
Does a Little Difference Make a Big Difference? Bovine β-Casein A1 and A2 Variants and Human Health—An Update

 ,
,  ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
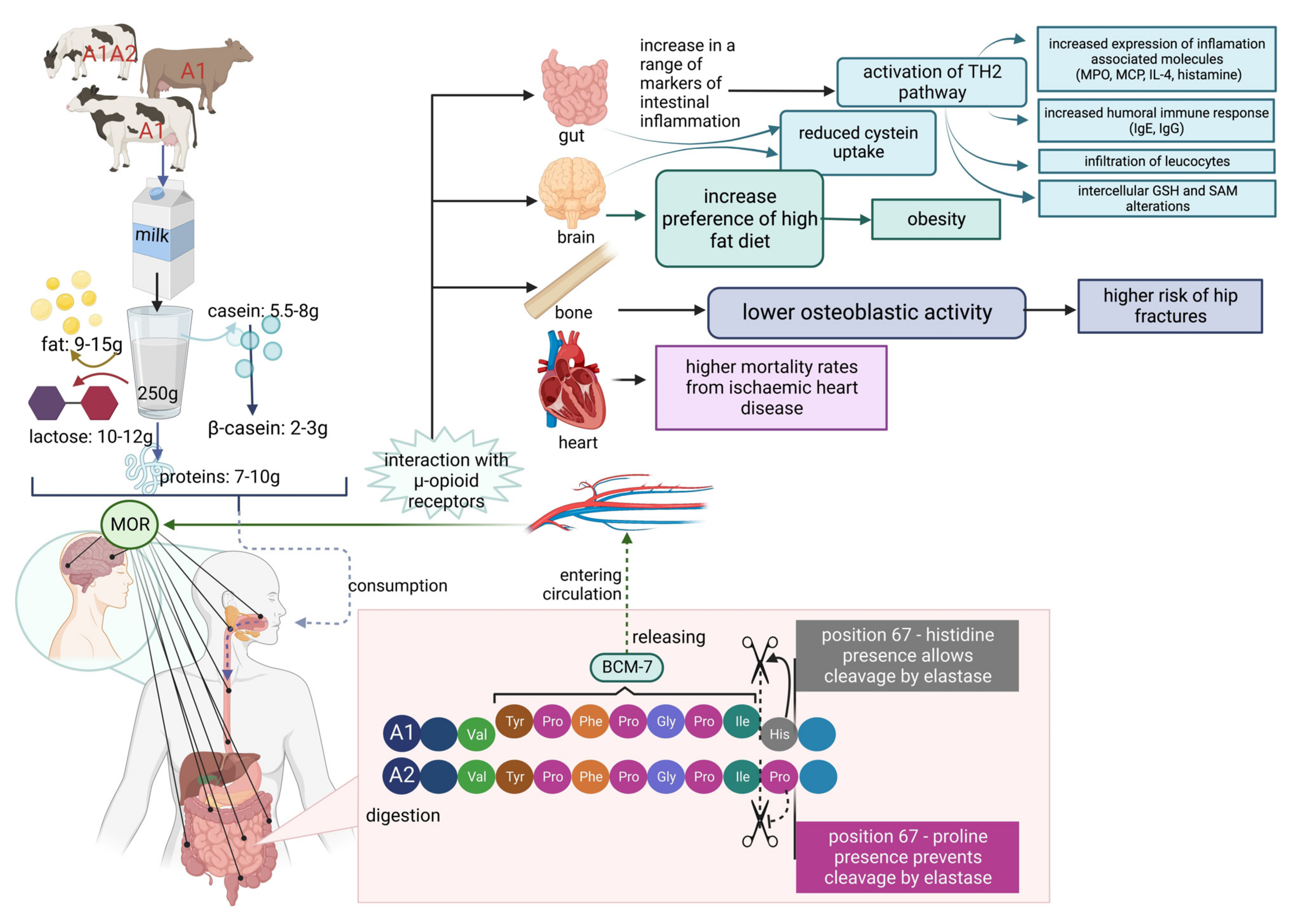{kind=link}
{kind=link}
Abstract
:1. Introduction
2. β-Casein Gene
2.1. Structure and Genetic Variants
2.2. Variants and Their Frequency in Dairy Cattle Breeds
3. Beta-Casein Protein
4. β-Casein Digestion
5. β-Casomorphins
6. β-Casomorphin-7 in Milk and Milk Products
7. β-Casein Variants A1/A2 and Human Health
7.1. Lactose Intolerance
7.2. Inflammatory Response and Gastrointestinal Problems
7.3. Allergy
7.4. Diabetes Mellitus
7.5. Ischemic Heart Disease
7.6. Nervous System
8. β-Casein Variants A1/A2 in Dairy Cattle Breeding
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Daniloski, D.; McCarthy, N.A.; Markoska, T.; Auldist, M.J.; Vasiljevic, T. Conformational and Physicochemical Characteristics of Bovine Skim Milk Obtained from Cows with Different Genetic Variants of β-Casein. Food Hydrocoll. 2022, 124, 107186. [Google Scholar] [CrossRef]
- Kaminski, S.; Cieslinska, A.; Kostyra, E. Polymorphism of Bovine Beta-Casein and Its Potential Effect on Human Health. J. Appl. Genet. 2007, 48, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, M.; Mukesh, M.; Kataria, R.S.; Mishra, B.P.; Joshii, B.K. Milk Proteins and Human Health: A1/A2 Milk Hypothesis. Indian J. Endocrinol. Metab. 2012, 16, 856. [Google Scholar] [CrossRef] [PubMed]
- Thiruvengadam, M.; Venkidasamy, B.; Thirupathi, P.; Chung, I.-M.; Subramanian, U. β-Casomorphin: A Complete Health Perspective. Food Chem. 2021, 337, 127765. [Google Scholar] [CrossRef] [PubMed]
- Swinburn, B. Beta Casein A 1 and A 2 in Milk and Human Health; Report to New Zealand Food Safety Authority; School of Health Sciences: Melbourne, Australia, 2004. [Google Scholar]
- Review of the Potential Health Impact of β-Casomorphins and Related Peptides | EFSA. Available online: https://www.efsa.europa.eu/pl/efsajournal/pub/rn-231 (accessed on 2 October 2022).
