
The Regulation of Nitrate Reductases in Response to Abiotic Stress in Arabidopsis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result
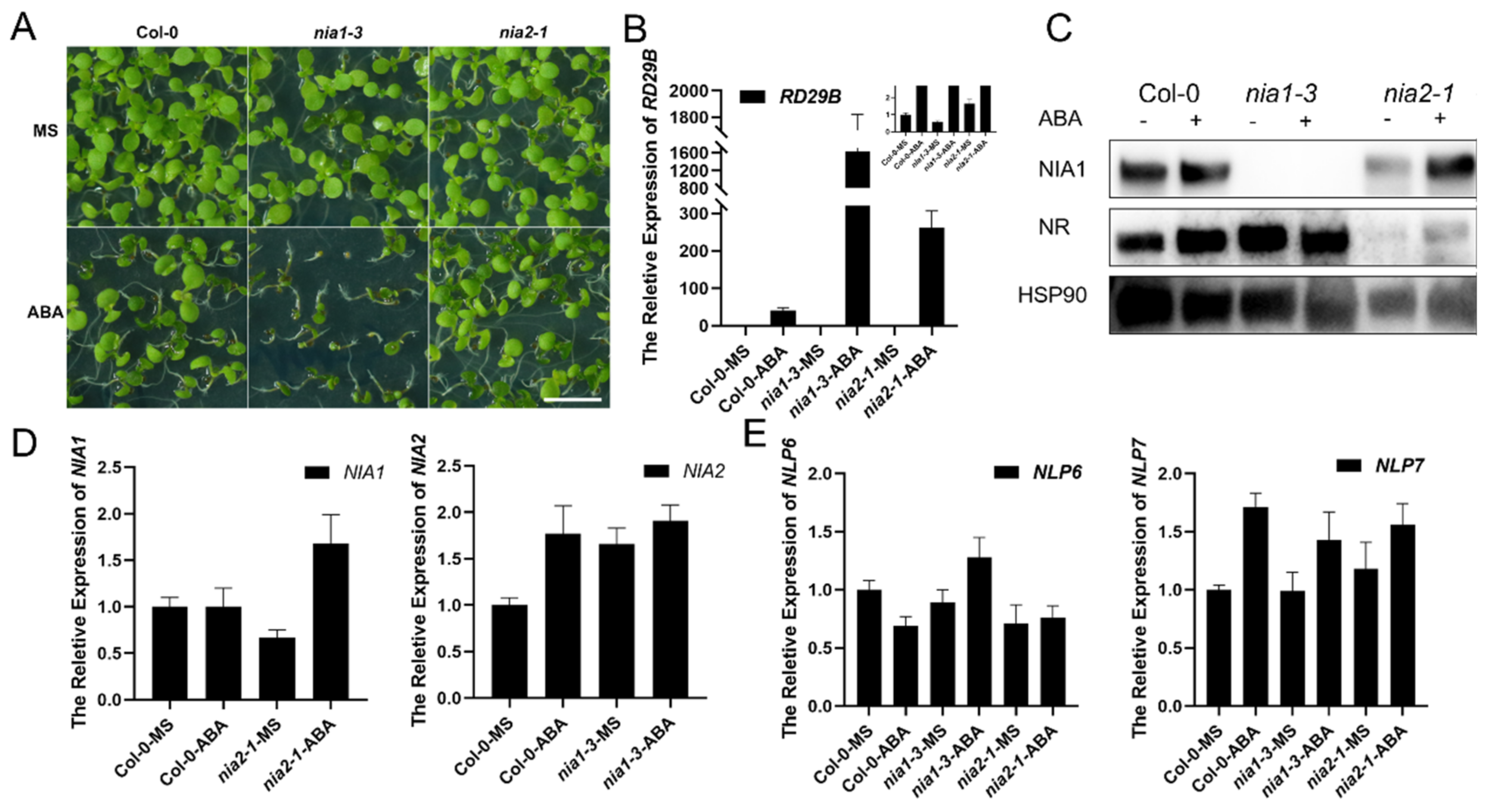
2.1. Mutation of NIA1 Is Hypersensitive under ABA Treatment
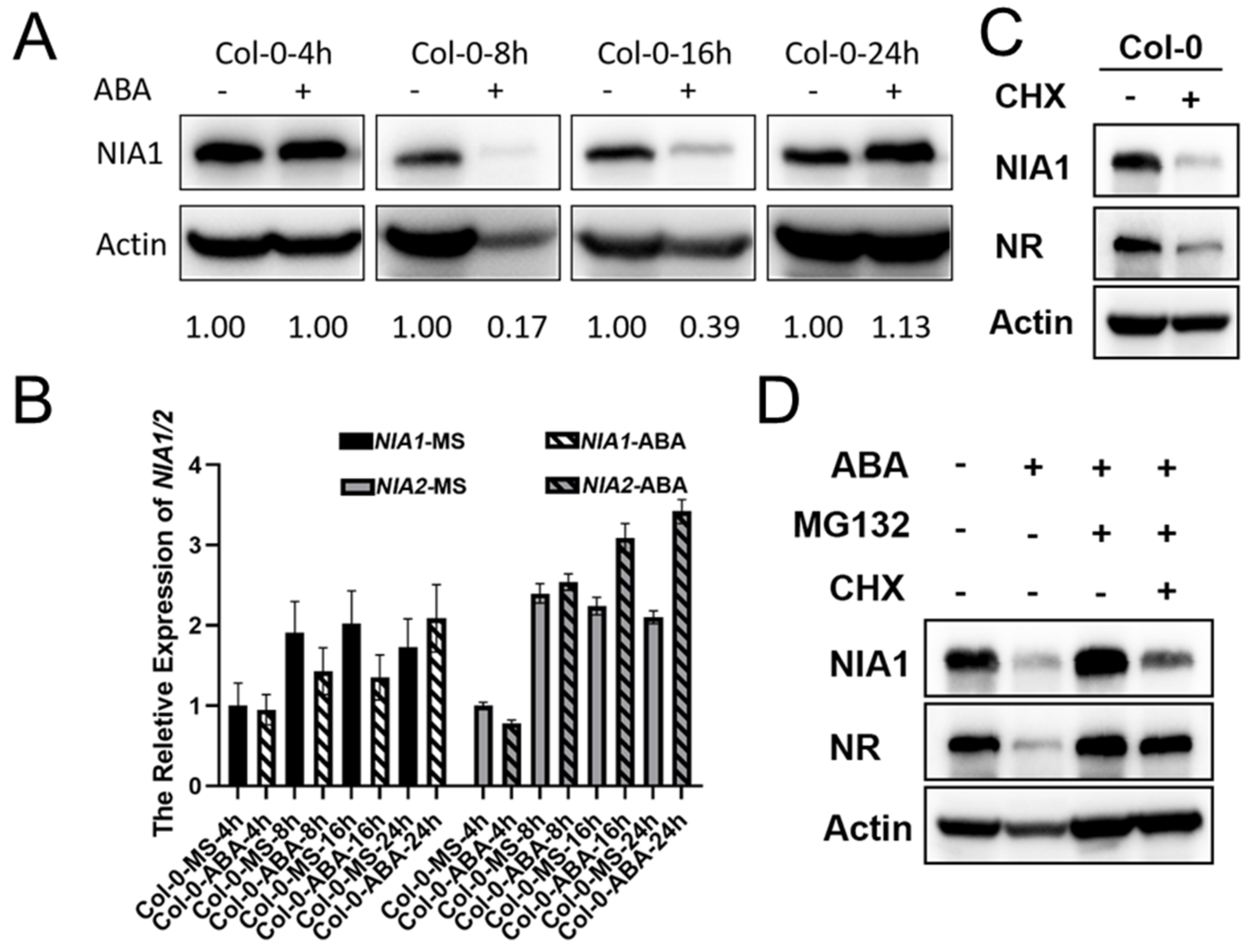
2.2. ABA Treatment Rapidly Regulates the Abundance of NR Proteins via a 26S Proteasome Manner
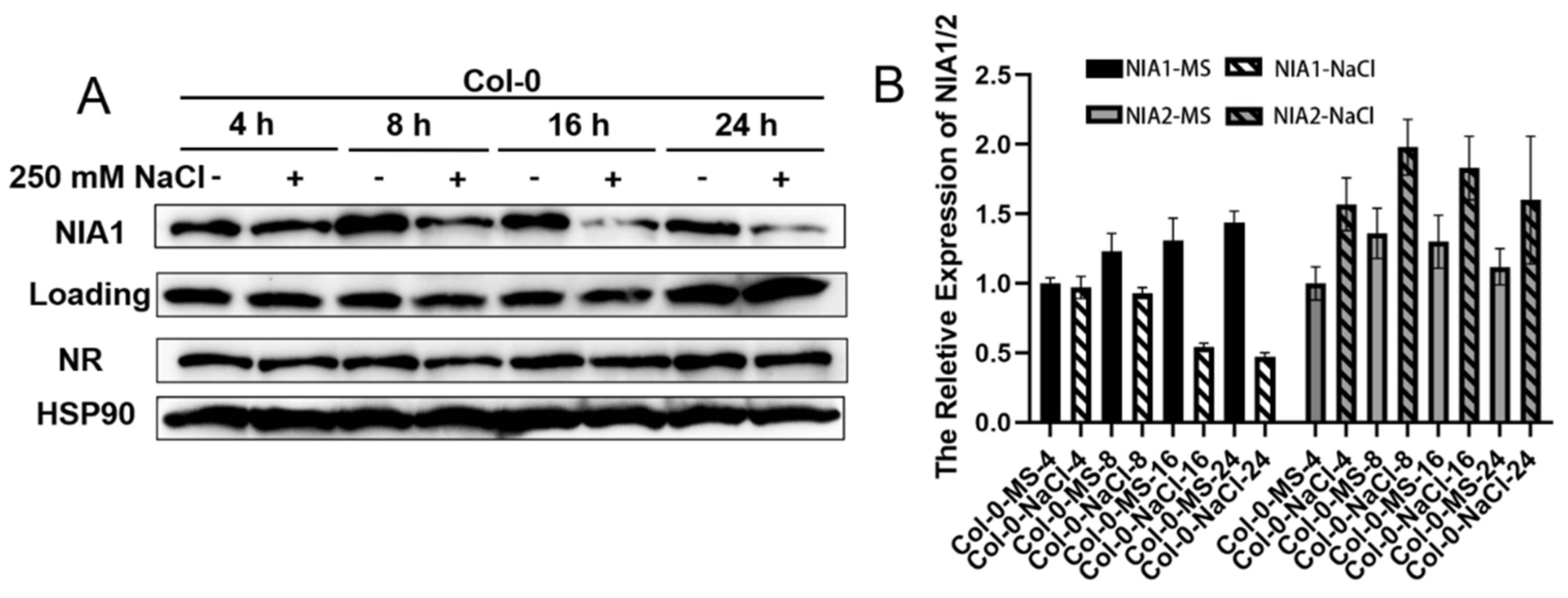
2.3. Single Mutation of NIA1 or NIA2 Shows a Similar Phenotype under Long-Term NaCl Stress
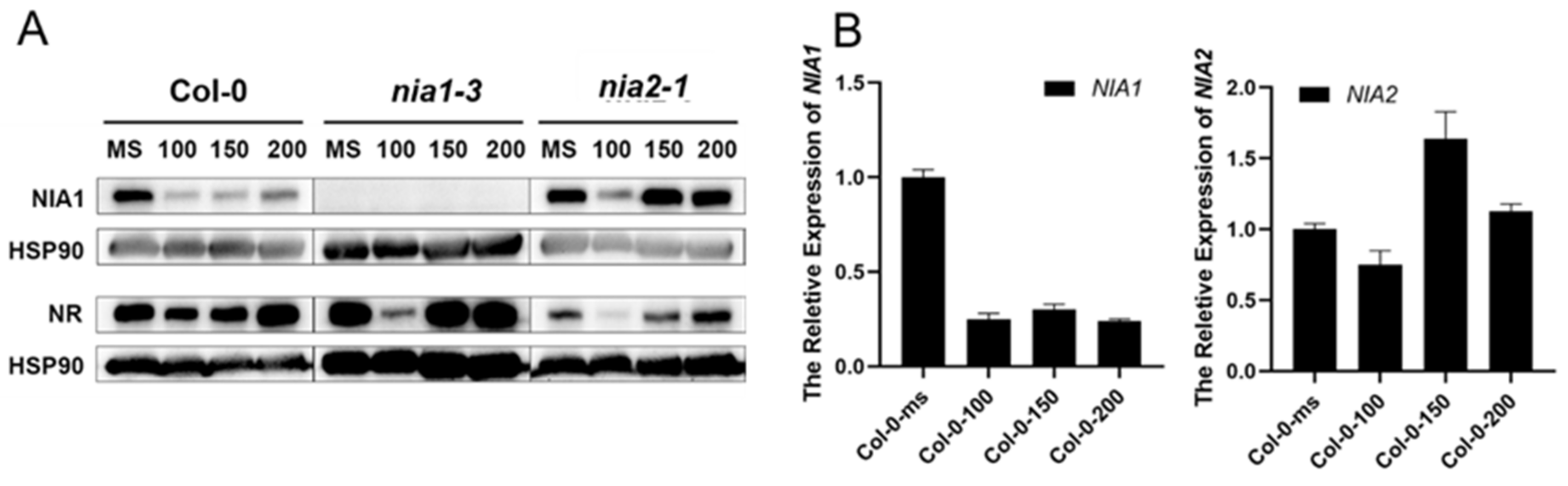
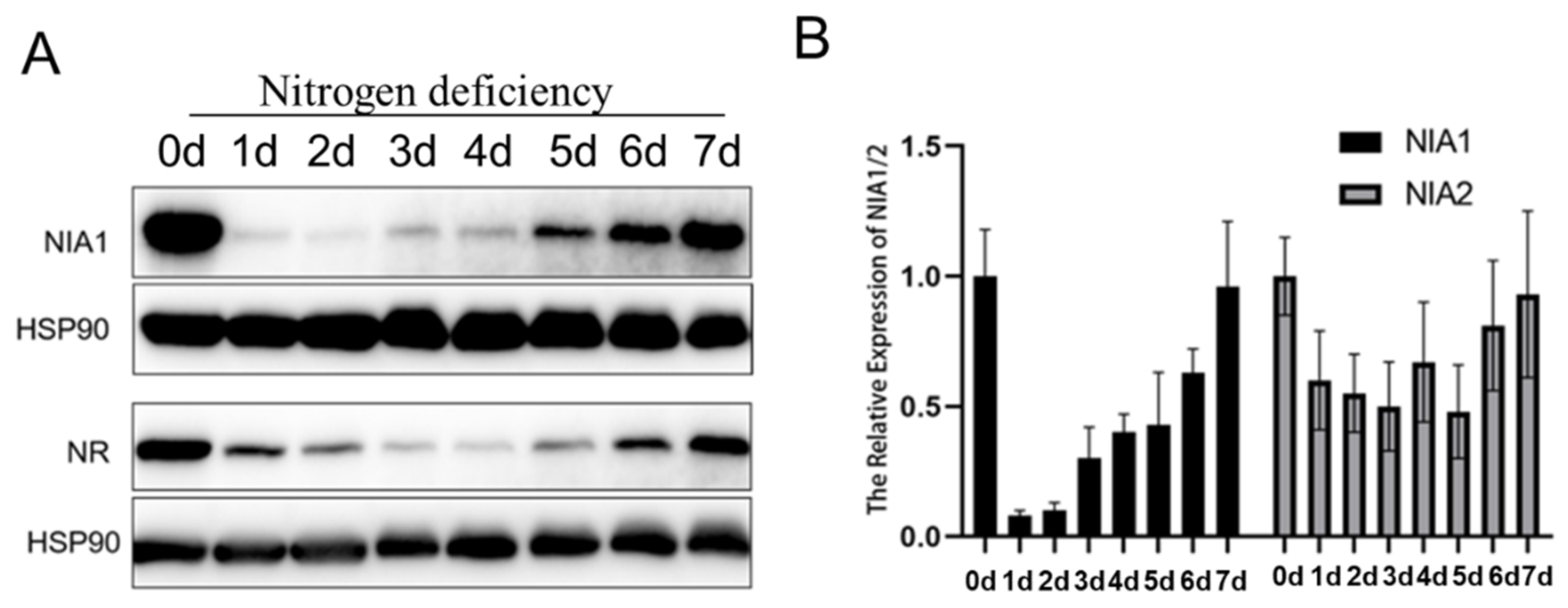
2.4. Nitrogen Deficiency Stress Also Dynamically Downregulates Both NIA1 and NIA2 in the Transcriptional Level
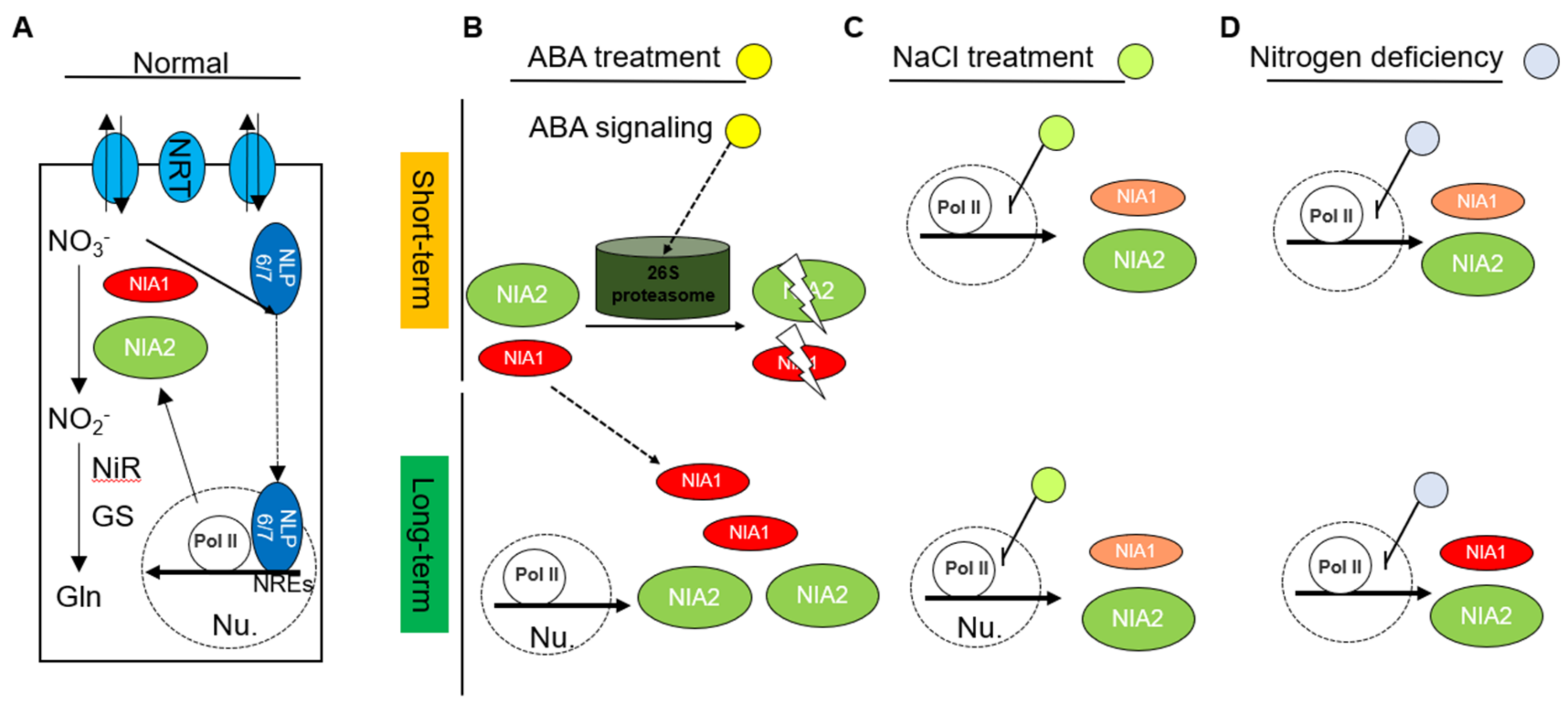
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Growth Condition
4.3. Gene Expression Analysis by Quantitative PCR
4.4. Total Protein Extraction and Immunoblot Assays
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant. Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef]
- Maathuis, F.J. Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef]
- Miller, A.J.; Cramer, M.D. Root nitrogen acquisition and assimilation. Plant Soil 2005, 274, 1–36. [Google Scholar] [CrossRef]
