
Male Rat Offspring Are More Impacted by Maternal Obesity Induced by Cafeteria Diet than Females—Additive Effect of Postweaning Diet

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
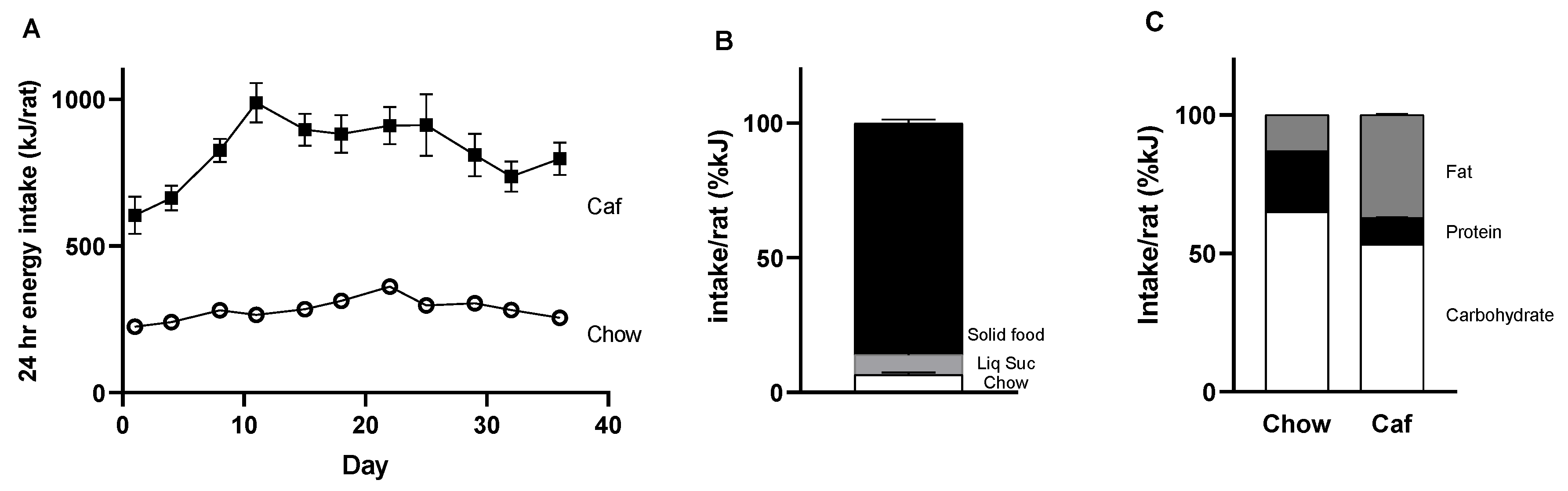
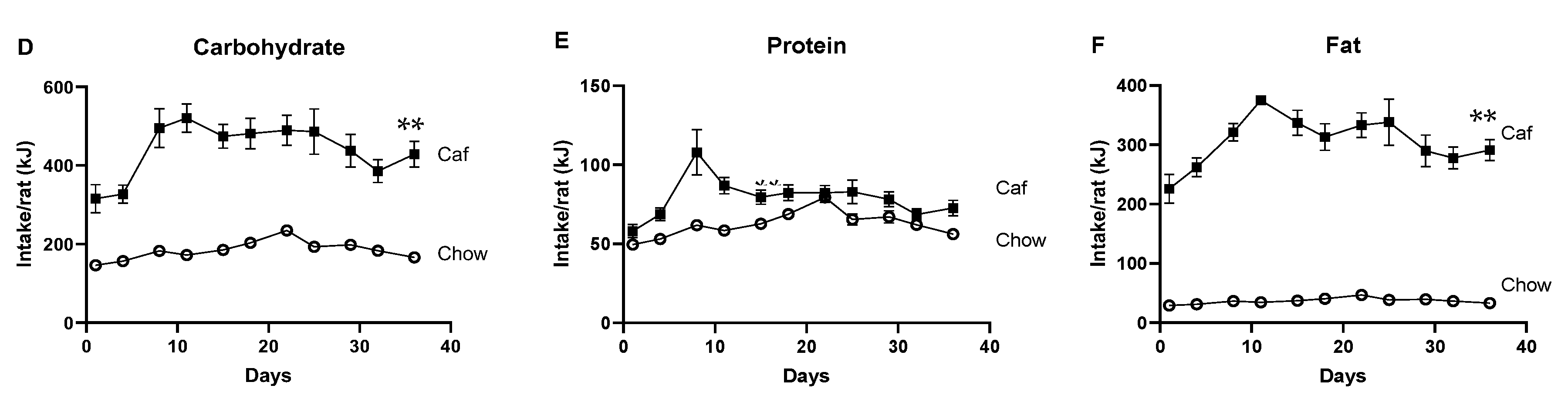
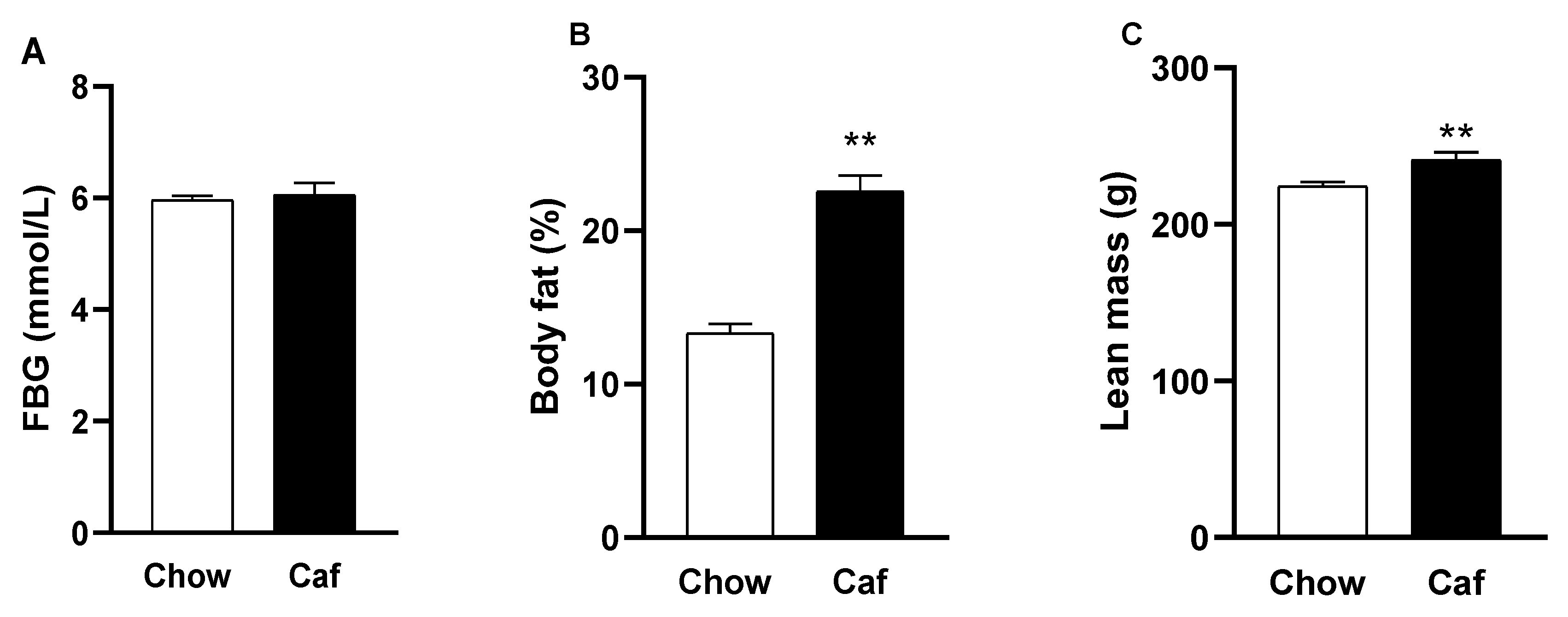
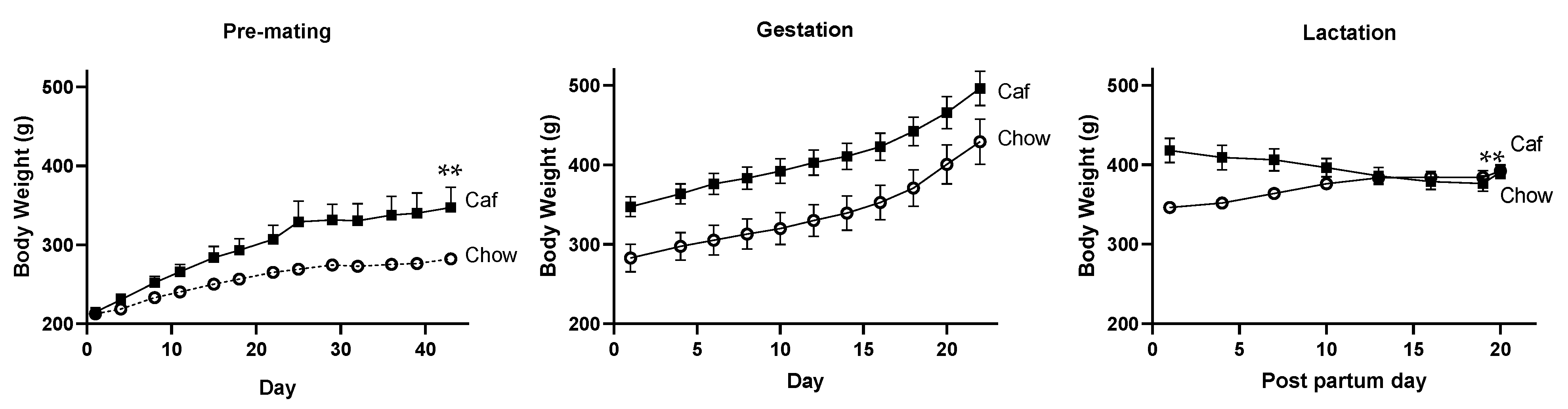
2.1. Dams
2.1.1. Food Intake of Dams
2.1.2. Fasting Blood Glucose and Echo-MRI
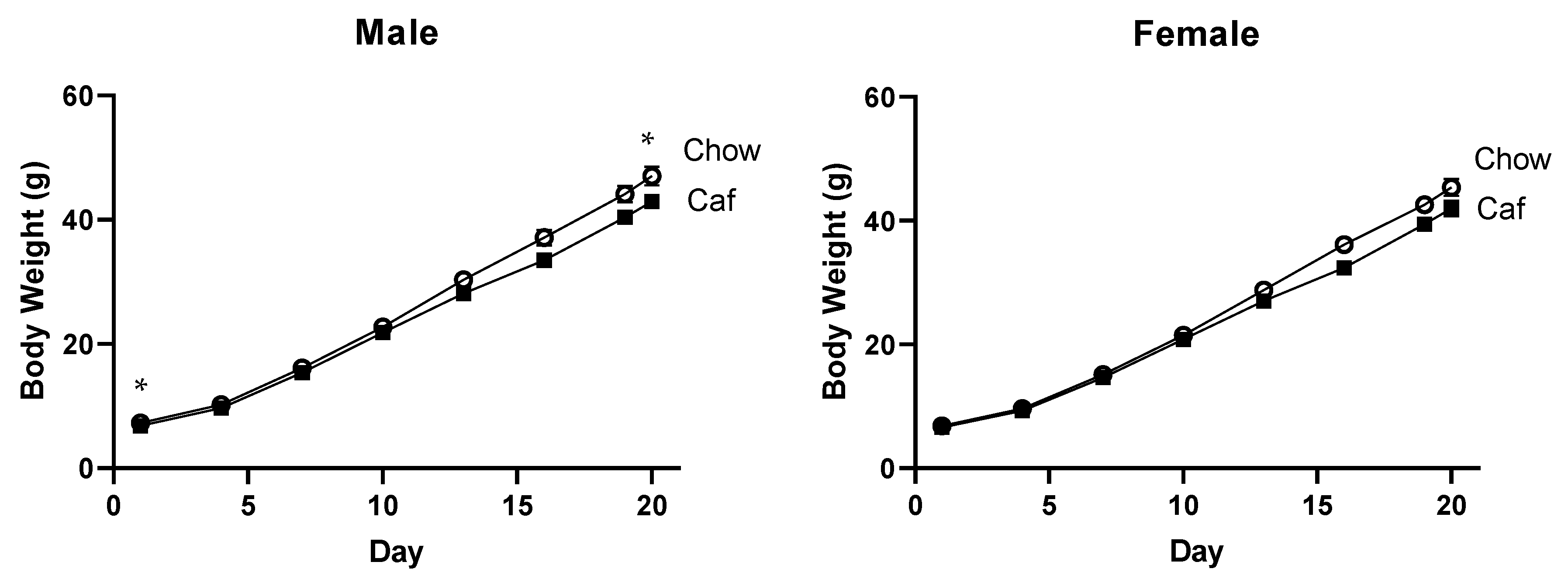
2.1.3. Body Weight
2.1.4. Birth Characteristics
2.1.5. Dam Endpoint Measures
2.2. Offspring
2.2.1. Pre-Weaning
2.2.2. Post-Weaning Measures
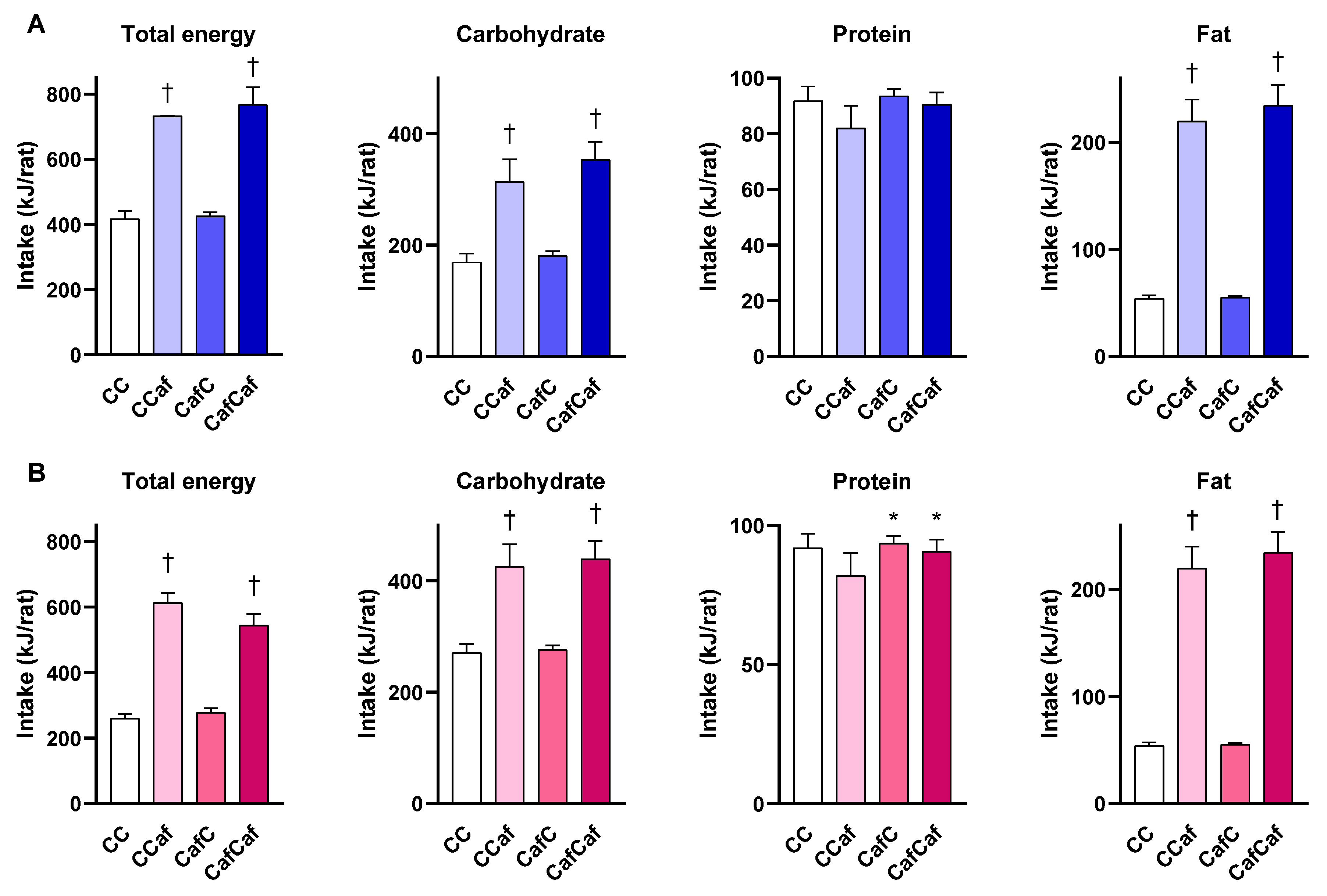
Food Intake
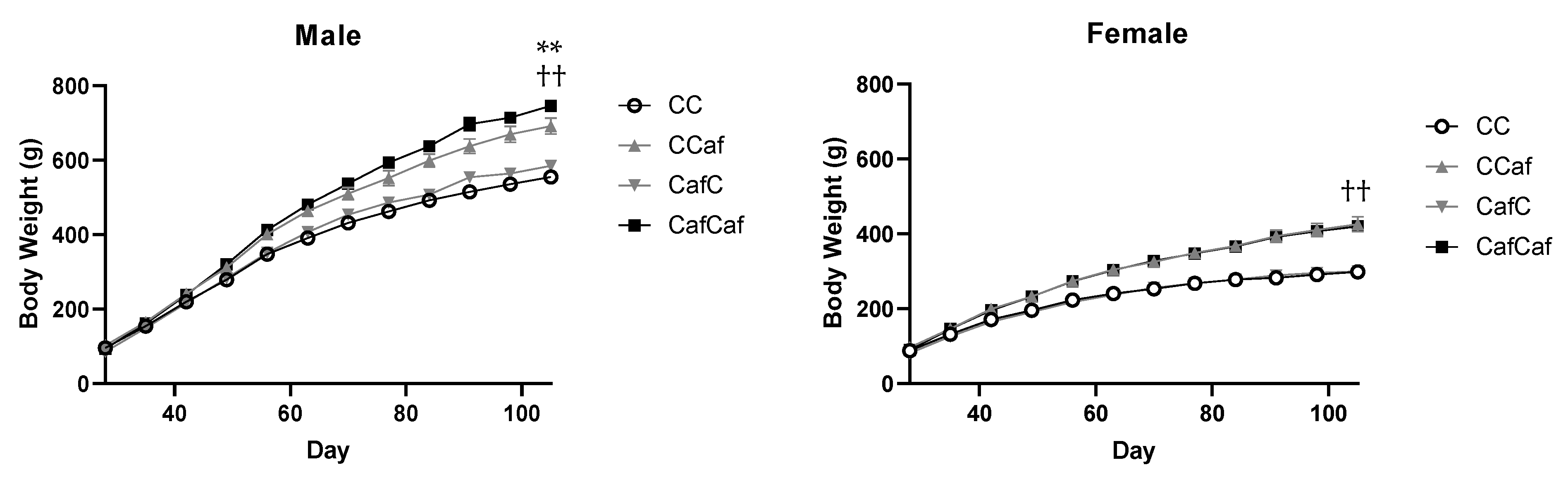
Body Weight
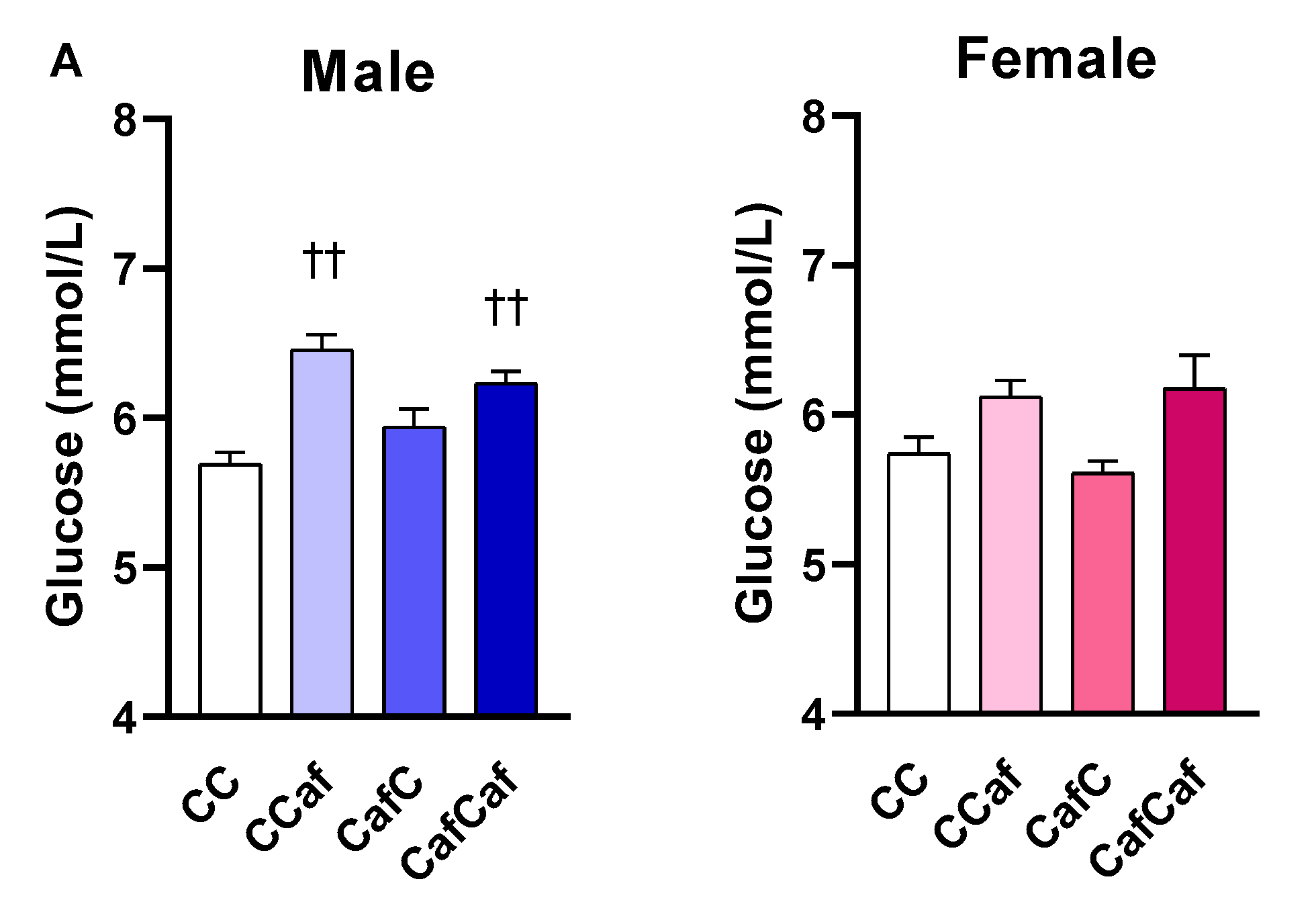
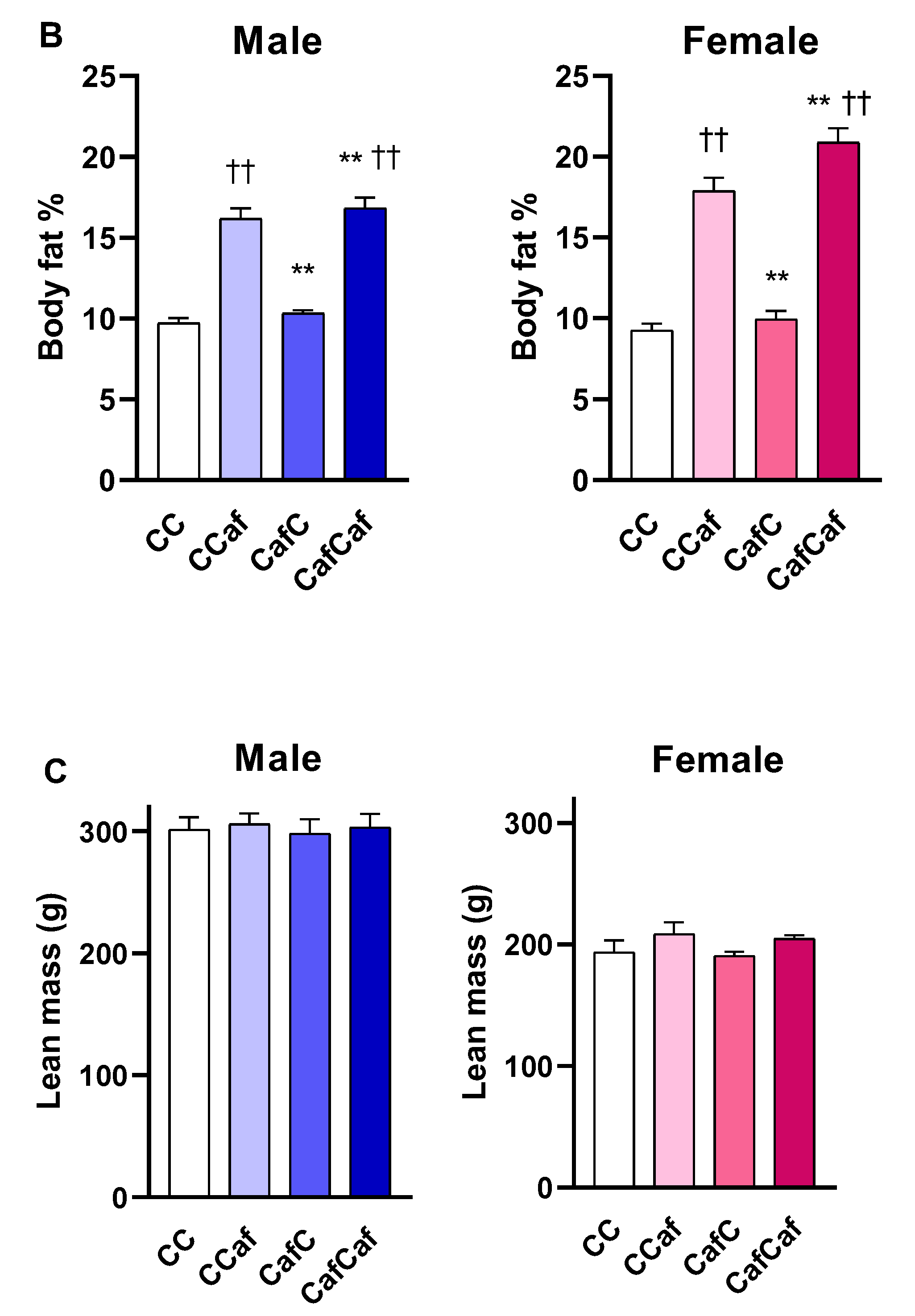
Fasting Blood Glucose and Echo-MRI
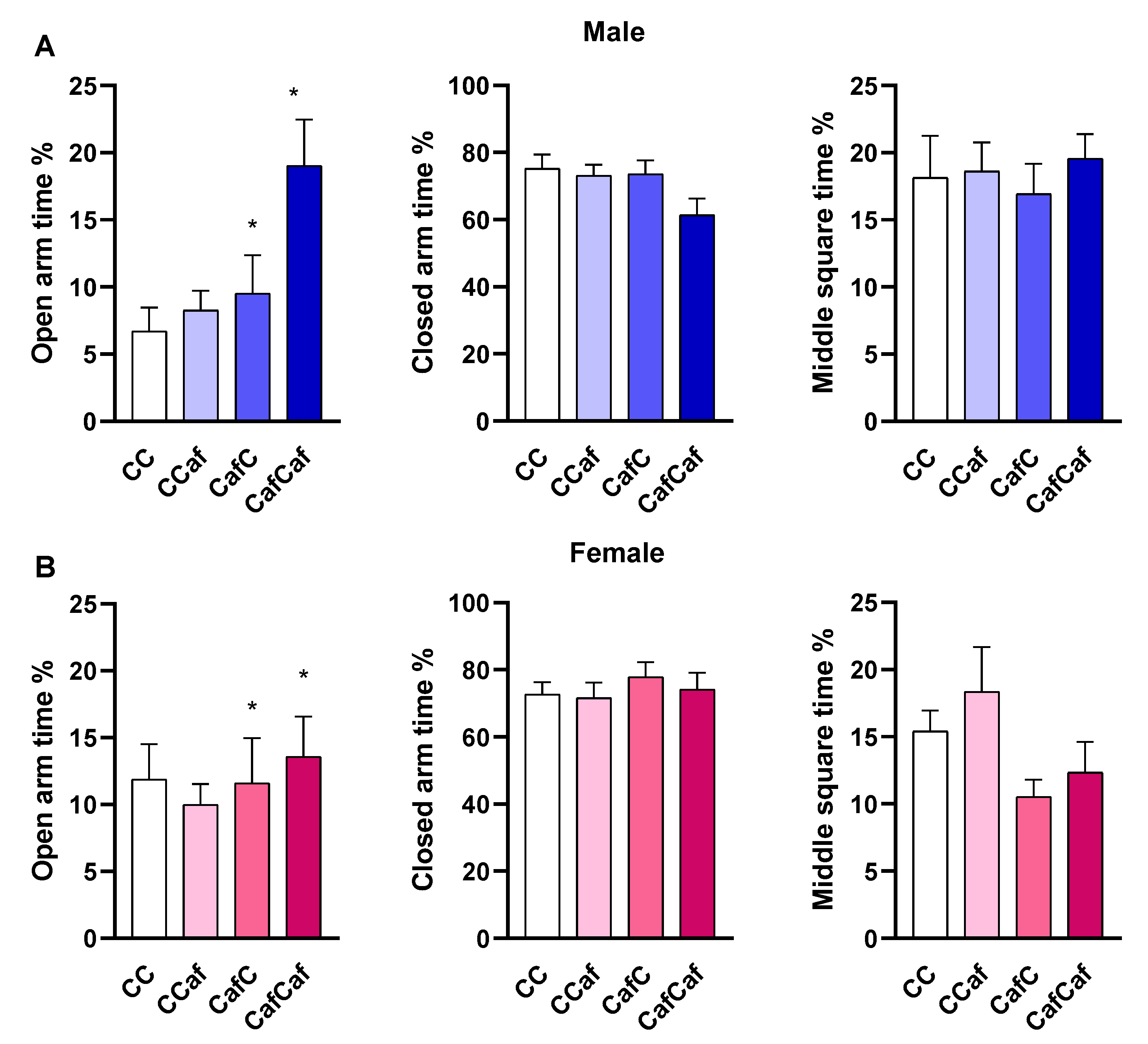
Elevated Plus Maze (EPM) Behaviour
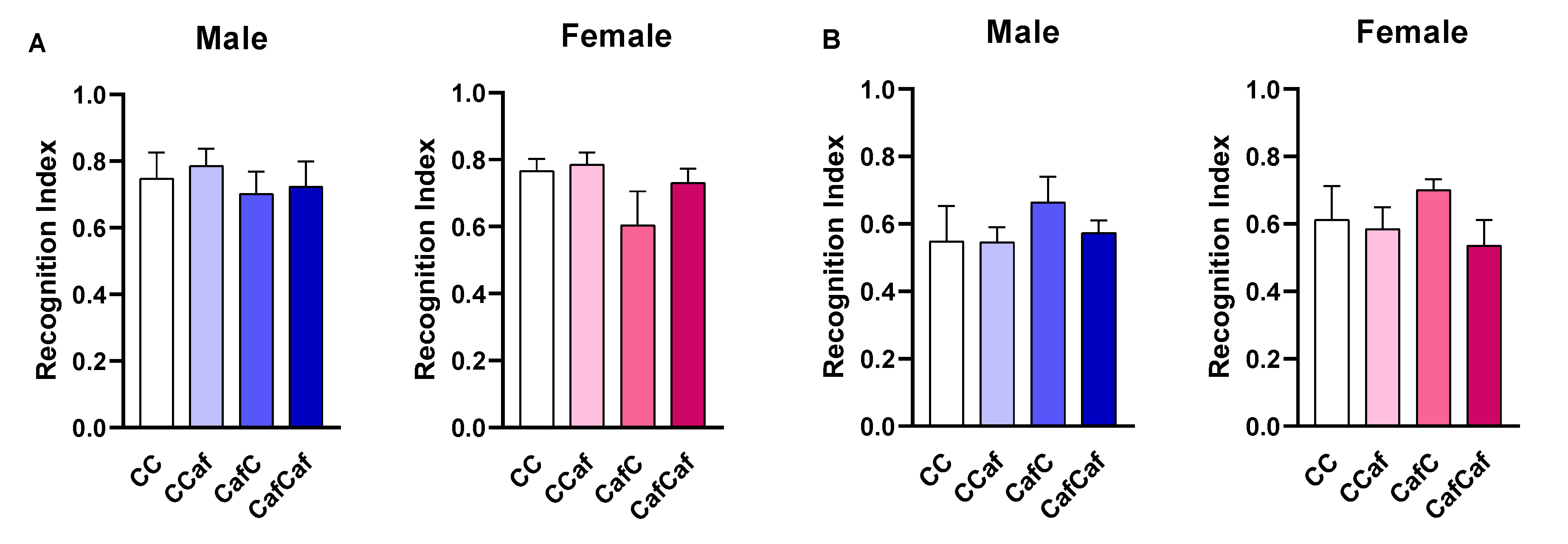
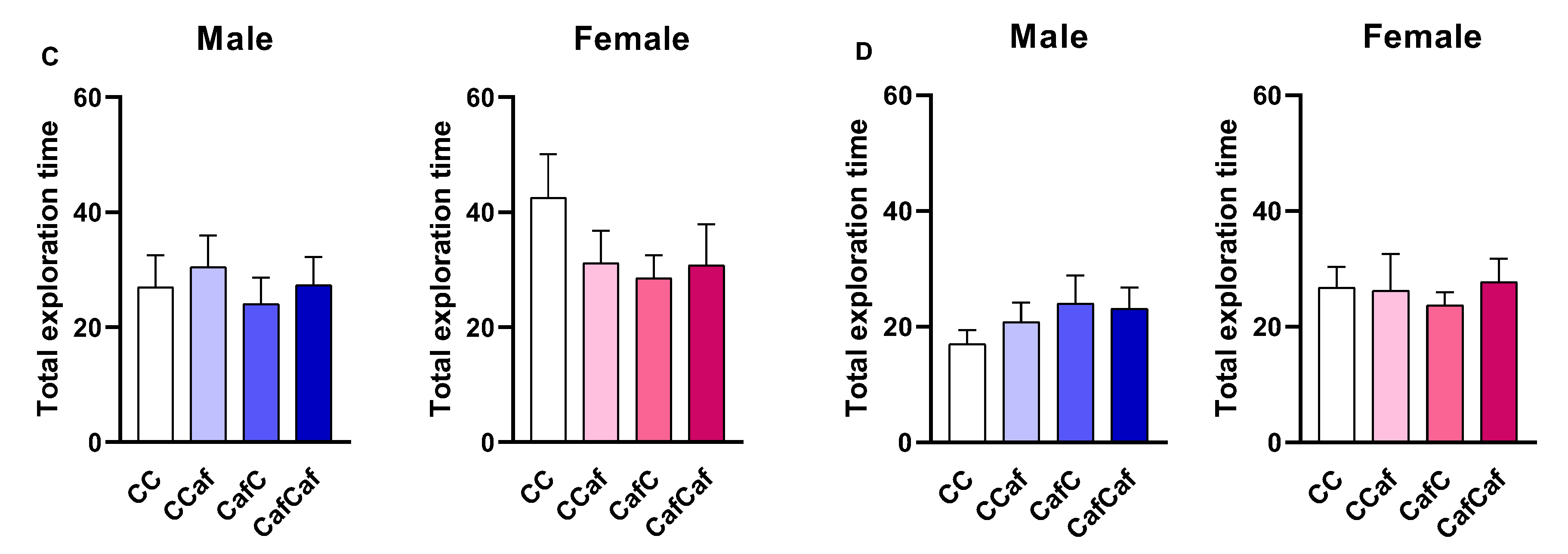
Place/Object Test Behaviour
Endpoint Measures in 14 Week Old Adult Offspring
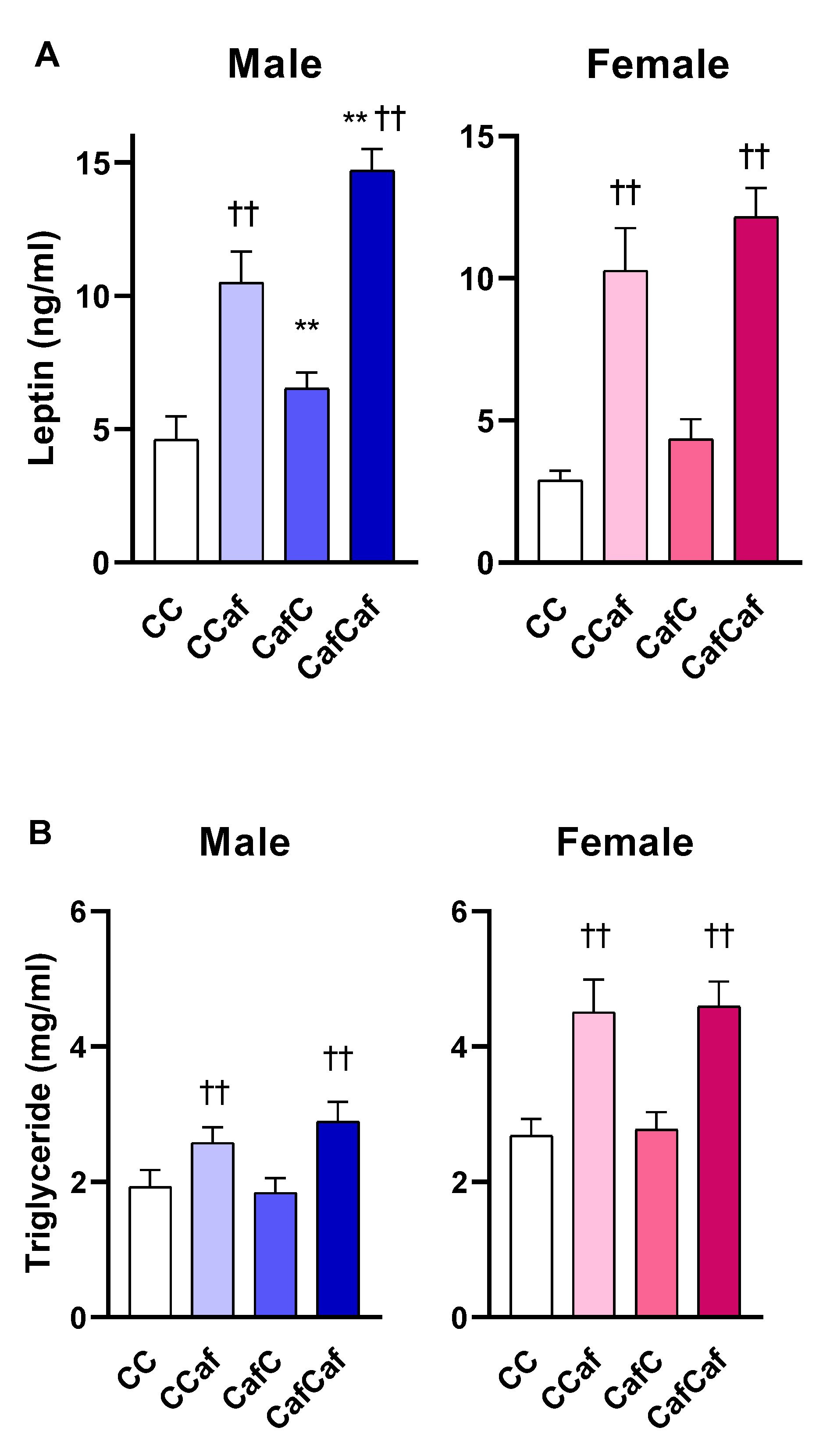
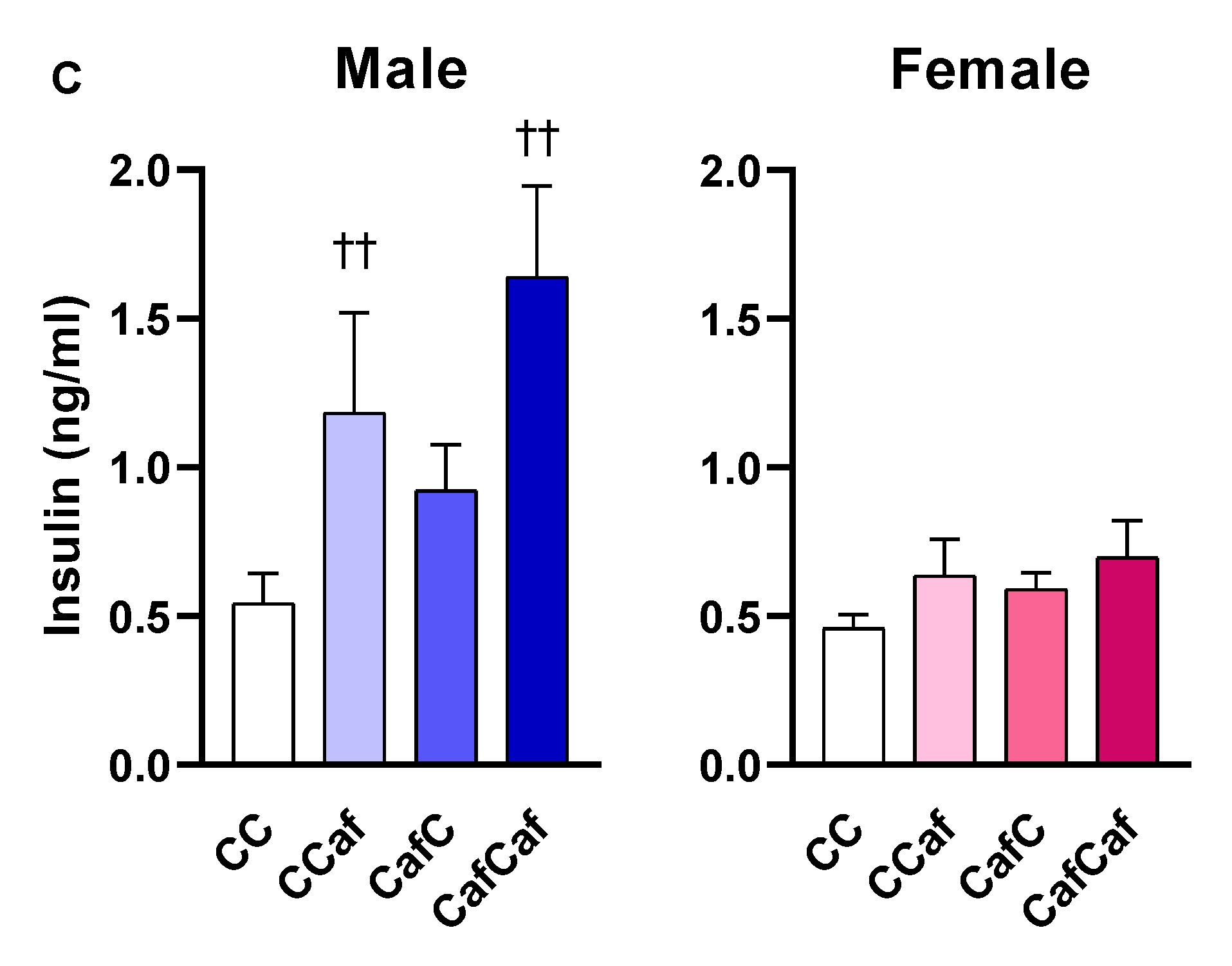
Plasma Measures in 14 Week Old Adult Offspring
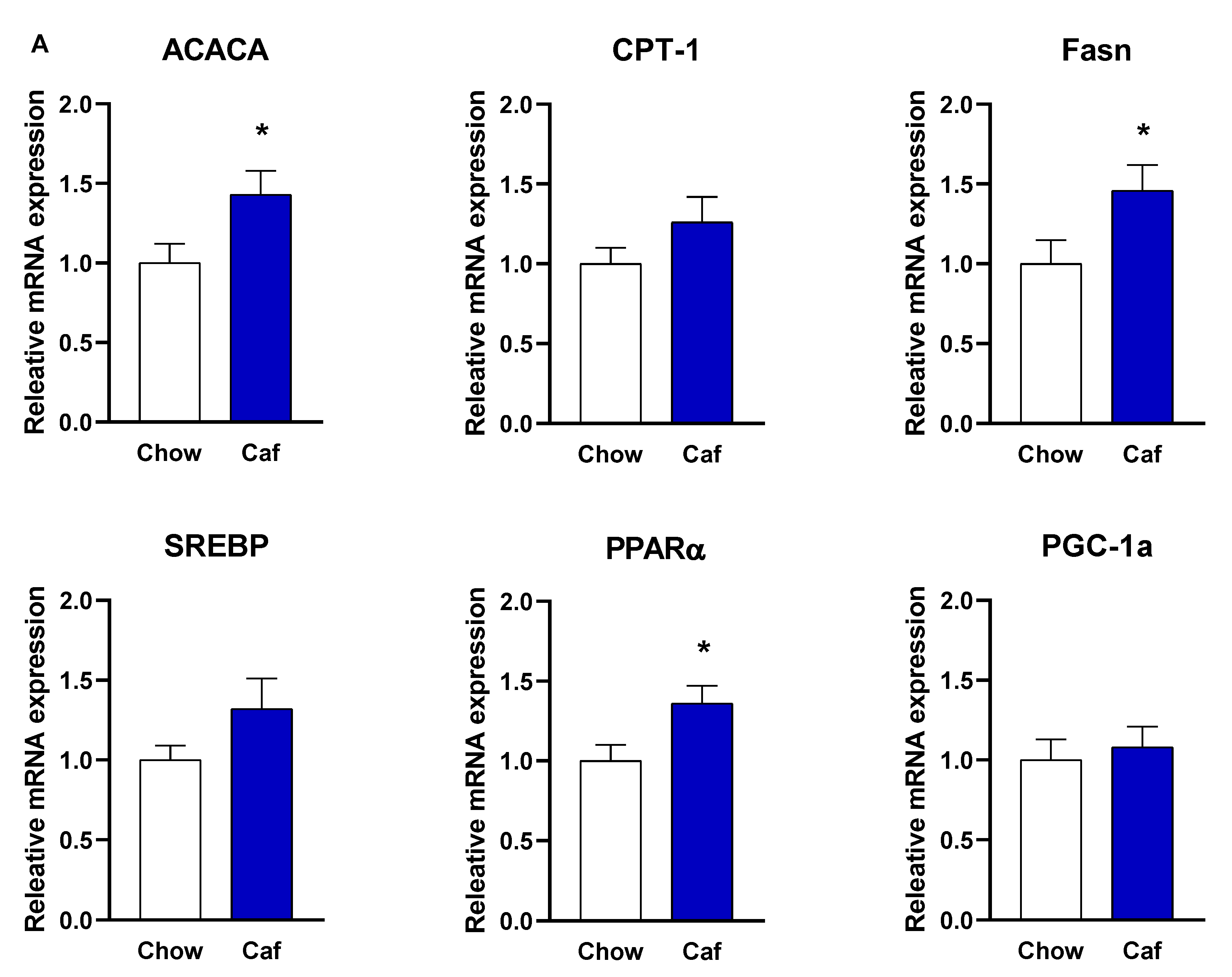
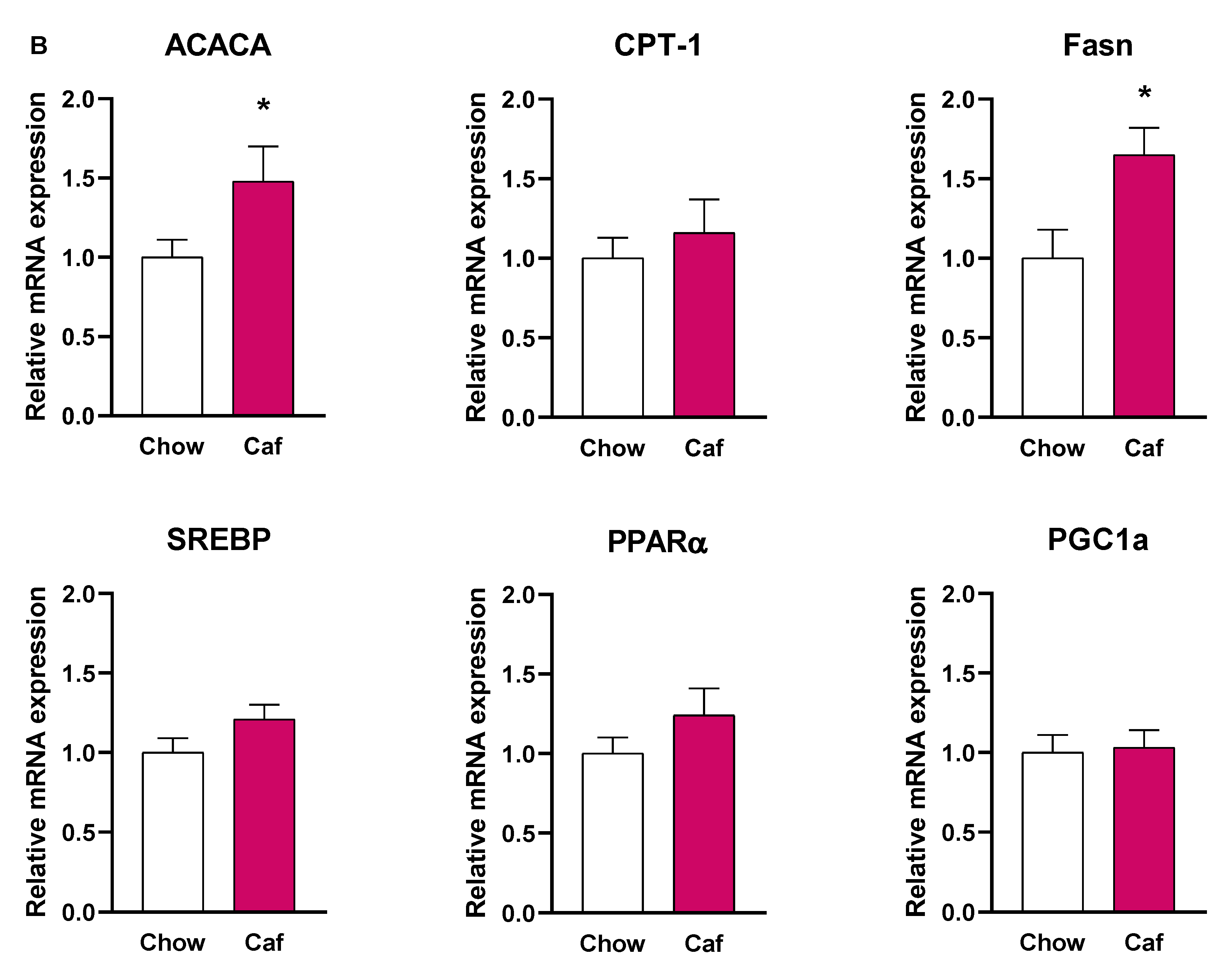
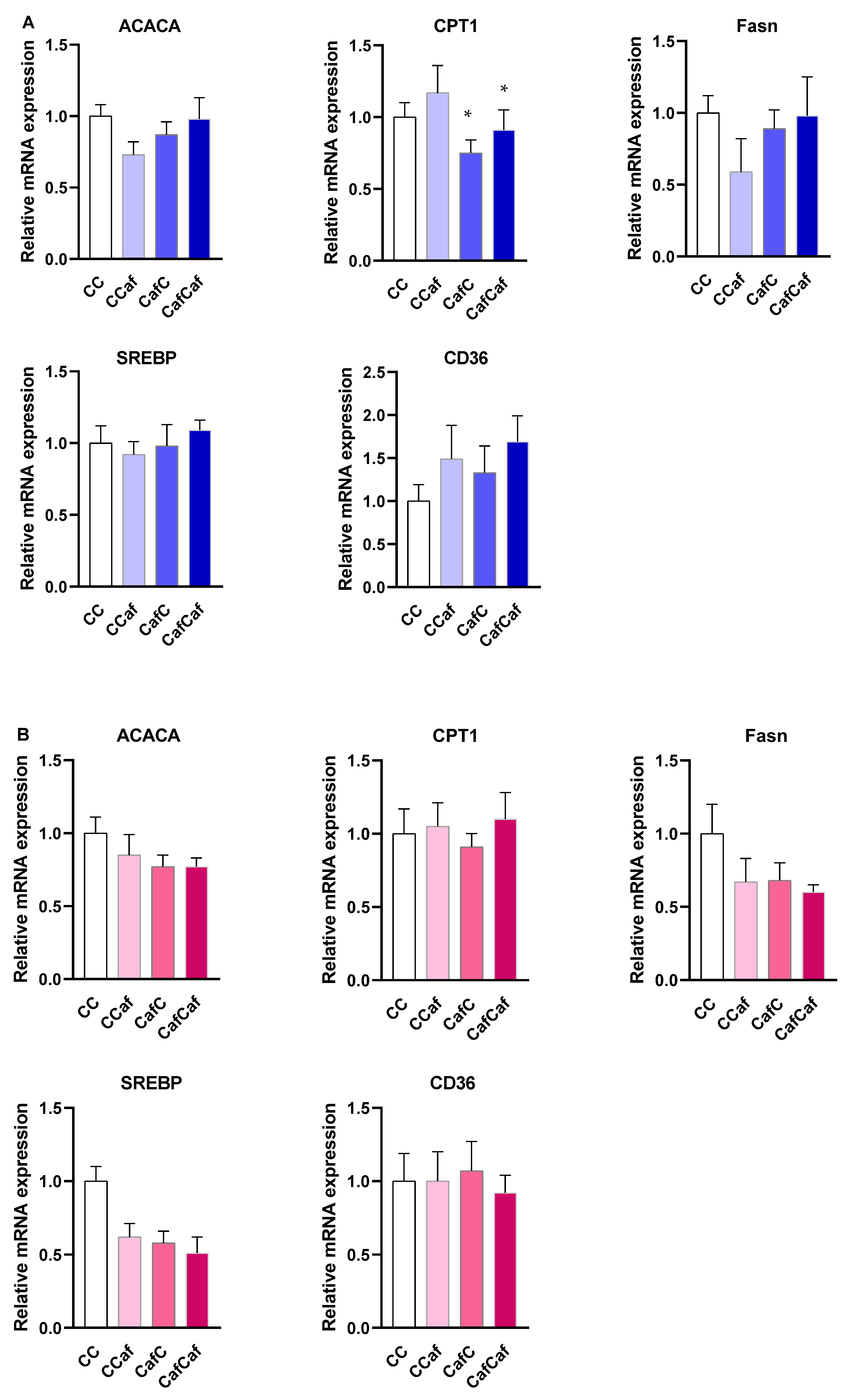
Liver Gene Expression in Weanling (PND 20) and Adult Offspring (14 Weeks)
3. Discussion
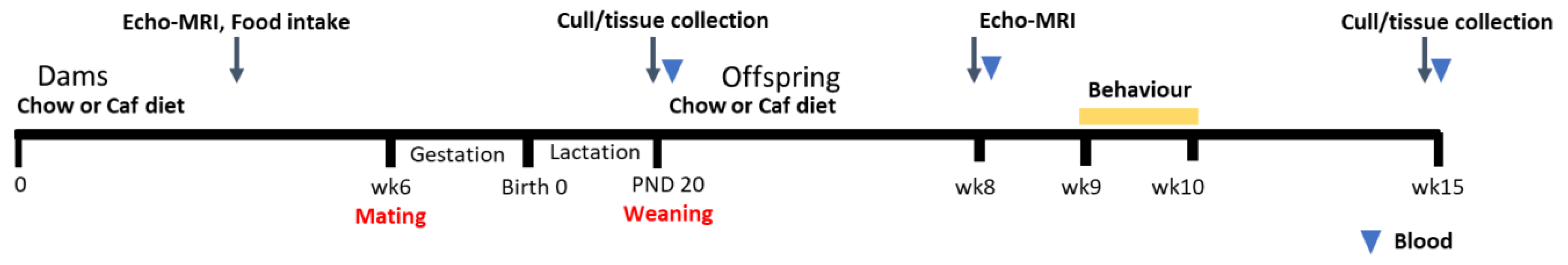
4. Materials and Methods
4.1. Animals and Diet
4.2. Food Intake, Body Composition, and Plasma Glucose Measures in Dams
4.3. Mating and Lactation
4.4. Weaning Tissue Collection and Diet Allocation
4.5. Offspring Metabolic Measures
4.6. Offspring Anxiety-Like Behaviour and Cognitive Measures
4.6.1. Elevated Plus Maze (EPM)
4.6.2. Place and Object Tests
4.7. Adult Plasma and Tissue Collection
4.8. Plasma Hormone Measurements
4.9. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.9.1. RNA Extraction and cDNA Synthesis
4.9.2. Real Time qPCR
4.10. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haththotuwa, R.N.; Wijeyaratne, C.N.; Senarath, U. Worldwide epidemic of obesity. In Obesity and Obstetrics; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–8. [Google Scholar]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity among Adults and Youth: United States, 2015–2016. NCHS Data Brief. No. 288. 2017. Available online: https://stacks.cdc.gov/view/cdc/49223 (accessed on 25 November 2021).