- Miller, M.J.; Witherly, S.A.; Clark, D.A. Casein: A Milk Protein with Diverse Biologic Consequences. Proc. Soc. Exp. Biol. Med. 1990, 195, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Ostersen, S.; Foldager, J.; Hermansen, J.E. Effects of Stage of Lactation, Milk Protein Genotype and Body Condition at Calving on Protein Composition and Renneting Properties of Bovine Milk. J. Dairy Res. 1997, 64, 207–219. [Google Scholar] [CrossRef]
- Caroli, A.M.; Chessa, S.; Erhardt, G.J. Invited Review: Milk Protein Polymorphisms in Cattle: Effect on Animal Breeding and Human Nutrition. J. Dairy Sci. 2009, 92, 5335–5352. [Google Scholar] [CrossRef] [Green Version]
- Cieslak, J.; Wodas, L.; Borowska, A.; Pawlak, P.; Czyzak-Runowska, G.; Wojtowski, J.; Puppel, K.; Kuczynska, B.; Mackowski, M. 5’-Flanking Variants of Equine Casein Genes (CSN1S1, CSN1S2, CSN2, CSN3) and Their Relationship with Gene Expression and Milk Composition. J. Appl. Genet. 2019, 60, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Threadgill, D.W.; Womack, J.E. Genomic Analysis of the Major Bovine Milk Protein Genes. Nucleic Acids Res. 1990, 18, 6935–6942. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, L.; Leone, P.; Sgaramella, V. Long Range Restriction Analysis of the Bovine Casein Genes. Nucleic Acids Res. 1990, 18, 6829. [Google Scholar] [CrossRef]
- Rijnkels, M.; Elnitski, L.; Miller, W.; Rosen, J.M. Multispecies Comparative Analysis of a Mammalian-Specific Genomic Domain Encoding Secretory Proteins. Genomics 2003, 82, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Bonsing, J.; Ring, J.M.; Stewart, A.F.; Mackinlay, A.G. Complete Nucleotide Sequence of the Bovine Beta-Casein Gene. Aust. J. Biol. Sci. 1988, 41, 527–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aschaffenburg, R.; Sen, A.; Thompson, M.P. Genetic Variants of Casein in Indian and African Zebu Cattle. Comp. Biochem. Physiol. 1968, 25, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.F.; Kopfler, F.C. Detection of New Types of β-Casein by Polyacrylamide Gel Electrophoresis at Acid PH: A Proposed Nomenclature. Biochem. Biophys. Res. Commun. 1966, 22, 388–392. [Google Scholar] [CrossRef]
- Kiddy, C.A.; Peterson, R.F.; Kopfler, F.C. Genetic control of the variants of β-casein A. J. Dairy Sci. 1966, 49, 742. [Google Scholar]
- Aschaffenburg, R. Imherited Casein Variants in Cow’s Milk. Nature 1961, 192, 431–432. [Google Scholar] [CrossRef]
- Aschaffenburg, R. Inherited Casein Variants in Cow’s Milk: II. Breed Differences in the Occurrence of β-Casein Variants. J. Dairy Res. 1963, 30, 251–258. [Google Scholar] [CrossRef]
- Voglino, G.F. A New β-Casein Variant in Piedmont Cattle. Anim. Blood Groups Biochem. Genet. 1972, 3, 61–62. [Google Scholar] [CrossRef]
- Visser, S.; Slangen, C.J.; Lagerwerf, F.M.; Van Dongen, W.D.; Haverkamp, J. Identification of a New Genetic Variant of Bovine Beta-Casein Using Reversed-Phase High-Performance Liquid Chromatography and Mass Spectrometric Analysis. J. Chromatogr. A 1995, 711, 141–150. [Google Scholar] [CrossRef]
- Dong, C.; Ng-Kwai-Hang, K.F. Characterization of a Non-Electrophoretic Genetic Variant of β-Casein by Peptide Mapping and Mass Spectrometric Analysis. Int. Dairy J. 1998, 8, 967–972. [Google Scholar] [CrossRef]
- Han, S.K.; Shin, Y.C.; Byun, H.D. Biochemical, Molecular and Physiological Characterization of a New β-Casein Variant Detected in Korean Cattle. Anim. Genet. 2000, 31, 49–51. [Google Scholar] [CrossRef] [PubMed]
- Senocq, D.; Mollé, D.; Pochet, S.; Léonil, J.; Dupont, D.; Levieux, D. A New Bovine β-Casein Genetic Variant Characterized by a Met93→Leu93 Substitution in the Sequence A2. Lait 2002, 82, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Jann, O.; Ceriotti, G.; Caroli, A.; Erhardt, G. A New Variant in Exon VII of Bovine β-Casein Gene (CSN2) and Its Distribution among European Cattle Breeds. J. Anim. Breed. Genet. 2002, 119, 65–68. [Google Scholar] [CrossRef]
- Gallinat, J.L.; Qanbari, S.; Drögemüller, C.; Pimentel, E.C.G.; Thaller, G.; Tetens, J. DNA-Based Identification of Novel Bovine Casein Gene Variants. J. Dairy Sci. 2013, 96, 699–709. [Google Scholar] [CrossRef] [Green Version]
- Swaisgood, H.E. Review and Update of Casein Chemistry1, 2. J. Dairy Sci. 1993, 76, 3054–3061. [Google Scholar] [CrossRef]
- Truswell, A.S. The A2 Milk Case: A Critical Review. Eur. J. Clin. Nutr. 2005, 59, 623–631. [Google Scholar] [CrossRef] [Green Version]
- Roginski, H.; Fuquay, J.W.; Fox, P.F. Encyclopedia of Dairy Sciences; Academic Press: Cambridge, MA, USA, 2003; Volume 1–4. [Google Scholar]
- Groves, M.L. Some Minor Components of Casein and Other Phosphoproteins in Milk. A Review. J. Dairy Sci. 1969, 52, 1155–1165. [Google Scholar] [CrossRef]
- Seibert, B.; Erhardt, G.; Senft, B. Procedure for Simultaneous Phenotyping of Genetic Variants in Cow’s Milk by Isoelectric Focusing. Anim. Blood Groups Biochem. Genet. 1985, 16, 183–191. [Google Scholar] [CrossRef]
- Caroli, A.M.; Savino, S.; Bulgari, O.; Monti, E. Detecting β-Casein Variation in Bovine Milk. Molecules 2016, 21, 141. [Google Scholar] [CrossRef] [Green Version]
- Farrell, H.M.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the Proteins of Cows’ Milk--Sixth Revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef] [Green Version]
- Grosclaude, F.; Mahé, M.-F.; Mercier, J.-C. Comparaison Du Polymorphisme Génétique Des Lactoprotéines Du Zébu et Des Bovins. Ann. Genet. Sel. Anim. 1974, 6, 305–329. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, I.; Gaur, G.K.; Singh, U.; Kumar, S.; Kumar, S.; Mann, S. Beta-Casein (CSN2) Polymorphism in Ongole (Indian Zebu) and Frieswal (HF × Sahiwal Crossbred) Cattle. Indian J. Biotechnol. 2013, 12, 195–198. [Google Scholar]