- Jämtgård, S.; Näsholm, T.; Huss-danell, K. Characteristics of amino acid uptake in barley. Plant Soil 2007, 302, 221–231. [Google Scholar] [CrossRef]
- Crawford, N.M. Nitrate: Nutrient and signal for plant growth. Plant Cell 1995, 7, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krouk, G.; Crawford, N.M.; Coruzzi, G.M. Nitrate signaling: Adaptation to fluctuating environments. Curr. Opin. Plant Biol. 2010, 13, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, S. Transcription factors involved in controlling the expression of nitrate reductase genes in higher plants. Plant Sci. 2014, 229, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Konishi, M.; Yanagisawa, S.; Omata, T. Nitrite transport activity of a novel HPP family protein conserved in cyanobacteria and chloroplasts. Plant Cell Physiol. 2014, 55, 1311–1324. [Google Scholar] [CrossRef] [Green Version]
- Hoff, T.; Truong, H.N.; Caboche, M. The use of mutants and transgenic plants to study nitrate assimilation. Plant Cell Environ. 1994, 17, 489–506. [Google Scholar] [CrossRef]
- Wilkinson, J.Q.; Crawford, N.M. Identification of the Arabidopsis CHL3 gene as the nitrate reductase structural gene NIA2. Plant Cell 1991, 3, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, J.Q.; Crawford, N.M. Identification and characterization of a chlorate-resistant mutant of Arabidopsis thaliana with mutations in both nitrate reductase structural genes NIA1 and NIA2. Mol. Gen. Genet. 1993, 239, 289–297. [Google Scholar] [CrossRef]
