
Genome-Wide Identification and Characterization of SET Domain Family Genes in Brassica napus L.


Abstract
:1. Introduction
2. Results
2.1. Identification of SDGs in B. napus
2.2. Phylogenetic Analysis of BnSDGs
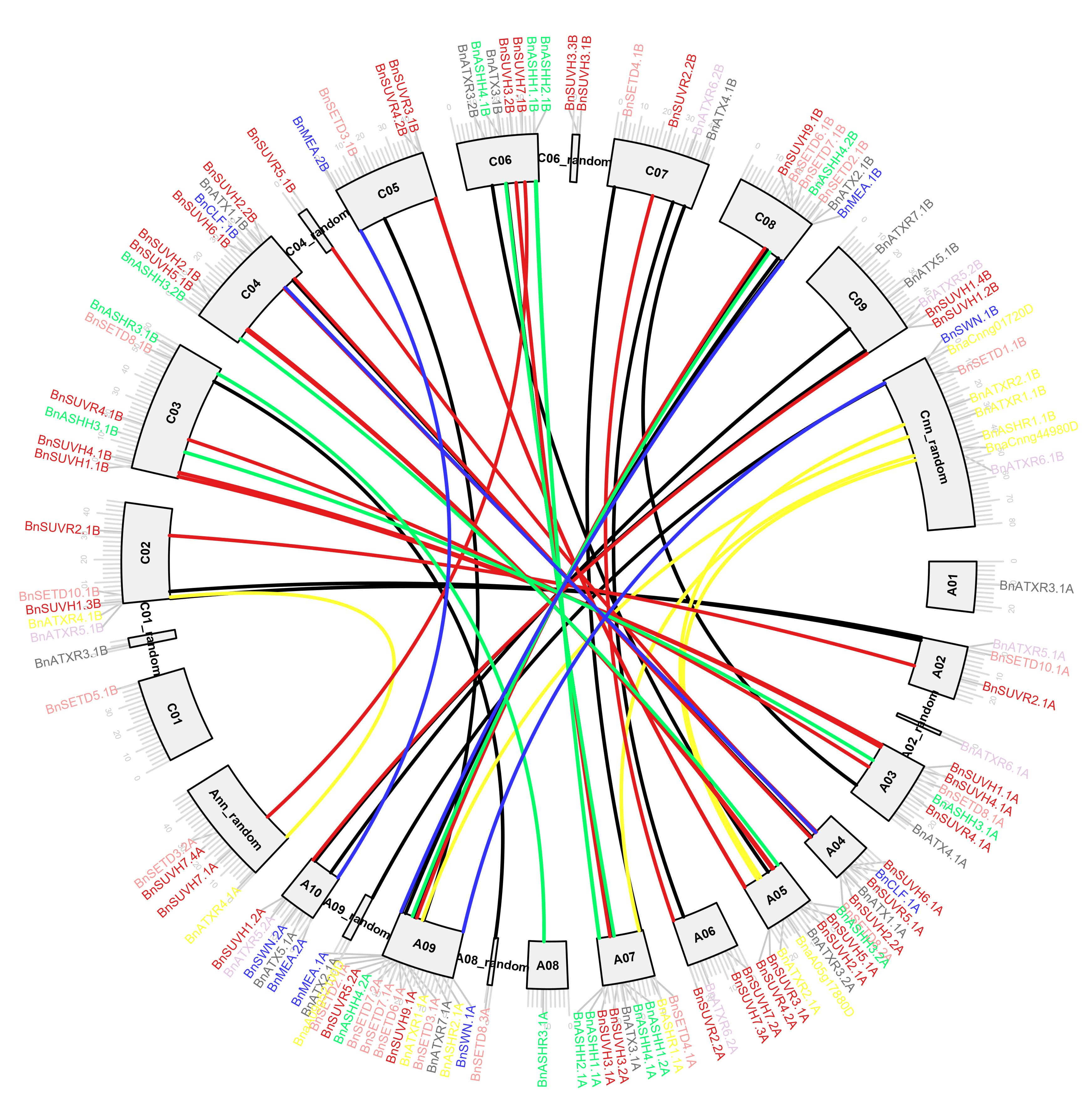
2.3. Assignment of BnSDGs to Chromosomes and Synteny Analysis
2.4. Structural (Domain and Motif Conservation) and Functional Analysis (Gene Ontology (GO)) of BnSDGs Proteins
2.5. Gene Structure and Promoter Analysis of BnSDGs
2.6. In Silico Differential Expression Analysis
2.7. In Silico Abiotic Stress Expression Analysis
2.8. The Expression Validation by Reverse Transcription-Quantitative PCR
3. Discussions
4. Materials and Methods
4.1. Identification of SET Domain-Containing Genes (SDGs) in Brassica napus (B. napus)
4.2. Phylogenetic Analysis
4.3. Chromosomal Location, Synteny Analysis, and Ka/Ks Ratio
4.4. The Biophysical, Structural and Functional Analysis of BnSDG Proteins
4.5. The Identification of Cis-Regulatory Elements and Transcription Binding Sites in Promoter Regions
4.6. In Silico Expression Analysis of BnSDGs
4.7. Plant Material, RNA Extraction, and Reverse Transcription-Quantitative PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kornberg, R.D. Chromatin structure: A repeating unit of histones and DNA. Science 1974, 184, 868–871. [Google Scholar] [CrossRef] [PubMed]
- Jenuwein, T.; Allis, C.D. Translating the histone code. Science 2001, 293, 1074–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef]
- Feng, Q.; Wang, H.; Ng, H.H.; Erdjument-bromage, H.; Tempst, P.; Struhl, K.; Zhang, Y.; Hill, C.; Carolina, N. Methylation of H3-Lysine 79 Is Mediated by a New Family of HMTases without a SET Domain University of North Carolina at Chapel Hill. Current 2002, 12, 1052–1058. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Lu, F.; Cui, X.; Cao, X. Histone methylation in higher plants. Annu. Rev. Plant Biol. 2010, 61, 395–420. [Google Scholar] [CrossRef]
- Trievel, R.C.; Flynn, E.M.; Houtz, R.L.; Hurley, J.H. Mechanism of multiple lysine methylation by the SET domain enzyme Rubisco LSMT. Nat. Struct. Biol. 2003, 10, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Marmorstein, R. Structure of SET domain proteins: A new twist on histone methylation. Trends Biochem. Sci. 2003, 28, 59–62. [Google Scholar] [CrossRef]
- Rea, S.; Eisenhaber, F.; O’Carroll, D.; Strahl, B.D.; Sun, Z.W.; Schmid, M.; Opravil, S.; Mechtier, K.; Ponting, C.P.; Allis, C.D.; et al. Regulation of chromatin structure by site-specific histone H3 methyltransferases. Nature 2000, 406, 593–599. [Google Scholar] [CrossRef]
- Baumbusch, L.O.; Thorstensen, T.; Krauss, V.; Fischer, A.; Naumann, K.; Assalkhou, R.; Schulz, I.; Reuter, G.; Aalen, R.B. The Arabidopsis thaliana genome contains at least 29 active genes encoding SET domain proteins that can be assigned to four evolutionarily conserved classes. Nucleic. Acids. Res. 2001, 29, 4319–4333. [Google Scholar] [CrossRef]
- Ng, D.W.K.; Wang, T.; Chandrasekharan, M.B.; Aramayo, R.; Kertbundit, S.; Hall, T.C. Plant SET domain-containing proteins: Structure, function and regulation. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 2007, 1769, 316–329. [Google Scholar] [CrossRef] [Green Version]