- Huse, O.; Hettiarachchi, J.; Gearon, E.; Nichols, M.; Allender, S.; Peeters, A. Obesity in Australia. Obes. Res. Clin. Pract. 2018, 12, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Vahratian, A. Prevalence of Overweight and Obesity Among Women of Childbearing Age: Results from the 2002 National Survey of Family Growth. Matern. Child Health J. 2009, 13, 268–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiss, K.; Breckenkamp, J.; Borde, T.; Brenne, S.; David, M.; Razum, O. Contribution of overweight and obesity to adverse preg-nancy outcomes among immigrant and non-immigrant women in Berlin, Germany. Eur. J. Public Health 2015, 25, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callaway, L.K.; Chang, A.M.; McIntyre, H.; Prins, J. The prevalence and impact of overweight and obesity in an Australian obstetric population. Med. J. Aust. 2006, 184, 56–59. [Google Scholar] [CrossRef]
- Lindam, A.; Johansson, S.; Stephansson, O.; Wikström, A.-K.; Cnattingius, S. High Maternal Body Mass Index in Early Pregnancy and Risks of Stillbirth and Infant Mortality—A Population-Based Sibling Study in Sweden. Am. J. Epidemiol. 2016, 184, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Guelinckx, I.; Devlieger, R.; Beckers, K.; Vansant, G. Maternal obesity: Pregnancy complications, gestational weight gain and nutrition. Obes. Rev. 2008, 9, 140–150. [Google Scholar] [CrossRef]
- Catalano, P.M.; Ehrenberg, H.M. Review article: The short- and long-term implications of maternal obesity on the mother and her offspring. BJOG Int. J. Obstet. Gynaecol. 2006, 113, 1126–1133. [Google Scholar] [CrossRef]
- Green, L.R.; Hester, R.L. Parental Obesity: Intergenerational Programming and Consequences; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Voerman, E.; Santos, S.; Golab, B.P.; Amiano, P.; Ballester, F.; Barros, H.; Bergström, A.; Charles, M.-A.; Chatzi, L.; Chevrier, C.; et al. Maternal body mass index, gestational weight gain, and the risk of overweight and obesity across childhood: An individual participant data meta-analysis. PLoS Med. 2019, 16, e1002744. [Google Scholar] [CrossRef]
- Schack-Nielsen, L.; Michaelsen, K.F.; Gamborg, M.; Mortensen, E.L.; Sørensen, T.I.A. Gestational weight gain in relation to offspring body mass index and obesity from infancy through adulthood. Int. J. Obes. 2010, 34, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Fraser, A.; Tilling, K.; Macdonald-Wallis, C.; Sattar, N.; Brion, M.-J.; Benfield, L.; Ness, A.; Deanfield, J.; Hingorani, A.; Nelson, S.M.; et al. Association of Maternal Weight Gain in Pregnancy With Offspring Obesity and Metabolic and Vascular Traits in Childhood. Circulation 2010, 121, 2557–2564. [Google Scholar] [CrossRef] [Green Version]
- Dello Russo, M.; Ahrens, W.; De Vriendt, T.; Marild, S.; Molnar, D.; Moreno, L.A.; Reeske, L.A.; Veidebaum, T.; Kourides, Y.A.; Barba, G.; et al. Gestational weight gain and adiposity, fat distribution, metabolic profile, and blood pressure in offspring: The IDEFICS project. Int. J. Obes. 2013, 37, 914–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrey, K.M.; Reynolds, R.M.; Prescott, S.L.; Nyirenda, M.; Jaddoe, V.W.V.; Eriksson, J.G.; Broekman, B.F.P. Influence of maternal obesity on the long-term health of offspring. Lancet Diabetes Endocrinol. 2017, 5, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.M.; Segovia, S.A.; Vickers, M.H. Experimental Models of Maternal Obesity and Neuroendocrine Programming of Metabolic Disorders in Offspring. Front. Endocrinol. 2017, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, E.; Nathanielsz, P.W. Mechanisms by which maternal obesity programs offspring for obesity: Evidence from animal studies. Nutr. Rev. 2013, 71 (Suppl. 1), S42–S54. [Google Scholar] [CrossRef]
- Ribaroff, G.A.; Wastnedge, E.; Drake, A.J.; Sharpe, R.M.; Chambers, T.J.G. Animal models of maternal high fat diet exposure and effects on metabolism in offspring: A meta-regression analysis. Obes. Rev. 2017, 18, 673–686. [Google Scholar] [CrossRef]
- Chen, H.; Morris, M.J. Differential Responses of Orexigenic Neuropeptides to Fasting in Offspring of Obese Mothers. Obesity 2009, 17, 1356–1362. [Google Scholar] [CrossRef] [Green Version]
- Bahari, H.; Caruso, V.; Morris, M.J. Late-Onset Exercise in Female Rat Offspring Ameliorates the Detrimental Metabolic Impact of Maternal Obesity. Endocrinology 2013, 154, 3610–3621. [Google Scholar] [CrossRef] [Green Version]
- Kjaergaard, M.; Nilsson, C.; Rosendal, A.; Nielsen, M.O.; Raun, K. Maternal chocolate and sucrose soft drink intake induces hepatic steatosis in rat offspring associated with altered lipid gene expression profile. Acta Physiol. 2014, 210, 142–153. [Google Scholar] [CrossRef]
- Bayol, S.A.; Simbi, B.H.; Fowkes, R.C.; Stickland, N.C. A maternal “junk food” diet in pregnancy and lactation promotes nonal-coholic fatty liver disease in rat offspring. Endocrinology 2010, 151, 1451–1461. [Google Scholar] [CrossRef] [Green Version]
- Dahlhoff, M.; Pfister, S.; Blutke, A.; Rozman, J.; Klingenspor, M.; Deutsch, M.; Rathkolb, B.; Fink, B.; Gimpfl, M.; de Angelis, M.H.; et al. Peri-conceptional obesogenic exposure induces sex-specific programming of disease susceptibilities in adult mouse offspring. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2014, 1842, 304–317. [Google Scholar] [CrossRef] [Green Version]
- Hasebe, K.; Kendig, M.D.; Morris, M.J. Mechanisms Underlying the Cognitive and Behavioural Effects of Maternal Obesity. Nutrients 2021, 13, 240. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.E.; Barry, C.; Sabhlok, A.; Russell, K.; Majors, A.; Kollins, S.H.; Fuemmeler, B.F. Maternal pre-pregnancy obesity and child neuro-developmental outcomes: A meta-analysis. Obes Rev. 2018, 19, 464–484. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.; Schieve, L.A.; Sharma, A.J.; Hinkle, S.N.; Li, R.; Lind, J.N. Maternal Prepregnancy Body Mass Index and Child Psychosocial Development at 6 Years of Age. Pediatrics 2015, 135, e1198–e1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contu, L.; Hawkes, C.A. A review of the impact of maternal obesity on the cognitive function and mental health of the off-spring. Int. J. Mol. Sci. 2017, 18, 1093. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; de Vega, W.; Sivanathan, S.; St-Cyr, S.; McGowan, P. Maternal high-fat diet alters anxiety behavior and glucocorticoid signaling in adolescent offspring. Neuroscience 2014, 272, 92–101. [Google Scholar] [CrossRef]
- Sasaki, A.; de Vega, W.C.; St-Cyr, S.; Pan, P.; McGowan, P. Perinatal high fat diet alters glucocorticoid signaling and anxiety behavior in adulthood. Neuroscience 2013, 240, 1–12. [Google Scholar] [CrossRef]
- Graf, A.E.; Lallier, S.W.; Waidyaratne, G.; Thompson, M.D.; Tipple, T.E.; Hester, M.E.; Trask, A.J.; Rogers, L.K. Maternal high fat diet exposure is associated with increased hepcidin levels, decreased myelination, and neurobehavioral changes in male offspring. Brain Behav. Immun. 2016, 58, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Zieba, J.; Uddin, G.M.; Youngson, N.A.; Karl, T.; Morris, M.J. Long-term behavioural effects of maternal obesity in C57BL/6J mice. Physiol. Behav. 2019, 199, 306–313. [Google Scholar] [CrossRef]
- O’Reilly, J.R.O.; Reynolds, R.M. The risk of maternal obesity to the long-term health of the offspring. Clin. Endocrinol. 2012, 78, 9–16. [Google Scholar] [CrossRef]
- Andres, A.; Hull, H.R.; Shankar, K.; Casey, P.H.; Cleves, M.A.; Badger, T.M. Longitudinal body composition of children born to mothers with normal weight, overweight, and obesity. Obesity 2015, 23, 1252–1258. [Google Scholar] [CrossRef]
- Rajasingam, D.; Seed, P.T.; Briley, A.L.; Shennan, A.H.; Poston, L. A prospective study of pregnancy outcome and biomarkers of oxidative stress in nulliparous obese women. Am. J. Obstet. Gynecol. 2009, 200, 395.e1–395.e9. [Google Scholar] [CrossRef] [PubMed]
- Pomar, C.A.; Van Nes, R.; Sánchez, J.; Picó, C.; Keijer, J.; Palou, A. Maternal consumption of a cafeteria diet during lactation in rats leads the offspring to a thin-outside-fat-inside phenotype. Int. J. Obes. 2017, 41, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Mucellini, A.B.; Goularte, J.F.; Da Cunha, A.C.D.A.; Caceres, R.C.; Noschang, C.; Da Silva Benetti, C.; Silveira, P.P.; Sanvitto, G.L. Effects of exposure to a cafeteria diet during gestation and after weaning on the metabolism and body weight of adult male offspring in rats. Br. J. Nutr. 2014, 111, 1499–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolls, B.J.; Rowe, E.A. Pregnancy and lactation in the obese rat: Effects on maternal and pup weights. Physiol. Behav. 1982, 28, 393–400. [Google Scholar] [CrossRef]