- Galangau, F.; Daniel-Vedele, F.; Moureaux, T.; Dorbe, M.F.; Leydecker, M.T.; Caboche, M. Expression of leaf nitrate reductase genes from tomato and tobacco in relation to light-dark regimes and nitrate supply. Plant Physiol. 1988, 88, 383–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Else, M.J.; Lea, U.S.; Lillo, C. HY5 and HYH are positive regulators of nitrate reductase in seedlings and rosette stage plants. Planta 2008, 227, 559–564. [Google Scholar] [CrossRef]
- Yang, Z.J.; Midmore, D.J. A model for the circadian oscillations in expression and activity of nitrate reductase in higher plants. Ann. Bot. 2005, 6, 1019–1026. [Google Scholar] [CrossRef] [Green Version]
- Lejay, L.; Quillere, I.; Roux, Y.; Tillard, P.; Cliquet, J.B.; Meyer, C. Abolition of posttranscriptional regulation of nitrate reductase partially prevents the decrease in leaf NO3− reduction when photosynthesis is inhibited by CO2 deprivation, but not in darkness. Plant Physiol. 1997, 115, 623–631. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Cai, S.; Wang, Y.; Chen, Z.H. Loss of nitrate reductases NIA1 and NIA2 impairs stomatal closure by altering genes of core aba signaling components in Arabidopsis. Plant Signal. Behav. 2016, 11, 1456–1469. [Google Scholar] [CrossRef] [Green Version]
- Tian, Q.Y.; Sun, P.; Zhang, W.H. Ethylene is involved in nitrate-dependent root growth and branching in Arabidopsis thaliana. New Phytol. 2009, 184, 918–931. [Google Scholar] [CrossRef]
- Lu, J.L.; Ertl, J.R.; Chen, C.M. Transcriptional regulation of nitrate reductase mRNA levels by cytokinin-abscisic acid interactions in etiolated barley leaves. Plant Physiol. 1992, 98, 1255–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, M.; Yanagisawa, S. Arabidopsis NIN-like transcription factors have a central role in nitrate signalling. Nat. Commun. 2013, 4, 1617. [Google Scholar] [CrossRef] [Green Version]
- Castaings, L.; Camargo, A.; Pocholle, D.; Gaudon, V.; Texier, Y.; Boutet-Mercey, S.; Taconnat, L.; Renou, J.P.; Daniel-Vedele, F.; Fernandez, E.; et al. The nodule inception-like protein 7 modulates nitrate sensing and metabolism in Arabidopsis. Plant J. 2009, 57, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Schauser, L.; Wieloch, W.; Stougaard, J. Evolution of NIN-like proteins in Arabidopsis, rice, and Lotus japonicus. J. Mol.Evol. 2005, 60, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Marchive, C.; Roudier, F.; Castaings, L.; Bréhaut, V.; Blondet, E.; Colot, V.; Meyer, C.; Krapp, A. Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat. Commun. 2013, 4, 1713. [Google Scholar] [CrossRef] [PubMed]
- Park, B.S.; Song, J.T.; Seo, H.S. Nitrate reductase activity is stimulated by the E3 sumo ligase AtSIZ1. Nat. Commun. 2011, 2, 400. [Google Scholar] [CrossRef] [Green Version]
- Su, W.; Huber, S.C.; Crawford, N.M. Identification in vitro of a post-translational regulatory site in the hinge 1 region of Arabidopsis nitrate reductase. Plant Cell 1996, 8, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKintosh, C.; Meek, S.E.M. Regulation of plant NR activity by reversible phosphorylation, 14-3-3 proteins and proteolysis. Cell. Mol. Life Sci. 2001, 58, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, W.M.; Brendle-Behnisch, E. Rapid modulation of spinach leaf nitrate reductase activity by photosynthesis I. Modulation in vivo by CO2 availability. Plant Physiol. 1991, 96, 363–367. [Google Scholar] [CrossRef] [Green Version]
- Huber, J.L.; Huber, S.C.; Campbell, W.H.; Redinbaugh, M.G. Reversible light/dark modulation of spinach leaf nitrate reductase activity involves protein phosphorylation. Arch Biochem. Biophys. 1992, 296, 58–65. [Google Scholar] [CrossRef]
- Huber, J.L.; Redinbaugh, M.G.; Huber, S.C.; Campbell, W.H. Regulation of maize leaf nitrate reductase activity involves both gene expression and protein phosphorylation. Plant Physiol. 1994, 106, 1667–1674. [Google Scholar] [CrossRef] [Green Version]
- Weiner, H.; Kaiser, W.M. 14-3-3 proteins control proteolysis of nitrate reductase in spinach leaves. FEBS Lett. 1994, 455, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Joo, K.; Bong, P.; Sang, P.; Han, L.; Song, J.; Hak, S. Nitrate reductases are relocalized to the nucleus by AtSIZ1 and their levels are negatively regulated by cop1 and ammonium. Int. J. Mol. Sci. 2018, 19, 1202. [Google Scholar] [CrossRef] [Green Version]
- Jonassen, E.M.; Sévin, D.C.; Lillo, C. The bZIP transcription factors HY5 and HYH are positive regulators of the main nitrate reductase gene in Arabidopsis leaves, NIA2, but negative regulators of the nitrate uptake gene NRT1.1. J. Plant Physiol. 2009, 166, 2071–2076. [Google Scholar] [CrossRef]
- Wilson, I.D.; Ribeiro, D.M.; Bright, J.; Confraria, A.; Harrison, J.; Barros, R.S.; Desikan, R.; Neill, S.J.; Hancock, J.T. Role of nitric oxide in regulating stomatal apertures. Plant Signal. Behav. 2009, 4, 467–469. [Google Scholar] [CrossRef] [Green Version]
- Hussain, Q.; Asim, M.; Zhang, R.; Khan, R.; Farooq, S.; Wu, J.S. Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress. Biomolecules 2021, 11, 1159. [Google Scholar] [CrossRef] [PubMed]
- Roychoudhury, A.; Paul, S.; Basu, S. Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress. Plant Cell Rep. 2013, 32, 985–1006. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, X.; Zhang, K.; An, H.; Hu, K.; Wen, J.; Shen, J.; Ma, C.; Yi, B.; Tu, J.; et al. Comparative analysis of the Brassica napus root and leaf transcript profiling in response to drought stress. Int. J. Mol. Sci. 2015, 16, 18752–18777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Guegler, K.; LaBrie, S.T.; Crawford, N.M. Genomic analysis of a nutrient response in Arabidopsis reveals diverse expression patterns and novel metabolic and potential regulatory genes that are induced by nitrate. Plant Cell 2000, 12, 1491–1510. [Google Scholar] [CrossRef]
- Chen, K.; Guo, J.; Bressan, A.R.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.C.; Zhao, Y.; Li, Z.P.; Hsu, C.C.; Liu, X.; Fu, L.W.; Hou, Y.J.; Du, Y.Y.; Xie, S.J.; Zhang, C.J.; et al. Reciprocal Regulation of the TOR Kinase and ABA Receptor Balances Plant Growth and Stress Response. Mol. Cell 2018, 69, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate Reductase Regulates Plant Nitric Oxide Homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef]
- Wu, H.H.; Li, B.S.; Iwakawa, H.O.; Pan, Y.J.; Tang, X.L.; Ling-Hu, Q.Y.; Liu, Y.L.; Sheng, S.X.; Feng, L.; Zhang, H.; et al. Plant 22-nt siRNAs mediate translational repression and stress adaptation. Nature 2020, 581, 89–93. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, X.; Peng, Y.; Li, Z.; Guo, H.; Xia, X.; Li, B.; Yin, W. The Regulation of Nitrate Reductases in Response to Abiotic Stress in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 1202. https://doi.org/10.3390/ijms23031202
Tang X, Peng Y, Li Z, Guo H, Xia X, Li B, Yin W. The Regulation of Nitrate Reductases in Response to Abiotic Stress in Arabidopsis. International Journal of Molecular Sciences. 2022; 23(3):1202. https://doi.org/10.3390/ijms23031202
Chicago/Turabian StyleTang, Xianli, Yang Peng, Zheng Li, Hongwei Guo, Xinli Xia, Bosheng Li, and Weilun Yin. 2022. "The Regulation of Nitrate Reductases in Response to Abiotic Stress in Arabidopsis" International Journal of Molecular Sciences 23, no. 3: 1202. https://doi.org/10.3390/ijms23031202
APA StyleTang, X., Peng, Y., Li, Z., Guo, H., Xia, X., Li, B., & Yin, W. (2022). The Regulation of Nitrate Reductases in Response to Abiotic Stress in Arabidopsis. International Journal of Molecular Sciences, 23(3), 1202. https://doi.org/10.3390/ijms23031202