- Rahman, S.; Islam, A.; Alam, M.; Hossain, M.; Alim, M.A.; Salimmullah, M.; Alam, J. In Analysis of Beta-Casein Gene Variants of Milk in Cattle. In Proceedings of the 3rd International Exhibition on Dairy Aqua and Pet, Dhaka, Bangladesh, 18–20 February 2016; pp. 45–50. [Google Scholar]
- Jiménez-Montenegro, L.; Alfonso, L.; Mendizabal, J.A.; Urrutia, O. Worldwide Research Trends on Milk Containing Only A2 β-Casein: A Bibliometric Study. Animals 2022, 12, 1909. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Hosen, M.B.; Faruk, M.O.; Hasan, M.M.; Kabir, Y.; Howlader, M.Z.H. Association of Vitamin D and Vitamin D Binding Protein (DBP) Gene Polymorphism with Susceptibility of Type 2 Diabetes Mellitus in Bangladesh. Gene 2017, 636, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Bech, A.-M.; Kristiansen, K.R. Milk Protein Polymorphism in Danish Dairy Cattle and the Influence of Genetic Variants on Milk Yield. J. Dairy Res. 1990, 57, 53–62. [Google Scholar] [CrossRef]
- Van Eenennaam, A.; Medrano, J.F. Milk Protein Polymorphisms in California Dairy Cattle. J. Dairy Sci. 1991, 74, 1730–1742. [Google Scholar] [CrossRef]
- Swaisgood, H.E. Chemistry of the Caseins. Adv. Dairy Chem. 1992, 1, 63. [Google Scholar]
- Baranyi, M.; Boesze, Z.; Buchberger, J.; Krause, I. Genetic Polymorphism of Milk Proteins in Hungarian Cattle Breeds and PCR Amplification of Beta-Lactoglobulin EXON 5 to Identify Genetic Variant J by RFLP; International Dairy Federation: Brussels, Belgium, 1997. [Google Scholar]
- Lien, S.; Rogne, S. Bovine Casein Haplotypes: Number, Frequencies and Applicability as Genetic Markers. Anim. Genet. 1993, 24, 373–376. [Google Scholar] [CrossRef]
- Ikonene, T.; Ojala, M.; Ruottinen, O. Effects of Beta- and Kappa-Casein Genotypes on First Lactation Milk Production Traits in Finnish Ayrshire Cows; International Dairy Federation: Brussels, Belgium, 1997. [Google Scholar]
- Curik, I.; Havranek, J.; Samarzija, D. Milk Protein Polymorphism and Genetic Structure of Croatian Simmental Cattle; International Dairy Federation: Brussels, Belgium, 1997. [Google Scholar]
- Ehrmann, S.; Bartenschlager, H.; Geldermann, H. Quantification of Gene Effects on Single Milk Proteins in Selected Groups of Dairy Cows. J. Anim. Breed. Genet. 1997, 114, 121–132. [Google Scholar] [CrossRef]
- Winkelman, A.M.; Wickham, B.W. Associations between Milk Protein Genetic Variants and Production Traits in New Zealand Dairy Cattle; International Dairy Federation: Brussels, Belgium, 1997. [Google Scholar]
- Boettcher, P.J.; Caroli, A.; Stella, A.; Chessa, S.; Budelli, E.; Canavesi, F.; Ghiroldi, S.; Pagnacco, G. Effects of Casein Haplotypes on Milk Production Traits in Italian Holstein and Brown Swiss Cattle. J. Dairy Sci. 2004, 87, 4311–4317. [Google Scholar] [CrossRef] [Green Version]
- Kamiński, S.; Ruść, A.; Cieślińska, A. A Note on Frequency of A1 and A2 Variants of Bovine Beta-Casein Locus in Polish Holstein Bulls. J. Anim. Feed Sci. 2006, 15, 195–198. [Google Scholar] [CrossRef]
- Heck, J.M.L.; Schennink, A.; van Valenberg, H.J.F.; Bovenhuis, H.; Visker, M.H.P.W.; van Arendonk, J.A.M.; van Hooijdonk, A.C.M. Effects of Milk Protein Variants on the Protein Composition of Bovine Milk. J. Dairy Sci. 2009, 92, 1192–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visker, M.H.P.W.; Dibbits, B.W.; Kinders, S.M.; van Valenberg, H.J.F.; van Arendonk, J.A.M.; Bovenhuis, H. Association of Bovine β-Casein Protein Variant I with Milk Production and Milk Protein Composition. Anim. Genet. 2011, 42, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Molee, A.; Boonek, L.; Rungsakinnin, N. The Effect of Beta and Kappa Casein Genes on Milk Yield and Milk Composition in Different Percentages of Holstein in Crossbred Dairy Cattle. Anim. Sci. J. 2011, 82, 512–516. [Google Scholar] [CrossRef]
- Cieślińska, A.; Kostyra, E.; Kostyra, H.; Oleński, K.; Fiedorowicz, E.; Kamiński, S. Milk from Cows of Different β-Casein Genotypes as a Source of β-Casomorphin-7. Int. J. Food Sci. Nutr. 2012, 63, 426–430. [Google Scholar] [CrossRef]
- Dinc, H.; Ozkan, E.; Koban, E.; Togan, I. Beta-Casein A1/A2, Kappa-Casein and Beta-Lactoglobulin Polymorphisms in Turkish Cattle Breeds. Arch. Anim. Breed. 2013, 56, 650–657. [Google Scholar] [CrossRef] [Green Version]
- Mir, S.N.; Ullah, O.; Sheikh, R. Genetic Polymorphism of Milk Protein Variants and Their Association Studies with Milk Yield in Sahiwal Cattle. Afr. J. Biotechnol. 2014, 13, 555–565. [Google Scholar] [CrossRef]
- Gustavsson, F.; Buitenhuis, A.J.; Johansson, M.; Bertelsen, H.P.; Glantz, M.; Poulsen, N.A.; Lindmark Månsson, H.; Stålhammar, H.; Larsen, L.B.; Bendixen, C.; et al. Effects of Breed and Casein Genetic Variants on Protein Profile in Milk from Swedish Red, Danish Holstein, and Danish Jersey Cows. J. Dairy Sci. 2014, 97, 3866–3877. [Google Scholar] [CrossRef] [Green Version]
- Gholami, M.; Hafezian, S.H.; Rahimi, G.; Farhadi, A.; Rahimi, Z.; Kahrizi, D.; Kiani, S.; Karim, H.; Vaziri, S.; Muhammadi, S.; et al. Allele Specific-PCR and Melting Curve Analysis Showed Relatively High Frequency of β-Casein Gene A1 Allele in Iranian Holstein, Simmental and Native Cows. Cell. Mol. Biol. (Noisy-Le-Grand) 2016, 62, 138–143. [Google Scholar] [CrossRef]
- Dai, R.; Fang, Y.; Zhao, W.; Liu, S.; Ding, J.; Xu, K.; Yang, L.; He, C.; Ding, F.; Meng, H. Identification of Alleles and Genotypes of Beta-Casein with DNA Sequencing Analysis in Chinese Holstein Cow. J. Dairy Res. 2016, 83, 312–316. [Google Scholar] [CrossRef]
- Rangel, A.H.N.; Zaros, L.G.; Lima, T.C.; Borba, L.H.F.; Novaes, L.P.; Mota, L.F.M.; Silva, M.S. Polymorphism in the Beta Casein Gene and Analysis of Milk Characteristicsin Gir and Guzerá Dairy Cattle. Genet. Mol. Res. 2017, 16, gmr16029592. [Google Scholar] [CrossRef] [PubMed]
- Massella, E.; Piva, S.; Giacometti, F.; Liuzzo, G.; Zambrini, A.V.; Serraino, A. Evaluation of Bovine Beta Casein Polymorphism in Two Dairy Farms Located in Northern Italy. Ital. J. Food Saf. 2017, 6, 6904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uniacke-Lowe, T.; Huppertz, T.; Fox, P.F. Equine Milk Proteins: Chemistry, Structure and Nutritional Significance. Int. Dairy J. 2010, 20, 609–629. [Google Scholar] [CrossRef]
- Cheng, H.; Dong, H.; Wusigale; Liang, L. A Comparison of β-Casein Complexes and Micelles as Vehicles for Trans-/Cis-Resveratrol. Food Chem. 2020, 330, 127209. [Google Scholar] [CrossRef] [PubMed]
- Duerasch, A.; Herrmann, P.; Hogh, K.; Henle, T. Study on β-Casein Depleted Casein Micelles: Micellar Stability, Enzymatic Cross-Linking, and Suitability as Nanocarriers. J. Agric. Food Chem. 2020, 68, 13940–13949. [Google Scholar] [CrossRef]
- Li, M.; Fokkink, R.; Ni, Y.; Kleijn, J.M. Bovine Beta-Casein Micelles as Delivery Systems for Hydrophobic Flavonoids. Food Hydrocoll. 2019, 96, 653–662. [Google Scholar] [CrossRef]
- Raynes, J.K.; Day, L.; Augustin, M.A.; Carver, J.A. Structural Differences between Bovine A1 and A2 β-Casein Alter Micelle Self-Assembly and Influence Molecular Chaperone Activity. J. Dairy Sci. 2015, 98, 2172–2182. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yu, Z.; Zhao, X.; Han, R.; Huang, D.; Yang, Y.; Cheng, G. Comparative Proteomic Characterization of Bovine Milk Containing β-Casein Variants A1A1 and A2A2, and Their Heterozygote A1A2. J. Sci. Food Agric. 2021, 101, 718–725. [Google Scholar] [CrossRef]
- Petrat-Melin, B.; Andersen, P.; Rasmussen, J.T.; Poulsen, N.A.; Larsen, L.B.; Young, J.F. In Vitro Digestion of Purified β-Casein Variants A(1), A(2), B, and I: Effects on Antioxidant and Angiotensin-Converting Enzyme Inhibitory Capacity. J. Dairy Sci. 2015, 98, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Brantl, V.; Teschemacher, H.; Henschen, A.; Lottspeich, F. Novel Opioid Peptides Derived from Casein (Beta-Casomorphins). I. Isolation from Bovine Casein Peptone. Hoppe Seylers Z. Physiol. Chem. 1979, 360, 1211–1216. [Google Scholar] [CrossRef]
- Chang, K.J.; Su, Y.F.; Brent, D.A.; Chang, J.K. Isolation of a Specific Mu-Opiate Receptor Peptide, Morphiceptin, from an Enzymatic Digest of Milk Proteins. J. Biol. Chem. 1985, 260, 9706–9712. [Google Scholar] [CrossRef] [PubMed]
- Kostyra, E.; Sienkiewicz-Szłapka, E.; Jarmołowska, B.; Krawczuk, S.; Kostyra, H. Opioid Peptides Derived from Milk Proteins. Pol. J. Food Nutr. Sci. 2004, 13, 25–35. [Google Scholar]
- Meisel, H.; FitzGerald, R.J. Opioid Peptides Encrypted in Intact Milk Protein Sequences. Br. J. Nutr. 2000, 84 (Suppl. 1), S27–S31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylund, G.; Pettersson, A.; Bengtsson, C.; Khorram-Manesh, A.; Nordgren, S.; Delbro, D.S. Functional Expression of Mu-Opioid Receptors in the Human Colon Cancer Cell Line, HT-29, and Their Localization in Human Colon. Dig. Dis. Sci. 2008, 53, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Bidlack, J.M. Detection and Function of Opioid Receptors on Cells from the Immune System. Clin. Diagn. Lab. Immunol. 2000, 7, 719–723. [Google Scholar] [CrossRef] [Green Version]
- Mansour, A.; Fox, C.A.; Akil, H.; Watson, S.J. Opioid-Receptor MRNA Expression in the Rat CNS: Anatomical and Functional Implications. Trends Neurosci. 1995, 18, 22–29. [Google Scholar] [CrossRef]
- Teschemacher, H. Opioid Receptor Ligands Derived from Food Proteins. Curr. Pharm. Des. 2003, 9, 1331–1344. [Google Scholar] [CrossRef] [Green Version]
- Phelan, M.; Aherne, A.; FitzGerald, R.J.; O’Brien, N.M. Casein-Derived Bioactive Peptides: Biological Effects, Industrial Uses, Safety Aspects and Regulatory Status. Int. Dairy J. 2009, 19, 643–654. [Google Scholar] [CrossRef]
- Schmelzer, C.E.H.; Schöps, R.; Reynell, L.; Ulbrich-Hofmann, R.; Neubert, R.H.H.; Raith, K. Peptic Digestion of β-Casein: Time Course and Fate of Possible Bioactive Peptides. J. Chromatogr. A 2007, 1166, 108–115. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Bioactive Peptides: Production and Functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Johansson, M.; Åkerstedt, M.; Li, S.; Zamaratskaia, G.; Sternesjö Lundh, Å. Casein Breakdown in Bovine Milk by a Field Strain of Staphylococcus Aureus. J. Food Prot. 2013, 76, 1638–1642. [Google Scholar] [CrossRef] [PubMed]
- Noni, I.D. Release of β-Casomorphins 5 and 7 during Simulated Gastro-Intestinal Digestion of Bovine β-Casein Variants and Milk-Based Infant Formulas. Food Chem. 2008, 110, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, H.; Pihlanto-Leppäla, A.; Rantamäki, P.; Tupasela, T. Impact of Processing on Bioactive Proteins and Peptides. Trends Food Sci. Technol. 1998, 9, 307–319. [Google Scholar] [CrossRef]
- Hartwig, A. Einfluß Genetischer Varianten Bovinen β-Caseins [beta-Caseins] auf Entstehung und Aktivität Biologisch Aktiver Peptide. Ph.D. Dissertation, University of Giessen, Giessen, Germany, 1997. [Google Scholar]
- Meisel, H. Biochemical Properties of Regulatory Peptides Derived from Mil Proteins. Pept. Sci. 1997, 43, 119–128. [Google Scholar] [CrossRef]
- Jinsmaa, Y.; Yoshikawa, M. Enzymatic Release of Neocasomorphin and Beta-Casomorphin from Bovine Beta-Casein. Peptides 1999, 20, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, H.; Pihlanto, A. Food-Derived Bioactive Peptides—Opportunities for Designing Future Foods. Curr. Pharm. Des. 2003, 9, 1297–1308. [Google Scholar] [CrossRef] [Green Version]
- Ul Haq, M.R.; Kapila, R.; Kapila, S. Release of β-Casomorphin-7/5 during Simulated Gastrointestinal Digestion of Milk β-Casein Variants from Indian Crossbred Cattle (Karan Fries). Food Chem. 2015, 168, 70–79. [Google Scholar] [CrossRef]
- Elliott, R.B.; Harris, D.P.; Hill, J.P.; Bibby, N.J.; Wasmuth, H.E. Type I (Insulin-Dependent) Diabetes Mellitus and Cow Milk: Casein Variant Consumption. Diabetologia 1999, 42, 292–296. [Google Scholar] [CrossRef] [Green Version]
- McLachlan, C.N. Beta-Casein A1, Ischaemic Heart Disease Mortality, and Other Illnesses. Med. Hypotheses 2001, 56, 262–272. [Google Scholar] [CrossRef] [Green Version]
- Cieślińska, A.; Kamiński, S.; Kostyra, E.; Sienkiewicz-Szłapka, E. Beta-Casomorphin 7 in Raw and Hydrolyzed Milk Derived from Cows of Alternative β-Casein Genotypes. Milchwiss. Milk Sci. Int. 2007, 62, 125–127. [Google Scholar]
- Duarte-Vázquez, M.Á.; García-Ugalde, C.; Villegas-Gutiérrez, L.M.; García-Almendárez, B.E.; Rosado, J.L. Production of Cow’s Milk Free from Beta-Casein A1 and Its Application in the Manufacturing of Specialized Foods for Early Infant Nutrition. Foods 2017, 6, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambers, T.T.; Broeren, S.; Heck, J.; Bragt, M.; Huppertz, T. Processing Affects Beta-Casomorphin Peptide Formation during Simulated Gastrointestinal Digestion in Both A1 and A2 Milk. Int. Dairy J. 2021, 121, 105099. [Google Scholar] [CrossRef]
- Svedberg, J.; de Haas, J.; Leimenstoll, G.; Paul, F.; Teschemacher, H. Demonstration of Beta-Casomorphin Immunoreactive Materials in in Vitro Digests of Bovine Milk and in Small Intestine Contents after Bovine Milk Ingestion in Adult Humans. Peptides 1985, 6, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Boutrou, R.; Gaudichon, C.; Dupont, D.; Jardin, J.; Airinei, G.; Marsset-Baglieri, A.; Benamouzig, R.; Tomé, D.; Leonil, J. Sequential Release of Milk Protein-Derived Bioactive Peptides in the Jejunum in Healthy Humans. Am. J. Clin. Nutr. 2013, 97, 1314–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FitzGerald, R.J.; Murray, B.A.; Walsh, D.J. Hypotensive Peptides from Milk Proteins. J. Nutr. 2004, 134, 980S–988S. [Google Scholar] [CrossRef] [PubMed]
- Ramabadran, K. Pharmacology of β-Casomorphins, Opioid Peptides Derived from Milk Protein. Asia Pac. J. Pharm. 1989, 4, 45–58. [Google Scholar]
- Hafeez, Z.; Cakir-Kiefer, C.; Girardet, J.-M.; Jardin, J.; Perrin, C.; Dary, A.; Miclo, L. Hydrolysis of Milk-Derived Bioactive Peptides by Cell-Associated Extracellular Peptidases of Streptococcus Thermophilus. Appl. Microbiol. Biotechnol. 2013, 97, 9787–9799. [Google Scholar] [CrossRef] [PubMed]
- Asledottir, T.; Picariello, G.; Mamone, G.; Ferranti, P.; Røseth, A.; Devold, T.G.; Vegarud, G.E. Degradation of β-Casomorphin-7 through in Vitro Gastrointestinal and Jejunal Brush Border Membrane Digestion. J. Dairy Sci. 2019, 102, 8622–8629. [Google Scholar] [CrossRef]
- Engel, M.; Hoffmann, T.; Wagner, L.; Wermann, M.; Heiser, U.; Kiefersauer, R.; Huber, R.; Bode, W.; Demuth, H.-U.; Brandstetter, H. The Crystal Structure of Dipeptidyl Peptidase IV (CD26) Reveals Its Functional Regulation and Enzymatic Mechanism. Proc. Natl. Acad. Sci. USA 2003, 100, 5063–5068. [Google Scholar] [CrossRef] [Green Version]
- Lambeir, A.-M.; Durinx, C.; Scharpé, S.; De Meester, I. Dipeptidyl-Peptidase IV from Bench to Bedside: An Update on Structural Properties, Functions, and Clinical Aspects of the Enzyme DPP IV. Crit. Rev. Clin. Lab. Sci. 2003, 40, 209–294. [Google Scholar] [CrossRef]
- Kreil, G.; Umbach, M.; Brantl, V.; Teschemacher, H. Studies on the Enzymatic Degradation of Beta-Casomorphins. Life Sci. 1983, 33 (Suppl. 1), 137–140. [Google Scholar] [CrossRef] [PubMed]
- Osborne, S.; Chen, W.; Addepalli, R.; Colgrave, M.; Singh, T.; Tran, C.; Day, L. In Vitro Transport and Satiety of a Beta-Lactoglobulin Dipeptide and Beta-Casomorphin-7 and Its Metabolites. Food Funct. 2014, 5, 2706–2718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sienkiewicz-Szłapka, E.; Jarmołowska, B.; Krawczuk, S.; Kostyra, E.; Kostyra, H.; Bielikowicz, K. Transport of Bovine Milk-Derived Opioid Peptides across a Caco-2 Monolayer. Int. Dairy J. 2009, 19, 252–257. [Google Scholar] [CrossRef]
- Jarmołowska, B.; Teodorowicz, M.; Fiedorowicz, E.; Sienkiewicz-Szłapka, E.; Matysiewicz, M.; Kostyra, E. Glucose and Calcium Ions May Modulate the Efficiency of Bovine β-Casomorphin-7 Permeability through a Monolayer of Caco-2 Cells. Peptides 2013, 49, 59–67. [Google Scholar] [CrossRef] [PubMed]
- De Noni, I.; Cattaneo, S. Occurrence of β-Casomorphins 5 and 7 in Commercial Dairy Products and in Their Digests Following in Vitro Simulated Gastro-Intestinal Digestion. Food Chem. 2010, 119, 560–566. [Google Scholar] [CrossRef]
- Jarmołowska, B.; Krawczuk, S. The Influence of Storage on Contents of Selected Antagonist and Agonist Opioid Peptides in Fermented Milk Drinks. Milchwissenschaft 2012, 67, 130–133. [Google Scholar]
- Nguyen, D.D.; Solah, V.A.; Johnson, S.K.; Charrois, J.W.A.; Busetti, F. Isotope Dilution Liquid Chromatography–Tandem Mass Spectrometry for Simultaneous Identification and Quantification of Beta-Casomorphin 5 and Beta-Casomorphin 7 in Yoghurt. Food Chem. 2014, 146, 345–352. [Google Scholar] [CrossRef]
- Muehlenkamp, M.R.; Warthesen, J.J. Beta-Casomorphins: Analysis in Cheese and Susceptibility to Proteolytic Enzymes from Lactococcus Lactis Ssp. Cremoris. J. Dairy Sci. 1996, 79, 20–26. [Google Scholar] [CrossRef]
- Saito, T. Antihypertensive Peptides Derived from Bovine Casein and Whey Proteins. Adv. Exp. Med. Biol. 2008, 606, 295–317. [Google Scholar] [CrossRef]
- Toelstede, S.; Hofmann, T. Quantitative Studies and Taste Re-Engineering Experiments toward the Decoding of the Nonvolatile Sensometabolome of Gouda Cheese. J. Agric. Food. Chem. 2008, 56, 5299–5307. [Google Scholar] [CrossRef]
- Jarmołowska, B.; Kostyra, E.; Krawczuk, S.; Kostyra, H. β-Casomorphin-7 Isolated from Brie Cheese. J. Sci. Food Agric. 1999, 79, 1788–1792. [Google Scholar] [CrossRef]
- Contents of Agonistic and Antagonistic Opioid Peptides in Different Cheese Varieties. Available online: https://nauka-polska.pl/#/profile/publication?id=2118297&_k=z8mnca (accessed on 2 November 2022).
- Jarmołowska, B.; Bielikowicz, K.; Iwan, M.; Sidor, K.; Kostyra, E.; Kaczmarski, M. Serum Activity of Dipeptidyl Peptidase IV (DPPIV; EC 3.4.14.5) in Breast-Fed Infants with Symptoms of Allergy. Peptides 2007, 28, 678–682. [Google Scholar] [CrossRef]
- Sturner, R.; Chang, K. Opioid Peptide Content in Infant Formulas. Pediatr. Res 1988, 23, 4–10. [Google Scholar]
- Küllenberg de Gaudry, D.; Lohner, S.; Schmucker, C.; Kapp, P.; Motschall, E.; Hörrlein, S.; Röger, C.; Meerpohl, J.J. Milk A1 β-Casein and Health-Related Outcomes in Humans: A Systematic Review. Nutr. Rev. 2019, 77, 278–306. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, R.V.; Kumar, A.; Yadav, J.S. Analysis of Beta-Casein Gene (CSN2) Polymorphism in Tharparkar and Frieswal Cattle. Indian J. Anim. Res. 2019, 54, 1–5. [Google Scholar] [CrossRef]
- Thorsdottir, I.; Birgisdottir, B.E.; Johannsdottir, I.M.; Harris, D.P.; Hill, J.; Steingrimsdottir, L.; Thorsson, A.V. Different Beta-Casein Fractions in Icelandic versus Scandinavian Cow’s Milk May Influence Diabetogenicity of Cow’s Milk in Infancy and Explain Low Incidence of Insulin-Dependent Diabetes Mellitus in Iceland. Pediatrics 2000, 106, 719–724. [Google Scholar] [CrossRef]
- Laugesen, M.; Elliott, R. Ischaemic Heart Disease, Type 1 Diabetes, and Cow Milk A1 Beta-Casein. N. Z. Med. J. 2003, 116, U295. [Google Scholar]
- Sun, Z.; Zhang, Z.; Wang, X.; Cade, R.; Elmir, Z.; Fregly, M. Relation of β-Casomorphin to Apnea in Sudden Infant Death Syndrome. Peptides 2003, 24, 937–943. [Google Scholar] [CrossRef]
- Tailford, K.A.; Berry, C.L.; Thomas, A.C.; Campbell, J.H. A Casein Variant in Cow’s Milk Is Atherogenic. Atherosclerosis 2003, 170, 13–19. [Google Scholar] [CrossRef]
- Kost, N.V.; Sokolov, O.Y.; Kurasova, O.B.; Dmitriev, A.D.; Tarakanova, J.N.; Gabaeva, M.V.; Zolotarev, Y.A.; Dadayan, A.K.; Grachev, S.A.; Korneeva, E.V.; et al. β-Casomorphins-7 in Infants on Different Type of Feeding and Different Levels of Psychomotor Development. Peptides 2009, 30, 1854–1860. [Google Scholar] [CrossRef]
- Reichelt, K.L.; Tveiten Bioengineer, D.; Knivsberg, A.-M.; Brønstad, G. Peptides’ Role in Autism with Emphasis on Exorphins. Microb. Ecol. Health Dis. 2012, 23, 18958. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarski, M.; Wasilewska, J.; Lasota, M. Hypersensitivity to Hydrolyzed Cow’s Milk Protein Formula in Infants and Young Children with Atopic Eczema/Dermatitis Syndrome with Cow’s Milk Protein Allergy. Rocz. Akad. Med. Bialymst. 2005, 50, 274–278. [Google Scholar] [PubMed]
- Arvola, T.; Moilanen, E.; Vuento, R.; Isolauri, E. Weaning to Hypoallergenic Formula Improves Gut Barrier Function in Breast-Fed Infants with Atopic Eczema. J. Pediatr. Gastroenterol. Nutr. 2004, 38, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Chatchatee, P.; Järvinen, K.-M.; Bardina, L.; Beyer, K.; Sampson, H.A. Identification of IgE- and IgG-Binding Epitopes on As1-Casein: Differences in Patients with Persistent and Transient Cow’s Milk Allergy. J. Allergy Clin. Immunol. 2001, 107, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M.; Stepaniak, L.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Latent Bioactive Peptides in Milk Proteins: Proteolytic Activation and Significance in Dairy Processing. Crit. Rev. Food Sci. Nutr. 2002, 42, 223–239. [Google Scholar] [CrossRef]
- Zoghbi, S.; Trompette, A.; Claustre, J.; Homsi, M.E.; Garzón, J.; Jourdan, G.; Scoazec, J.-Y.; Plaisancié, P. β-Casomorphin-7 Regulates the Secretion and Expression of Gastrointestinal Mucins through a μ-Opioid Pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G1105–G1113. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Sun, J.; Jiang, Z.Q.; Yang, Y.X. Effects of Cow’s Milk Beta-Casein Variants on Symptoms of Milk Intolerance in Chinese Adults: A Multicentre, Randomised Controlled Study. Nutr. J. 2017, 16, 72. [Google Scholar] [CrossRef] [Green Version]
- Milan, A.M.; Shrestha, A.; Karlström, H.J.; Martinsson, J.A.; Nilsson, N.J.; Perry, J.K.; Day, L.; Barnett, M.P.G.; Cameron-Smith, D. Comparison of the Impact of Bovine Milk β-Casein Variants on Digestive Comfort in Females Self-Reporting Dairy Intolerance: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2020, 111, 149–160. [Google Scholar] [CrossRef]
- Ramakrishnan, M.; Eaton, T.K.; Sermet, O.M.; Savaiano, D.A. Milk Containing A2 β-Casein ONLY, as a Single Meal, Causes Fewer Symptoms of Lactose Intolerance than Milk Containing A1 and A2 β-Caseins in Subjects with Lactose Maldigestion and Intolerance: A Randomized, Double-Blind, Crossover Trial. Nutrients 2020, 12, 3855. [Google Scholar] [CrossRef]
- Brooke-Taylor, S.; Dwyer, K.; Woodford, K.; Kost, N. Systematic Review of the Gastrointestinal Effects of A1 Compared with A2 β-Casein. Adv. Nutr. 2017, 8, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Beneficial Effects of Milk Having A2 β-Casein Protein: Myth or Reality?|The Journal of Nutrition|Oxford Academic. Available online: https://academic.oup.com/jn/article/151/5/1061/6165053 (accessed on 19 October 2022).
- Migliore-Samour, D.; Jollès, P. Casein, a Prohormone with an Immunomodulating Role for the Newborn? Experientia 1988, 44, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Kurek, M.; Czerwionka-Szaflarska, M.; Doroszewska, G. Pseudoallergic Skin Reactions to Opiate Sequences of Bovine Casein in Healthy Children. Rocz. Akad. Med. Bialymst. 1995, 40, 480–485. [Google Scholar] [PubMed]
- Kurek, M.; Przybilla, B.; Hermann, K.; Ring, J. A Naturally Occurring Opioid Peptide from Cow’s Milk, Beta-Casomorphine-7, Is a Direct Histamine Releaser in Man. Int. Arch. Allergy Immunol. 1992, 97, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Fiedorowicz, E.; Markiewicz, L.H.; Sidor, K.; Świątecka, D.; Cieślińska, A.; Matysiewicz, M.; Piskorz-Ogórek, K.; Sienkiewicz-Szłapka, E.; Teodorowicz, M.; Świątecki, A.; et al. The Influence of Breast Milk and Infant Formulae Hydrolysates on Bacterial Adhesion and Caco-2 Cells Functioning. Food Res. Int. 2016, 89, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Summer, A.; Di Frangia, F.; Ajmone Marsan, P.; De Noni, I.; Malacarne, M. Occurrence, Biological Properties and Potential Effects on Human Health of β-Casomorphin 7: Current Knowledge and Concerns. Crit. Rev. Food Sci. Nutr. 2020, 60, 3705–3723. [Google Scholar] [CrossRef]
- Pal, S.; Woodford, K.; Kukuljan, S.; Ho, S. Milk Intolerance, Beta-Casein and Lactose. Nutrients 2015, 7, 7285–7297. [Google Scholar] [CrossRef]
- Haq, M.R.U.; Kapila, R.; Saliganti, V. Consumption of β-Casomorphins-7/5 Induce Inflammatory Immune Response in Mice Gut through Th2 Pathway. J. Funct. Foods 2014, 8, 150–160. [Google Scholar] [CrossRef]
- Barnett, M.P.G.; McNabb, W.C.; Roy, N.C.; Woodford, K.B.; Clarke, A.J. Dietary A1 β-Casein Affects Gastrointestinal Transit Time, Dipeptidyl Peptidase-4 Activity, and Inflammatory Status Relative to A2 β-Casein in Wistar Rats. Int. J. Food Sci. Nutr. 2014, 65, 720–727. [Google Scholar] [CrossRef]
- Ho, S.; Woodford, K.; Kukuljan, S.; Pal, S. Comparative Effects of A1 versus A2 Beta-Casein on Gastrointestinal Measures: A Blinded Randomised Cross-over Pilot Study. Eur. J. Clin. Nutr. 2014, 68, 994–1000. [Google Scholar] [CrossRef]
- Jianqin, S.; Leiming, X.; Lu, X.; Yelland, G.W.; Ni, J.; Clarke, A.J. Effects of Milk Containing Only A2 Beta Casein versus Milk Containing Both A1 and A2 Beta Casein Proteins on Gastrointestinal Physiology, Symptoms of Discomfort, and Cognitive Behavior of People with Self-Reported Intolerance to Traditional Cows’ Milk. Nutr. J. 2016, 15, 35. [Google Scholar] [CrossRef] [Green Version]
- Wageningen University & Research; Cieslinska, A.; Kostyra, E.; Savelkoul, H.F.J. Treating Autism Spectrum Disorder with Gluten-Free and Casein-Free Diet: The Underlying Microbiota-Gut-Brain Axis Mechanisms. CIIT 2017, 3, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deth, R.; Clarke, A.; Ni, J.; Trivedi, M. Clinical Evaluation of Glutathione Concentrations after Consumption of Milk Containing Different Subtypes of β-Casein: Results from a Randomized, Cross-over Clinical Trial. Nutr. J. 2016, 15, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, X.; Li, Z.; Ni, J.; Yelland, G. Effects of Conventional Milk Versus Milk Containing Only A2 β-Casein on Digestion in Chinese Children: A Randomized Study. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Stępnik, M.; Kurek, M. The Influence of Bovine Casein-Derived Exorphins on Mast Cells in Rodents. Rev. Française D’allergologie Et D’immunologie Clin. 2002, 42, 447–453. [Google Scholar] [CrossRef]
- Fiedorowicz, E.; Kaczmarski, M.; Cieślińska, A.; Sienkiewicz-Szłapka, E.; Jarmołowska, B.; Chwała, B.; Kostyra, E. β-casomorphin-7 alters μ-opioid receptor and dipeptidyl peptidase IV genes expression in children with atopic dermatitis. Peptides 2014, 62, 144–149. [Google Scholar] [CrossRef]
- Fiedorowicz, E.; Jarmołowska, B.; Iwan, M.; Kostyra, E.; Obuchowicz, R.; Obuchowicz, M. The Influence of μ-Opioid Receptor Agonist and Antagonist Peptides on Peripheral Blood Mononuclear Cells (PBMCs). Peptides 2011, 32, 707–712. [Google Scholar] [CrossRef]
- Gillespie, K.M. Type 1 Diabetes: Pathogenesis and Prevention. CMAJ 2006, 175, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Woodford, K.B. The A2 Milk Debate—Searching for the Evidence. Prim. Ind. Manag. 2004, 7, 29–33. [Google Scholar]
- Cavallo, M.G.; Monetini, L.; Walker, B.; Thorpe, R.; Pozzilli, P. Diabetes and Cows’ Milk. Lancet 1996, 348, 1655. [Google Scholar] [CrossRef]
- Birgisdottir, B.E.; Hill, J.P.; Thorsson, A.V.; Thorsdottir, I. Lower Consumption of Cow Milk Protein A1 Beta-Casein at 2 Years of Age, Rather than Consumption among 11- to 14-Year-Old Adolescents, May Explain the Lower Incidence of Type 1 Diabetes in Iceland than in Scandinavia. Ann. Nutr. Metab. 2006, 50, 177–183. [Google Scholar] [CrossRef]
- Pozzilli, P. Beta-Casein in Cow’s Milk: A Major Antigenic Determinant for Type 1 Diabetes? J. Endocrinol. Investig. 1999, 22, 562–567. [Google Scholar] [CrossRef]
- Monetini, L.; Barone, F.; Stefanini, L.; Petrone, A.; Walk, T.; Jung, G.; Thorpe, R.; Pozzilli, P.; Cavallo, M.G. Establishment of T Cell Lines to Bovine Beta-Casein and Beta-Casein-Derived Epitopes in Patients with Type 1 Diabetes. J. Endocrinol. 2003, 176, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chia, J.S.J.; McRae, J.L.; Kukuljan, S.; Woodford, K.; Elliott, R.B.; Swinburn, B.; Dwyer, K.M. A1 Beta-Casein Milk Protein and Other Environmental Pre-Disposing Factors for Type 1 Diabetes. Nutr. Diabetes 2017, 7, e274. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, M.S.; Shah, J.S.; Al-Mughairy, S.; Hodgson, N.W.; Simms, B.; Trooskens, G.A.; Van Criekinge, W.; Deth, R.C. Food-Derived Opioid Peptides Inhibit Cysteine Uptake with Redox and Epigenetic Consequences. J. Nutr. Biochem. 2014, 25, 1011–1018. [Google Scholar] [CrossRef] [Green Version]
- Bruni, V.; Dei, M. Eating Disorders in Adolescence. In Good Practice in Pediatric and Adolescent Gynecology; Fulghesu, A.M., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 131–141. ISBN 978-3-319-57162-1. [Google Scholar]
- Yin, H.; Miao, J.; Zhang, Y. Protective Effect of β-Casomorphin-7 on Type 1 Diabetes Rats Induced with Streptozotocin. Peptides 2010, 31, 1725–1729. [Google Scholar] [CrossRef]
- Yin, H.; Miao, J.; Ma, C.; Sun, G.; Zhang, Y. β-Casomorphin-7 Cause Decreasing in Oxidative Stress and Inhibiting NF-ΚB-INOS-NO Signal Pathway in Pancreas of Diabetes Rats. J. Food Sci. 2012, 77, C278–C282. [Google Scholar] [CrossRef]
- Zhang, W.; Miao, J.; Wang, S.; Zhang, Y. The Protective Effects of Beta-Casomorphin-7 against Glucose -Induced Renal Oxidative Stress In Vivo and Vitro. PLoS ONE 2013, 8, e63472. [Google Scholar] [CrossRef]
- Zhang, W.; Miao, J.; Ma, C.; Han, D.; Zhang, Y. β-Casomorphin-7 Attenuates the Development of Nephropathy in Type I Diabetes via Inhibition of Epithelial-Mesenchymal Transition of Renal Tubular Epithelial Cells. Peptides 2012, 36, 186–191. [Google Scholar] [CrossRef]
- Clemens, R.A. Milk A1 and A2 Peptides and Diabetes. Nestle Nutr. Workshop Ser. Pediatr. Program 2011, 67, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Torreilles, J.; Guérin, M.C. Casein-derived peptides can promote human LDL oxidation by a peroxidase-dependent and metal-independent process. Comptes Rendus Seances Soc. Biol. Fil. 1995, 189, 933–942. [Google Scholar]
- Steinerová, A.; Racek, J.; Stožický, F.; Zima, T.; Fialová, L.; Lapin, A. Antibodies Against Oxidized LDL—Theory and Clinical Use. Physiol. Res. 2001, 50, 11. [Google Scholar]
- Steinerová, A.; Racek, J.; Stožický, F.; Tatzber, F.; Lapin, A. Autoantibodies against Oxidized LDL in the First Phase of Life. Requires Authentication 1999, 37, 913–917. [Google Scholar] [CrossRef]