- Pontvianne, F.; Blevins, T.; Pikaard, C.S. Arabidopsis histone lysine methyltransferases. Adv. Bot. Res. 2010, 53, 1–22. [Google Scholar]
- Dong, H.; Liu, D.; Han, T.; Zhao, Y.; Sun, J.; Lin, S.; Cao, J.; Chen, Z.H.; Huang, L. Diversification and evolution of the SDG gene family in Brassica rapa after the whole genome triplication. Sci. Rep. 2015, 5, 16851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, L.; Zhou, S.-L.; Ma, H.; Zhang, L.-S. Expansion and diversification of the SET domain gene family following whole-genome duplications in Populus trichocarpa. BMC Evol. Biol. 2012, 12, 51. [Google Scholar] [CrossRef] [Green Version]
- Aquea, F.; Vega, A.; Timmermann, T.; Poupin, M.J.; Arce-Johnson, P. Genome-wide analysis of the SET DOMAIN GROUP family in grapevine. Plant Cell Rep. 2011, 30, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Xi, Y.; Cheng, B.; Zhu, S.; Kan, X. Identification and characterization of the SET domain gene family in maize. Mol. Biol. Rep. 2014, 41, 1341–1354. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Huang, X.; Ouyang, Y.; Yao, J. Genome-Wide Identification, Phylogenetic and Co-Expression Analysis of OsSET Gene Family in Rice. PLoS ONE 2013, 8, E0065426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cigliano, R.A.; Sanseverino, W.; Cremona, G.; Ercolano, M.R.; Conicella, C.; Consiglio, F.M. Genome-wide analysis of histone modifiers in tomato: Gaining an insight into their developmental roles. BMC Genomics 2013, 14, 57. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Xu, H.; Liu, Y.; Wang, X.; Xu, Q.; Deng, X. Genome-wide identification of sweet orange (Citrus sinensis) histone modification gene families and their expression analysis during the fruit development and fruit-blue mold infection process. Front. Plant Sci. 2015, 6, 607. [Google Scholar] [CrossRef] [Green Version]
- Peng, M.; Ying, P.; Liu, X.; Li, C.; Xia, R.; Li, J.; Zhao, M. Genome-wide identification of histone modifiers and their expression patterns during fruit abscission in litchi. Front. Plant Sci. 2017, 8, 639. [Google Scholar] [CrossRef]
- Huang, Y.; Mo, Y.; Chen, P.; Yuan, X.; Meng, F.; Zhu, S.; Liu, Z. Identification of SET Domain-Containing Proteins in Gossypium raimondii and Their Response to High Temperature Stress. Sci. Rep. 2016, 6, 32729. [Google Scholar] [CrossRef] [Green Version]
- Yadav, C.B.; Muthamilarasan, M.; Dangi, A.; Shweta, S.; Prasad, M. Comprehensive analysis of SET domain gene family in foxtail millet identifies the putative role of SiSET14 in abiotic stress tolerance. Sci. Rep. 2016, 6, 32621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batra, R.; Gautam, T.; Pal, S.; Chaturvedi, D.; Rakhi; Jan, I.; Balyan, H.S.; Gupta, P.K. Identification and characterization of SET domain family genes in bread wheat (Triticum aestivum L.). Sci. Rep. 2020, 10, 14624. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-H.; Qiu, H.-L.; Huang, Y.; Zhang, L.; Si, J.-P. Genome-wide identification and expression profiling of SET DOMAIN GROUP family in Dendrobium catenatum. BMC Plant Biol. 2020, 20, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazzonelli, C.I.; Cuttriss, A.J.; Cossetto, S.B.; Pye, W.; Crisp, P.; Whelan, J.; Finnegan, E.J.; Turnbull, C.; Pogson, B.J. Regulation of carotenoid composition and shoot branching in Arabidopsis by a chromatin modifying histone methyltransferase, SDG8. Plant Cell 2009, 21, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Grini, P.E.; Thorstensen, T.; Alm, V.; Vizcay-Barrena, G.; Windju, S.S.; Jørstad, T.S.; Wilson, Z.A.; Aalen, R.B. The ASH1 HOMOLOG 2 (ASHH2) histone H3 methyltransferase is required for ovule and anther development in Arabidopsis. PLoS ONE 2009, 4, e7817. [Google Scholar] [CrossRef] [Green Version]
