. 2020, 18, 76. [Google Scholar] [CrossRef] [PubMed]
- Mukandala, G.; Tynan, R.; Lanigan, S.; O’Connor, J.J. The effects of hypoxia and inflammation on synaptic signaling in the CNS. Brain Sci. 2016, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Lai, U.H.; Zhu, L.; Singh, A.; Ahmed, M.; Forsyth, N.R. Reactive oxygen species formation in the brain at different oxygen levels: The role of hypoxia inducible factors. Front. Cell Dev. Biol. 2018, 6, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovac, S.; Domijan, A.M.; Walker, M.C.; Abramov, A.Y. Seizure activity results in calcium- and mitochondria-independent ROS production via NADPH and xanthine oxidase activation. Cell Death Dis. 2014, 5, e1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perucca, E.; Brodie, M.J.; Kwan, P.; Tomson, T. 30 years of second-generation antiseizure medications: Impact and future perspectives. Lancet Neurol. 2020, 19, 544–556. [Google Scholar] [CrossRef]
- Baek, H.; Yi, M.H.; Pandit, S.; Park, J.B.; Kwon, H.H.; Zhang, E.; Kim, S.; Shin, N.; Kim, E.; Lee, Y.H.; et al. Altered expression of KCC2 in GABAergic interneuron contributes prenatal stress-induced epileptic spasms in infant rat. Neurochem. Int. 2016, 97, 57–64. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, H.H.; Jung, S.Y.; Park, H.; Shin, H.J.; Kim, D.W.; Song, H.-J.; Kang, J.W. Oxygenated Water Increases Seizure Threshold in Various Rodent Seizure Models. Int. J. Mol. Sci. 2022, 23, 14124. https://doi.org/10.3390/ijms232214124
Kwon HH, Jung SY, Park H, Shin HJ, Kim DW, Song H-J, Kang JW. Oxygenated Water Increases Seizure Threshold in Various Rodent Seizure Models. International Journal of Molecular Sciences. 2022; 23(22):14124. https://doi.org/10.3390/ijms232214124
Chicago/Turabian StyleKwon, Hyeok Hee, Seung Yeon Jung, Hyewon Park, Hyo Jung Shin, Dong Woon Kim, Hee-Jung Song, and Joon Won Kang. 2022. "Oxygenated Water Increases Seizure Threshold in Various Rodent Seizure Models" International Journal of Molecular Sciences 23, no. 22: 14124. https://doi.org/10.3390/ijms232214124
APA StyleKwon, H. H., Jung, S. Y., Park, H., Shin, H. J., Kim, D. W., Song, H. -J., & Kang, J. W. (2022). Oxygenated Water Increases Seizure Threshold in Various Rodent Seizure Models. International Journal of Molecular Sciences, 23(22), 14124. https://doi.org/10.3390/ijms232214124