- Heinecke, J. Mass Spectrometric Quantification of Amino Acid Oxidation Products in Proteins: Insights into Pathways That Promote LDL Oxidation in the Human Artery Wally. FASEB J. 1999, 13, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, S.; Kostyra, E.; Cieślińska, A.; Fiedorowicz, E. Consumption of Bovine SS-Casein Variants (A1 or A2) Does Not Affect Basic Hematological and Biochemical Indices. Milchwissenschaft 2012, 67, 238–241. [Google Scholar]
- Venn, B.J.; Skeaff, C.M.; Brown, R.; Mann, J.I.; Green, T.J. A Comparison of the Effects of A1 and A2 Beta-Casein Protein Variants on Blood Cholesterol Concentrations in New Zealand Adults. Atherosclerosis 2006, 188, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Han, D.-N.; Zhang, D.-H.; Wang, L.-P.; Zhang, Y.-S. Protective Effect of β-Casomorphin-7 on Cardiomyopathy of Streptozotocin-Induced Diabetic Rats via Inhibition of Hyperglycemia and Oxidative Stress. Peptides 2013, 44, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Kastin, A.J.; Banks, W.A.; Zadina, J.E. A Decade of Changing Perceptions about Neuropeptides. Ann. N. Y. Acad. Sci. 1990, 579, 1–7. [Google Scholar] [CrossRef]
- Banks, W.A.; Kastin, A.J. Saturable Transport of Peptides across the Blood-Brain Barrier. Life Sci. 1987, 41, 1319–1338. [Google Scholar] [CrossRef]
- Davis, T.P.; Abbruscato, T.J.; Egleton, R.D. Peptides at the Blood Brain Barrier: Knowing Me Knowing You. Peptides 2015, 72, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Rueda-Ruzafa, L.; Cruz, F.; Cardona, D.; Hone, A.J.; Molina-Torres, G.; Sánchez-Labraca, N.; Roman, P. Opioid System Influences Gut-Brain Axis: Dysbiosis and Related Alterations. Pharmacol. Res. 2020, 159, 104928. [Google Scholar] [CrossRef]
- Sun, Z.; Cade, R.J.; Fregly, M.J.; Privette, R.M. Beta-Casomorphine Induces Fos-like Immunoreactivity in Discrete Brain Regions Relevant to Schizophrenia and Autism. Autism 1999, 3, 67–83. [Google Scholar] [CrossRef]
- Reichelt, K.L.; Knivsberg, A.M. Can the Pathophysiology of Autism Be Explained by the Nature of the Discovered Urine Peptides? Nutr. Neurosci. 2003, 6, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, O.Y.; Pryanikova, N.A.; Kost, N.V.; Zolotarev, Y.A.; Ryukert, E.N.; Zozulya, A.A. Reactions between Beta-Casomorphins-7 and 5-HT2-Serotonin Receptors. Bull. Exp. Biol. Med. 2005, 140, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, O.; Kost, N.; Andreeva, O.; Korneeva, E.; Meshavkin, V.; Tarakanova, Y.; Dadayan, A.; Zolotarev, Y.; Grachev, S.; Mikheeva, I.; et al. Autistic Children Display Elevated Urine Levels of Bovine Casomorphin-7 Immunoreactivity. Peptides 2014, 56, 68–71. [Google Scholar] [CrossRef]
- Cieślińska, A.; Sienkiewicz-Szłapka, E.; Wasilewska, J.; Fiedorowicz, E.; Chwała, B.; Moszyńska-Dumara, M.; Cieśliński, T.; Bukało, M.; Kostyra, E. Influence of Candidate Polymorphisms on the Dipeptidyl Peptidase IV and μ-Opioid Receptor Genes Expression in Aspect of the β-Casomorphin-7 Modulation Functions in Autism. Peptides 2015, 65, 6–11. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Koseki, M.; Wakamatsu, M.; Matsumura, E. Effects of Systemic Administration of β-Casomorphin-5 on Learning and Memory in Mice. Eur. J. Pharmacol. 2006, 530, 81–87. [Google Scholar] [CrossRef]
- Bell, S.J.; Grochoski, G.T.; Clarke, A.J. Health Implications of Milk Containing β-Casein with the A2 Genetic Variant. Crit. Rev. Food Sci. Nutr. 2006, 46, 93–100. [Google Scholar] [CrossRef]
- Pasi, A.; Mahler, H.; Lansel, N.; Bernasconi, C.; Messiha, F.S. Beta-Casomorphin-Immunoreactivity in the Brain Stem of the Human Infant. Res. Commun. Chem. Pathol. Pharm. 1993, 80, 305–322. [Google Scholar]
- Wasilewska, J.; Kaczmarski, M.; Kostyra, E.; Iwan, M. Cow’s-Milk-Induced Infant Apnoea with Increased Serum Content of Bovine β-Casomorphin-5. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 772–775. [Google Scholar] [CrossRef]
- Wasilewska, J.; Sienkiewicz-Szłapka, E.; Kuźbida, E.; Jarmołowska, B.; Kaczmarski, M.; Kostyra, E. The Exogenous Opioid Peptides and DPPIV Serum Activity in Infants with Apnoea Expressed as Apparent Life Threatening Events (ALTE). Neuropeptides 2011, 45, 189–195. [Google Scholar] [CrossRef]
- Olenski, K.; Kamiński, S.; Szyda, J.; Cieslinska, A. Polymorphism of the Beta-Casein Gene and Its Associations with Breeding Value for Production Traits of Holstein–Friesian Bulls. Livest. Sci. 2010, 131, 137–140. [Google Scholar] [CrossRef]
- Nilsen, H.; Olsen, H.G.; Hayes, B.; Sehested, E.; Svendsen, M.; Nome, T.; Meuwissen, T.; Lien, S. Casein Haplotypes and Their Association with Milk Production Traits in Norwegian Red Cattle. Genet. Sel. Evol. 2009, 41, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, C.A.; Hickey, S.M.; Cullen, N.G.; Prosser, C.G.; Anderson, R.M.; Tate, M.L. Associations between Β-casein Genotype and Milk Yield and Composition in Grazing Dairy Cows. N. Z. J. Agric. Res. 2005, 48, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Kearney, J.F.; Amer, P.R.; Villanueva, B. Cumulative Discounted Expressions of Sire Genotypes for the Complex Vertebral Malformation and β-Casein Loci in Commercial Dairy Herds. J. Dairy Sci. 2005, 88, 4426–4433. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cieślińska, A.; Fiedorowicz, E.; Rozmus, D.; Sienkiewicz-Szłapka, E.; Jarmołowska, B.; Kamiński, S. Does a Little Difference Make a Big Difference? Bovine β-Casein A1 and A2 Variants and Human Health—An Update. Int. J. Mol. Sci. 2022, 23, 15637. https://doi.org/10.3390/ijms232415637
Cieślińska A, Fiedorowicz E, Rozmus D, Sienkiewicz-Szłapka E, Jarmołowska B, Kamiński S. Does a Little Difference Make a Big Difference? Bovine β-Casein A1 and A2 Variants and Human Health—An Update. International Journal of Molecular Sciences. 2022; 23(24):15637. https://doi.org/10.3390/ijms232415637
Chicago/Turabian StyleCieślińska, Anna, Ewa Fiedorowicz, Dominika Rozmus, Edyta Sienkiewicz-Szłapka, Beata Jarmołowska, and Stanisław Kamiński. 2022. "Does a Little Difference Make a Big Difference? Bovine β-Casein A1 and A2 Variants and Human Health—An Update" International Journal of Molecular Sciences 23, no. 24: 15637. https://doi.org/10.3390/ijms232415637
APA StyleCieślińska, A., Fiedorowicz, E., Rozmus, D., Sienkiewicz-Szłapka, E., Jarmołowska, B., & Kamiński, S. (2022). Does a Little Difference Make a Big Difference? Bovine β-Casein A1 and A2 Variants and Human Health—An Update. International Journal of Molecular Sciences, 23(24), 15637. https://doi.org/10.3390/ijms232415637