- Sánchez, J.; Priego, T.; Palou, M.; Tobaruela, A.; Palou, A.; Pico, C. Oral Supplementation with Physiological Doses of Leptin During Lactation in Rats Improves Insulin Sensitivity and Affects Food Preferences Later in Life. Endocrinology 2008, 149, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Ong, Z.Y.; Muhlhausler, B.S. Maternal “junk-food” feeding of rat dams alters food choices and development of the mesolimbic reward pathway in the offspring. FASEB J. 2011, 25, 2167–2179. [Google Scholar] [CrossRef] [Green Version]
- McDonald, S.D.; Han, Z.; Mulla, S.; Beyene, J.; Knowledge Synthesis Group. Overweight and obesity in mothers and risk of preterm birth and low birth weight infants: Systematic review and meta-analyses. BMJ 2010, 341, c3428. [Google Scholar] [CrossRef] [Green Version]
- Drake, A.J.; Reynolds, R.M. Focus on obesity: Impact of maternal obesity on offspring obesity and cardiometabolic disease risk. Reproduction 2010, 140, 387–398. [Google Scholar] [CrossRef]
- Francis, E.C.; Dabelea, D.; Shankar, K.; Perng, W. Maternal diet quality during pregnancy is associated with biomarkers of met-abolic risk among male offspring. Diabetologia 2021, 64, 2478–2490. [Google Scholar] [CrossRef]
- Rajia, S.; Chen, H.; Morris, M.J. Maternal overnutrition impacts offspring adiposity and brain appetite markers-modulation by postweaning diet. J. Neuroendocr. 2010, 22, 905–914. [Google Scholar] [CrossRef] [Green Version]
- Litzenburger, T.; Huber, E.-K.; Dinger, K.; Wilke, R.; Vohlen, C.; Selle, J.; Kadah, M.; Persigehl, T.; Heneweer, C.; Dötsch, J.; et al. Maternal high-fat diet induces long-term obesity with sex-dependent metabolic programming of adipocyte differentiation, hypertrophy and dysfunction in the offspring. Clin. Sci. 2020, 134, 921–939. [Google Scholar] [CrossRef] [PubMed]
- Riant, E.; Waget, A.; Cogo, H.; Arnal, J.-F.; Burcelin, R.; Gourdy, P. Estrogens Protect against High-Fat Diet-Induced Insulin Resistance and Glucose Intolerance in Mice. Endocrinology 2009, 150, 2109–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryzgalova, G.; Gao, H.; Ahren, B.; Zierath, J.R.; Galuska, D.; Steiler, T.L.; Dahlman-Wright, K.; Nilssosn, S.; Gustafsson, J.A.; Efendic, S.; et al. Evidence that oestrogen receptor-α plays an important role in the regulation of glucose homeostasis in mice: Insulin sensitivity in the liver. Diabetologia 2006, 49, 588–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, N.; Ezechukwu, H.C.; Holland, O.J.; Hryciw, D.H. Developmental programming of peripheral diseases in offspring exposed to maternal obesity during pregnancy. Am. J. Physiol. Integr. Comp. Physiol. 2020, 319, R507–R516. [Google Scholar] [CrossRef]
- Dudele, A.; Hougaard, K.S.; Kjolby, M.; Hokland, M.; Winther, G.; Elfving, B.; Wegener, G.; Nielsen, A.L.; Larsen, A.; Nøhr, M.K.; et al. Chronic maternal inflammation or high-fat-feeding programs offspring obesity in a sex-dependent manner. Int. J. Obes. 2017, 41, 1420–1426. [Google Scholar] [CrossRef]
- Kruse, M.; Seki, Y.; Vuguin, P.M.; Du, X.Q.; Fiallo, A.; Glenn, A.S.; Singer, S.; Breuhahn, K.; Katz, E.B.; Charron, M.J. High-Fat Intake During Pregnancy and Lactation Exacerbates High-Fat Diet-Induced Complications in Male Offspring in Mice. Endocrinology 2013, 154, 3565–3576. [Google Scholar] [CrossRef]
- Parente, L.B.; Aguila, M.B.; Mandarim-De-Lacerda, C.A. Deleterious effects of high-fat diet on perinatal and postweaning periods in adult rat offspring. Clin. Nutr. 2008, 27, 623–634. [Google Scholar] [CrossRef]
- Ruager-Martin, R.; Hyde, M.J.; Modi, N. Maternal obesity and infant outcomes. Early Hum. Dev. 2010, 86, 715–722. [Google Scholar] [CrossRef]
- Bringhenti, I.; Ornellas, F.; Martins, M.A.; Mandarim-De-Lacerda, C.; Aguila, M.B. Early hepatic insult in the offspring of obese maternal mice. Nutr. Res. 2015, 35, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Ornellas, F.; Souza-Mello, V.; Mandarim-De-Lacerda, C.A.; Aguila, M.B. Programming of Obesity and Comorbidities in the Progeny: Lessons from a Model of Diet-Induced Obese Parents. PLoS ONE 2015, 10, e0124737. [Google Scholar] [CrossRef]
- Giudetti, A.M.; Micioni Di Bonaventura, M.V.; Ferramosca, A.; Longo, S.; Micioni Di Bonaventura, E.; Friuli, M.; Romano, A.; Gaetani, S.; Cifani, C. Brief daily access to cafeteria-style diet impairs hepatic metabolism even in the absence of excessive body weight gain in rats. FASEB J. 2020, 34, 9358–9371. [Google Scholar] [CrossRef] [PubMed]
- Daniel, Z.C.; Akyol, A.; McMullen, S.; Langley-Evans, S.C. Exposure of neonatal rats to maternal cafeteria feeding during suckling alters hepatic gene expression and DNA methylation in the insulin signalling pathway. Genes Nutr. 2014, 9, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, K.D.; Cagampang, F.R.; Argenton, M.; Zhang, J.; Ethirajan, P.L.; Burdge, G.C.; Bateman, A.C.; Clough, G.F.; Poston, L.; Hanson, M.A.; et al. Maternal high-fat feeding primes steatohepatitis in adult mice offspring, involving mitochondrial dysfunction and altered lipogenesis gene expression. Hepatology 2009, 50, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.; Langley-Evans, S.; Voigt, J.-P. The impact of maternal cafeteria diet on anxiety-related behaviour and exploration in the offspring. Physiol. Behav. 2011, 103, 164–172. [Google Scholar] [CrossRef]
- Speight, A.; Davey, W.G.; McKenna, E.; Voigt, J.-P.W. Exposure to a maternal cafeteria diet changes open-field behaviour in the developing offspring. Int. J. Dev. Neurosci. 2016, 57, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Curi, H.T.; Dias, C.T.; da Luz Camargo, M.L.M.; dos Santos Gomez, P.; Gomes, M.F.P.; Beserra-Filho, J.I.A.; Medeiros, A.; Ribeiro, A.M.; Simabuco, F.M.; Lambertucci, R.H.; et al. Maternal high-fat diet increases anhedonic behavior and modulates hippocampal Mash1 and BDNF expression in adult offspring. Neurosci. Lett. 2021, 764, 136239. [Google Scholar] [CrossRef]
- Clark, T.D.; Crean, A.J.; Senior, A.M. Obesogenic diets induce anxiety in rodents: A systematic review and meta-analysis. Obes. Rev. 2021, e13399. [Google Scholar] [CrossRef]
- Cordner, Z.A.; Khambadkone, S.G.; Boersma, G.J.; Song, L.; Summers, T.N.; Moran, T.H.; Tamashiro, K.L. Maternal high-fat diet results in cog-nitive impairment and hippocampal gene expression changes in rat offspring. Exp. Neurol. 2019, 318, 92–100. [Google Scholar] [CrossRef]
- Leigh, S.-J.; Kendig, M.D.; Morris, M.J. Palatable Western-style Cafeteria Diet as a Reliable Method for Modeling Diet-induced Obesity in Rodenfts. J. Vis. Exp. 2019, 153, e60262. [Google Scholar]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2- ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maternal Diet | |||
|---|---|---|---|
| Chow (11) | Caf (9) | ||
| Litter size (n) | Total | 14.72 ± 1.61 | 14.77 ± 2.90 |
| Female | 7.18 ± 2.27 | 6.55 ± 2.24 | |
| Male | 7.54 ± 1.91 | 8.22 ± 1.71 | |
| Offspring body weight PND 1 (g) | Female | 6.85 ± 0.41 | 6.61 ± 0.32 |
| Male | 7.25 ± 0.44 | 6.81 ± 0.30 ** | |
| Chow (11) | Caf (9) | |
|---|---|---|
| Body weight (g) | 393.4 ± 7.2 | 393.8 ± 8.7 |
| Nasoanal length (cm) | 22.9 ± 0.2 | 23.1 ± 0.2 |
| Girth (cm) | 17.5 ± 0.2 | 17.7 ± 0.3 |
| Liver weight (g) | 18.04 ± 0.58 | 16.92 ± 0.48 |
| RP fat weight (g) | 2.93 ± 0.22 | 4.38 ± 0.56 * |
| Cecum (g) | 7.77 ± 0.41 | 5.15 ± 0.78 * |
| Blood glucose (mmol/L) | 9.69 ± 0.34 | 9.50 ± 0.53 |
| Insulin (ng/mL) | 0.75 ± 0.15 | 0.77 ± 0.10 |
| Leptin (ng/mL) | 1.98 ± 0.36 | 2.71 ± 0.53 |
| Plasma triglyceride (nM) | 2.30 ± 0.29 | 1.46 ± 0.30 |
| Liver triglyceride (mg TG/mg tissue) | 4.71 ± 0.48 | 7.02 ± 1.29 |
| Male | Female | |||
|---|---|---|---|---|
| Maternal Diet | Chow (n = 11) | Caf (n = 10) | Chow (n = 12) | Caf (n = 7) |
| Body weight (g) * | 46.03 ± 2.72 | 42.14 ± 1.22 | 44.31 ± 1.78 | 39.83 ± 1.00 |
| Nasoanal length (cm) * | 10.95 ± 0.24 | 10.55 ± 0.18 | 10.79 ± 0.16 | 10.29 ± 0.08 |
| Girth (cm) | 8.45 ± 0.22 | 7.94 ± 0.25 | 8.54 ± 0.15 | 8.21 ± 0.12 |
| Liver weight (g) * | 1.74 ± 0.12 | 1.51 ± 0.07 | 1.65 ± 0.09 | 1.39 ± 0.06 |
| RP fat weight (g) ** # ^ | 0.07 ± 0.01 | 0.17 ± 0.02 | 0.06 ± 0.01 | 0.09 ± 0.01 |
| Kidney (g) * | 0.26 ± 0.01 | 0.24 ± 0.01 | 0.27 ± 0.01 | 0.23 ± 0.01 |
| Cecum (g) | 0.35 ± 0.05 | 0.30 ± 0.02 | 0.28 ± 0.03 | 0.24 ± 0.02 |
| Blood glucose (mmol/L) * | 9.03 ± 0.37 | 8.38 ± 0.11 | 8.98 ± 0.24 | 8.29 ± 0.12 |
| Male | Female | |||||||
|---|---|---|---|---|---|---|---|---|
| Maternal Diet | Chow | Caf | Chow | Caf | ||||
| Offspring Diet | Chow 11 | Caf 11 | Chow 13 | Caf 13 | Chow 11 | Caf 11 | Chow 11 | Caf 13 |
| Body weight (g) | 581.1 ± 12.8 | 724 ± 24.4 †† | 619 ± 11.3 ** | 805.9 ± 14.4 †† ** | 309.4 ± 9.6 | 444.9 ± 27.8 †† | 312.9 ± 10 | 433.7 ± 16.6 †† |
| Length (cm) | 25.9 ± 0.2 | 26.9 ± 0.2 †† | 26.2 ± 0.1 | 27.3 ± 0.2 †† | 22.3 ± 0.2 | 23.2 ± 0.3 †† | 22.2 ± 0.2 | 23 ± 0.16 †† |
| Tibia (cm) | 4.31 ± 0.04 | 4.32 ± 0.04 | 4.33 ± 0.03 | 4.31 ± 0.04 | 3.77 ± 0.04 | 3.75 ± 0.04 | 3.72 ± 0.04 | 3.72 ± 0.04 |
| Girth (cm) | 20.4 ± 0.3 | 23.5 ± 0.5 †† | 21.6 ± 0.3 * | 24.5 ± 0.3 †† * | 15.4 ± 0.3 | 19.3 ± 0.8 †† | 16.1 ± 0.2 | 19.7 ± 0.5 †† |
| Liver (g) | 20.74 ± 0.8 | 25.39 ± 1.4 †† | 23.25 ± 0.5 * | 28.68 ± 1.0 †† * | 11.17 ± 0.4 | 13.56 ± 0.8 †† | 10.36 ± 0.3 | 12.68 ± 0.4 †† |
| Left ventricle (g) | 1.14 ± 0.06 | 1.33 ± 0.08 †† | 1.11 ± 0.03 | 1.27 ± 0.03 †† | 0.67 ± 0.03 | 0.88 ± 0.06 †† | 0.67 ± 0.02 | 0.81 ± 0.03 †† |
| RP fat (g) | 4.31 ± 0.4 | 12.96 ± 0.84 †† | 5.52 ± 0.55 * | 15.72 ± 0.80 †† * | 2.64 ± 0.58 | 7.2 ± 0.77 †† | 2.62 ± 0.37 | 6.14 ± 0.37 †† |
| Glucose (mmol/L) | 11.7 ± 0.5 | 11.3 ± 0.7 | 11.5 ± 0.3 | 10.9 ± 0.3 | 9.6 ± 0.3 | 11.4 ± 0.3 †† | 10.4 ± 0.4 | 11.8 ± 0.5 †† |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tajaddini, A.; Kendig, M.D.; Prates, K.V.; Westbrook, R.F.; Morris, M.J. Male Rat Offspring Are More Impacted by Maternal Obesity Induced by Cafeteria Diet than Females—Additive Effect of Postweaning Diet. Int. J. Mol. Sci. 2022, 23, 1442. https://doi.org/10.3390/ijms23031442
Tajaddini A, Kendig MD, Prates KV, Westbrook RF, Morris MJ. Male Rat Offspring Are More Impacted by Maternal Obesity Induced by Cafeteria Diet than Females—Additive Effect of Postweaning Diet. International Journal of Molecular Sciences. 2022; 23(3):1442. https://doi.org/10.3390/ijms23031442
Chicago/Turabian StyleTajaddini, Aynaz, Michael D. Kendig, Kelly V. Prates, R. Frederick Westbrook, and Margaret J. Morris. 2022. "Male Rat Offspring Are More Impacted by Maternal Obesity Induced by Cafeteria Diet than Females—Additive Effect of Postweaning Diet" International Journal of Molecular Sciences 23, no. 3: 1442. https://doi.org/10.3390/ijms23031442
APA StyleTajaddini, A., Kendig, M. D., Prates, K. V., Westbrook, R. F., & Morris, M. J. (2022). Male Rat Offspring Are More Impacted by Maternal Obesity Induced by Cafeteria Diet than Females—Additive Effect of Postweaning Diet. International Journal of Molecular Sciences, 23(3), 1442. https://doi.org/10.3390/ijms23031442