- Cartagena, J.A.; Matsunaga, S.; Seki, M.; Kurihara, D.; Yokoyama, M.; Shinozaki, K.; Fujimoto, S.; Azumi, Y.; Uchiyama, S.; Fukui, K. The Arabidopsis SDG4 contributes to the regulation of pollen tube growth by methylation of histone H3 lysines 4 and 36 in mature pollen. Dev. Biol. 2008, 315, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Berr, A.; McCallum, E.J.; Ménard, R.; Meyer, D.; Fuchs, J.; Dong, A.; Shen, W.-H. Arabidopsis SET DOMAIN GROUP2 is required for H3K4 trimethylation and is crucial for both sporophyte and gametophyte development. Plant Cell 2010, 22, 3232–3248. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Shen, W. Crucial function of histone lysine methylation in plant reproduction. Chin. Sci. Bull. 2011, 56, 3493–3499. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Tyson, M.D.; Jackson, S.S.; Yadegari, R. Partially redundant functions of two SET-domain polycomb-group proteins in controlling initiation of seed development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 13244–13249. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Li, D.; Jin, L.; Ruan, Y.; Shen, W.H.; Liu, C. Histone lysine methyltransferases BnaSDG8.A and BnaSDG8.C are involved in the floral transition in Brassica napus. Plant J. 2018, 95, 672–685. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, Y.; Liang, Y.; Zhou, D.; Li, S.; Lin, S.; Dong, H.; Huang, L. The function of histone lysine methylation related SET domain group proteins in plants. Protein Sci. 2020, 29, 1120–1137. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Xu, Y.; Yang, C.; Ouellette, L.; Niu, L.; Zhou, X.; Chu, L.; Zhuang, F.; Liu, J.; Wu, H. Histone tales: Lysine methylation, a protagonist in Arabidopsis development. J. Exp. Bot. 2020, 71, 793–807. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Wöhrmann, H.J.P.; Raissig, M.T.; Arand, J.; Gheyselinck, J.; Gagliardini, V.; Heichinger, C.; Walter, J.; Grossniklaus, U. The P olycomb group protein MEDEA and the DNA methyltransferase MET 1 interact to repress autonomous endosperm development in A rabidopsis. Plant J. 2013, 73, 776–787. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Ma, D.-P.; Li, J. The histone methyltransferase SDG8 regulates shoot branching in Arabidopsis. Biochem. Biophys. Res. Commun. 2008, 373, 659–664. [Google Scholar] [CrossRef]
- Napsucialy-Mendivil, S.; Alvarez-Venegas, R.; Shishkova, S.; Dubrovsky, J.G. Arabidopsis homolog of trithorax1 (ATX1) is required for cell production, patterning, and morphogenesis in root development. J. Exp. Bot. 2014, 65, 6373–6384. [Google Scholar] [CrossRef] [Green Version]
- Chanvivattana, Y.; Bishopp, A.; Schubert, D.; Stock, C.; Moon, Y.-H.; Sung, Z.R.; Goodrich, J. Interaction of Polycomb-group proteins controlling flowering in Arabidopsis. Plant Cell 2004, 1, 117–133. [Google Scholar]
- Berr, A.; McCallum, E.J.; