

Identification of Darunavir Derivatives for Inhibition of SARS-CoV-2 3CLpro
 ,
,
Abstract
:1. Introduction
2. Results
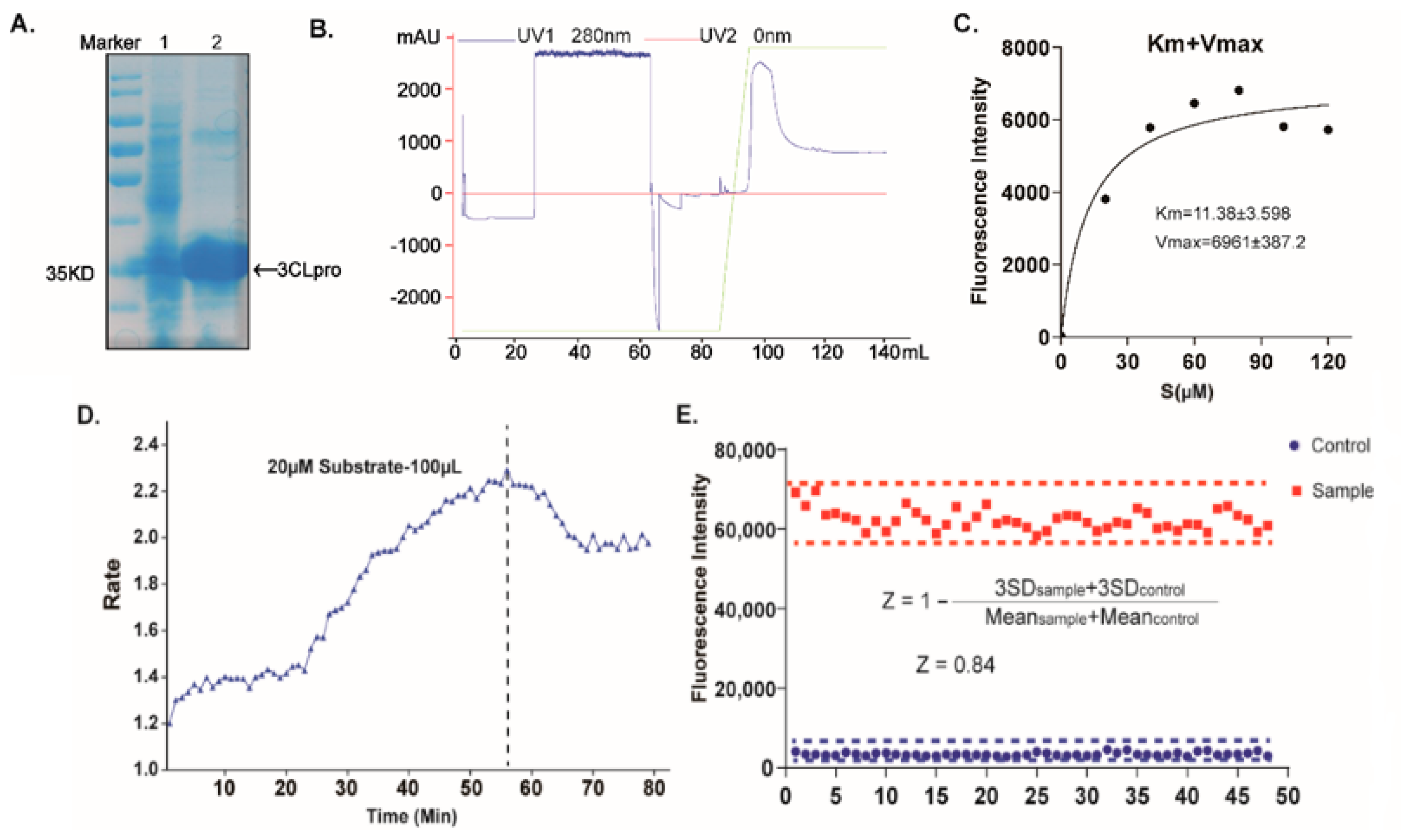
2.1. Establishing a High-Throughput Screening System with FRET Assay Targeting SARS-CoV-2 3CLpro
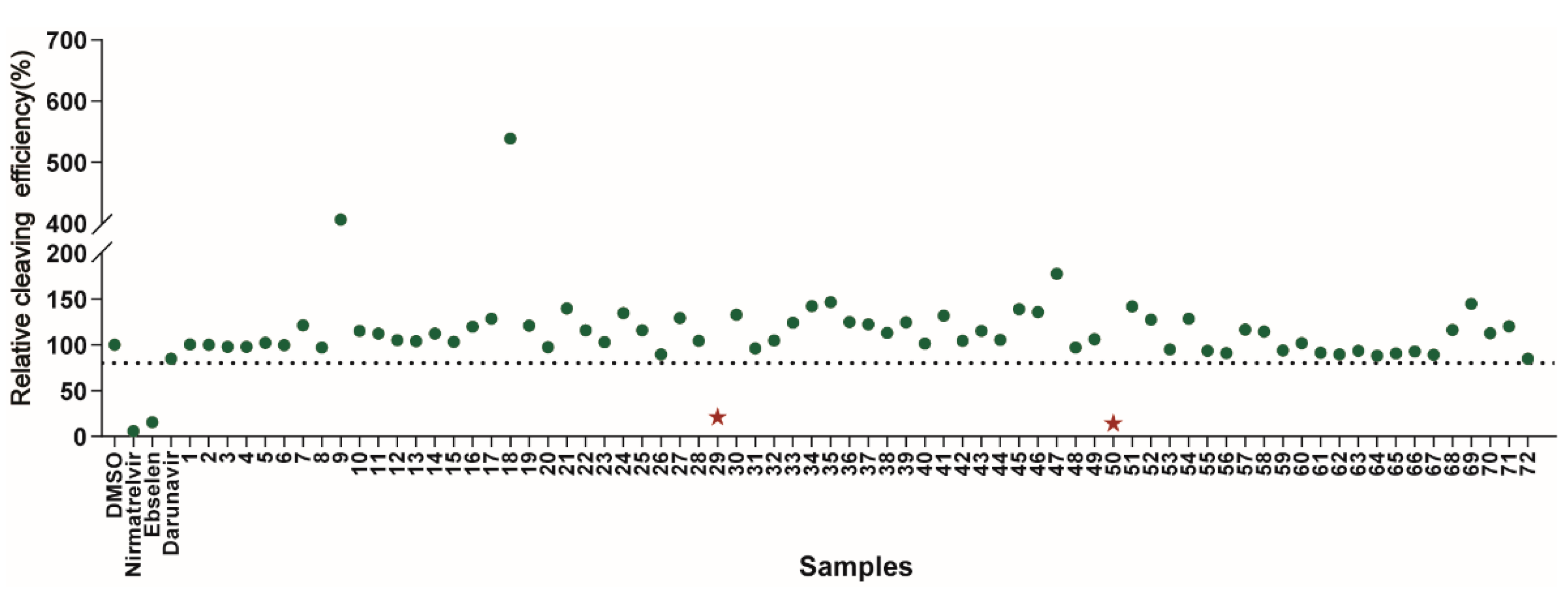
2.2. Screening of DRV Derivatives against SARS-CoV-2 3CLpro
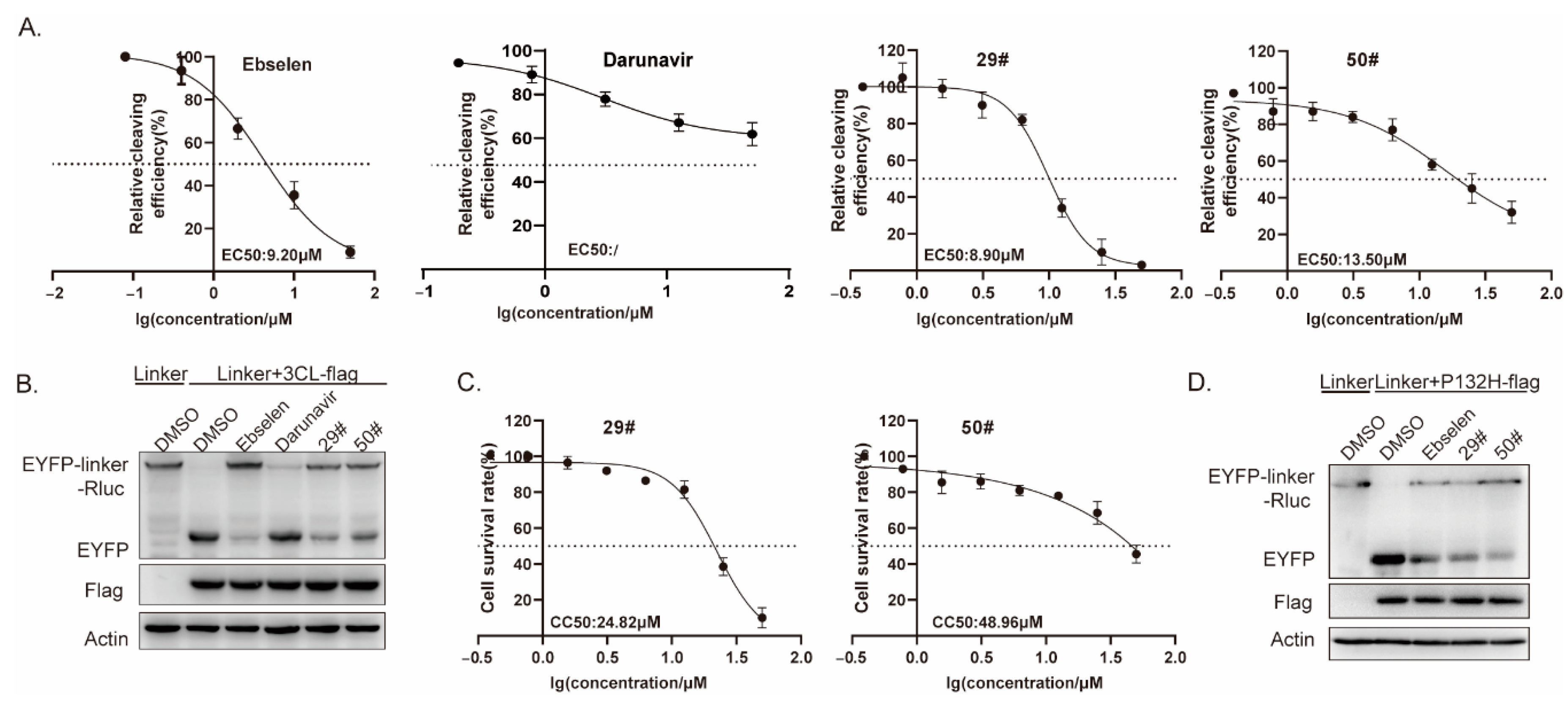
2.3. The Inhibitory Activity of Hits against SARS-CoV-2 3CLpro In Vitro
2.4. Binding of Hits to SARS-CoV-2 3CLpro
2.5. Modes of Interaction for SARS-CoV-2 3CLpro with Hits
2.6. The Inhibitory Activity of Hits against SARS-CoV-2 3CLpro at Cellular Level
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. Plasmid, Antibodies, and Reagents
4.3. In Vitro Enzymatic Inhibition Assay
4.4. BRET Assay
4.5. Western Blot
4.6. Cell Viability Assay
4.7. Bio-Layer Interferometry (BLI) Binding Assay
4.8. Docking (or Modeling)
4.9. Quantification and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. Addendum: A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 588, E6. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nature reviews. Microbiology 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kankanamalage, A.C.G.; Kim, Y.; Damalanka, V.C.; Rathnayake, A.D.; Fehr, A.R.; Mehzabeen, N.; Battaile, K.P.; Lovell, S.; Lushington, G.H.; Perlman, S.; et al. Structure-guided design of potent and permeable inhibitors of MERS coronavirus 3CL protease that utilize a piperidine moiety as a novel design element. Eur. J. Med. Chem. 2018, 150, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.; Su, H.; Zhao, W.; Xie, H.; Shao, Q.; Xu, Y. What coronavirus 3C-like protease tells us: From structure, substrate selectivity, to inhibitor design. Med. Res. Rev. 2021, 41, 1965–1998. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of M(pro) from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Zhang, B.; Jiang, X.-M.; Su, H.; Li, J.; Zhao, Y.; Xie, X.; Jin, Z.; Peng, J.; Liu, F.; et al. Structure-based design of antiviral drug candidates targeting the SARS-CoV-2 main protease. Science 2020, 368, 1331–1335. [Google Scholar] [CrossRef] [Green Version]
- Sk, F.; Roy, R.; Jonniya, N.A.; Poddar, S.; Kar, P. Elucidating biophysical basis of binding of inhibitors to SARS-CoV-2 main protease by using molecular dynamics simulations and free energy calculations. J. Biomol. Struct. Dyn. 2021, 39, 3649–3661. [Google Scholar] [CrossRef]
- Lobo-Galo, N.; Terrazas-López, M.; Martínez-Martínez, A.; Díaz-Sánchez, Á.G. FDA-approved thiol-reacting drugs that potentially bind into the SARS-CoV-2 main protease, essential for viral replication. J. Biomol. Struct. Dyn. 2021, 39, 3419–3427. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Zia, K.; Ashraf, S.; Uddin, R.; Ul-Haq, Z. Identification of chymotrypsin-like protease inhibitors of SARS-CoV-2 via integrated computational approach. J. Biomol. Struct. Dyn. 2021, 39, 2607–2616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, R.; Parves, R.; Paul, A.S.; Uddin, N.; Rahman, S.; Al Mamun, A.; Hossain, N.; Ali, A.; Halim, M.A. A molecular modeling approach to identify effective antiviral phytochemicals against the main protease of SARS-CoV-2. J. Biomol. Struct. Dyn. 2021, 39, 3213–3224. [Google Scholar] [CrossRef] [PubMed]
- Ton, A.T.; Gentile, F.; Hsing, M.; Ban, F.; Cherkasov, A. Rapid Identification of Potential Inhibitors of SARS-CoV-2 Main Protease by Deep Docking of 1.3 Billion Compounds. Mol. Inform. 2020, 39, e2000028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandeel, M.; Al-Nazawi, M. Virtual screening and repurposing of FDA approved drugs against COVID-19 main protease. Life Sci. 2020, 251, 117627. [Google Scholar] [CrossRef]
- Owen, D.R.; Allerton, C.M.N.; Anderson, A.S.; Aschenbrenner, L.; Avery, M.; Berritt, S.; Boras, B.; Cardin, R.D.; Carlo, A.; Coffman, K.J.; et al. An oral SARS-CoV-2 M(pro) inhibitor clinical candidate for the treatment of COVID-19. Science 2021, 374, 1586–1593. [Google Scholar] [CrossRef]
- Ahmad, B.; Batool, M.; Ain, Q.U.; Kim, M.S.; Choi, S. Exploring the Binding Mechanism of PF-07321332 SARS-CoV-2 Protease Inhibitor through Molecular Dynamics and Binding Free Energy Simulations. Int. J. Mol. Sci. 2021, 22, 9124. [Google Scholar] [CrossRef]
- Yang, K.S.; Leeuwon, S.Z.; Xu, S.; Liu, W.R. Evolutionary and Structural Insights about Potential SARS-CoV-2 Evasion of Nirmatrelvir. J. Med. Chem. 2022, 65, 8686–8698. [Google Scholar] [CrossRef]
- Nutho, B.; Mahalapbutr, P.; Hengphasatporn, K.; Pattaranggoon, N.C.; Simanon, N.; Shigeta, Y.; Hannongbua, S.; Rungrotmongkol, T. Why Are Lopinavir and Ritonavir Effective against the Newly Emerged Coronavirus 2019? Atomistic Insights into the Inhibitory Mechanisms. Biochemistry 2020, 59, 1769–1779. [Google Scholar] [CrossRef] [Green Version]
- Orkin, C.; DeJesus, E.; Khanlou, H.; Stoehr, A.; Supparatpinyo, K.; Lathouwers, E.; Lefebvre, E.; Opsomer, M.; Van de Casteele, T.; Tomaka, F. Final 192-week efficacy and safety of once-daily darunavir/ritonavir compared with lopinavir/ritonavir in HIV-1-infected treatment-naïve patients in the ARTEMIS trial. HIV Med. 2013, 14, 49–59. [Google Scholar] [CrossRef]
- Hariyanto, T.I.; Kristine, E.; Jillian Hardi, C.; Kurniawan, A. Efficacy of Lopinavir/Ritonavir Compared with Standard Care for Treatment of Coronavirus Disease 2019 (COVID-19): A Systematic Review. Infect. Disord. Drug Targets 2021, 21, e270421187364. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; De Clercq, E. Therapeutic options for the 2019 novel coronavirus (2019-nCoV). Nature reviews. Drug Discov. 2020, 19, 149–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abian, O.; Ortega-Alarcon, D.; Jimenez-Alesanco, A.; Ceballos-Laita, L.; Vega, S.; Reyburn, H.T.; Rizzuti, B.; Velazquez-Campoy, A. Structural stability of SARS-CoV-2 3CLpro and identification of quercetin as an inhibitor by experimental screening. Int. J. Biol. Macromol. 2020, 164, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Chung, T.D.; Oldenburg, K.R. A Simple Statistical Parameter for Use in Evaluation and Validation of High Throughput Screening Assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef]
- Zhu, M.; Du, X.N.; Li, Y.G.; Zhang, G.N.; Wang, J.X.; Wang, Y.C. Design, synthesis and biological evaluation of novel HIV-1 protease inhibitors with pentacyclic triterpenoids as P2-ligands. Bioorg. Med. Chem. Lett. 2019, 29, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Ma, L.; Zhou, H.; Dong, B.; Wang, Y.; Wang, Z.; Zhou, J.; Zhang, G.; Wang, J.; Liang, C.; et al. Preliminary SAR and biological evaluation of potent HIV-1 protease inhibitors with pyrimidine bases as novel P2 ligands to enhance activity against DRV-resistant HIV-1 variants. Eur. J. Med. Chem. 2020, 185, 111866. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Dong, B.; Zhang, G.N.; Wang, J.X.; Cen, S.; Wang, Y.C. Synthesis and biological evaluation of new HIV-1 protease inhibitors with purine bases as P2-ligands. Bioorg. Med. Chem. Lett. 2019, 29, 1541–1545. [Google Scholar] [CrossRef]
- Zhu, M.; Dou, Y.; Ma, L.; Dong, B.; Zhang, F.; Zhang, G.; Wang, J.; Zhou, J.; Cen, S.; Wang, Y. Novel HIV-1 Protease Inhibitors with Morpholine as the P2 Ligand to Enhance Activity against DRV-Resistant Variants. ACS Med. Chem. Lett. 2020, 11, 1196–1204. [Google Scholar] [CrossRef]
- Zhu, M.; Ma, L.; Wen, J.; Dong, B.; Wang, Y.; Wang, Z.; Zhou, J.; Zhang, G.; Wang, J.; Guo, Y.; et al. Rational design and Structure-Activity relationship of coumarin derivatives effective on HIV-1 protease and partially on HIV-1 reverse transcriptase. Eur. J. Med. Chem. 2020, 186, 111900. [Google Scholar] [CrossRef]
- Zhu, M.; Ma, L.; Dong, B.; Zhang, G.; Wang, J.; Zhou, J.; Cen, S.; Wang, Y. Synthesis and evaluation of potent human immunodeficiency virus 1 protease inhibitors with epimeric isopropanol as novel P1’ ligands. Future Med. Chem. 2020, 12, 775–794. [Google Scholar] [CrossRef]
- Zhu, M.; Ma, L.; Dong, B.; Zhang, G.; Wang, J.; Zhou, J.; Cen, S.; Wang, Y. Design and biological evaluation of novel HIV-1 protease inhibitors with isopropanol as P1’ ligand to enhance binding with S1’ subsite. Bioorg. Med. Chem. 2020, 28, 115623. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-M.; Yang, W.-L.; Yang, F.-Y.; Zhang, L.; Huang, W.-J.; Hou, W.; Fan, C.-F.; Jin, R.-H.; Feng, Y.-M.; Wang, Y.-C.; et al. Cathepsin L plays a key role in SARS-CoV-2 infection in humans and humanized mice and is a promising target for new drug development. Signal Transduct. Target. Ther. 2021, 6, 134. [Google Scholar] [CrossRef]
- Ma, L.; Li, Q.; Xie, Y.; Zhao, J.; Yi, D.; Guo, S.; Guo, F.; Wang, J.; Yang, L.; Cen, S. Repurposing of HIV/HCV protease inhibitors against SARS-CoV-2 3CLpro. Antivir. Res. 2022, 207, 105419. [Google Scholar] [CrossRef]
- Aoki, M.; Hayashi, H.; Yedidi, R.S.; Martyr, C.D.; Takamatsu, Y.; Aoki-Ogata, H.; Nakamura, T.; Nakata, H.; Das, D.; Yamagata, Y.; et al. C-5-Modified Tetrahydropyrano-Tetrahydofuran-Derived Protease Inhibitors (PIs) Exert Potent Inhibition of the Replication of HIV-1 Variants Highly Resistant to Various PIs, including Darunavir. J. Virol. 2015, 90, 2180–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, E.C.; Goldstein, S.A.; Rasmussen, A.L.; Robertson, D.L.; Crits-Christoph, A.; Wertheim, J.O.; Anthony, S.J.; Barclay, W.S.; Boni, M.F.; Doherty, P.C.; et al. The origins of SARS-CoV-2: A critical review. Cell 2021, 184, 4848–4856. [Google Scholar] [CrossRef] [PubMed]
- Simonis, A.; Theobald, S.J.; Fätkenheuer, G.; Rybniker, J.; Malin, J.J. A comparative analysis of remdesivir and other repurposed antivirals against SARS-CoV-2. EMBO Mol. Med. 2021, 13, e13105. [Google Scholar] [CrossRef]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir-Ritonavir in Adults Hospitalized with Severe Covid-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef]
- Abdelrahman, Z.; Li, M.; Wang, X. Comparative Review of SARS-CoV-2, SARS-CoV, MERS-CoV, and Influenza A Respiratory Viruses. Front. Immunol. 2020, 11, 552909. [Google Scholar] [CrossRef]
- Li, Q.; Yi, D.; Lei, X.; Zhao, J.; Zhang, Y.; Cui, X.; Xiao, X.; Jiao, T.; Dong, X.; Zhao, X.; et al. Corilagin inhibits SARS-CoV-2 replication by targeting viral RNA-dependent RNA polymerase. Acta Pharm. Sin. B 2021, 11, 1555–1567. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

| Type in P2 Ligands | R1 | R2 | R3 | |
|---|---|---|---|---|
| Pentacyclic triterpenoid |  | Ra = H, or CH3 | CH3 | OCH3, NO2, or NH2 |
| Purine |  | Ra = OCH3, H, Cl or NH2 | CH3 | OCH3, NO2, or NH2 |
| Pyrimidine |  | Ra = O, 2H, or NH2 | CH3 | OCH3, NO2, or NH2 |
| Morpholine |  | X = O, or SO2; n = 1, or 2 | CH3 | OCH3, NH2, NHOH, NO2, or CF3 |
| Piperidine |  | X = CH2, or O; n = 0, or 1 | CH3 | OCH3, or NH2 |
| Coumarin |  | Ra = H, CH3, NH2, OH, or OCH3 | CH3 | OCH3, NO2, or NH2 |
| Phenol |  | Ra = H, OCH3, OH, or Cl | CH3, (R)-OH, or (S)-OH | OCH3, NO2, NH2, F, or CF3 |
| Phenylpropionic acid |  | Ra = H, OCH3, or OH | H, or CH3 | OCH3, SCH3, or NH2 |
| Analyte | Ligand (3CLpro) | |||||
|---|---|---|---|---|---|---|
| Kon* (1/Ms) | Kon* Error | Kdis* (1/s) | Kdis* Error | KD* (M) | KD* Error | |
| 29# | 3.74 × 102 | 16.3 | 1.97 × 10−2 | 6.01 × 10−4 | 5.27 × 10−5 | 2.81 × 10−6 |
| 50# | 4.55 × 102 | 18.3 | 2.82 × 10−2 | 6.86 × 10−4 | 6.21 × 10−5 | 2.92 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Xie, Y.; Zhu, M.; Yi, D.; Zhao, J.; Guo, S.; Zhang, Y.; Wang, J.; Li, Q.; Wang, Y.; et al. Identification of Darunavir Derivatives for Inhibition of SARS-CoV-2 3CLpro. Int. J. Mol. Sci. 2022, 23, 16011. https://doi.org/10.3390/ijms232416011
Ma L, Xie Y, Zhu M, Yi D, Zhao J, Guo S, Zhang Y, Wang J, Li Q, Wang Y, et al. Identification of Darunavir Derivatives for Inhibition of SARS-CoV-2 3CLpro. International Journal of Molecular Sciences. 2022; 23(24):16011. https://doi.org/10.3390/ijms232416011
Chicago/Turabian StyleMa, Ling, Yongli Xie, Mei Zhu, Dongrong Yi, Jianyuan Zhao, Saisai Guo, Yongxin Zhang, Jing Wang, Quanjie Li, Yucheng Wang, and et al. 2022. "Identification of Darunavir Derivatives for Inhibition of SARS-CoV-2 3CLpro" International Journal of Molecular Sciences 23, no. 24: 16011. https://doi.org/10.3390/ijms232416011
APA StyleMa, L., Xie, Y., Zhu, M., Yi, D., Zhao, J., Guo, S., Zhang, Y., Wang, J., Li, Q., Wang, Y., & Cen, S. (2022). Identification of Darunavir Derivatives for Inhibition of SARS-CoV-2 3CLpro. International Journal of Molecular Sciences, 23(24), 16011. https://doi.org/10.3390/ijms232416011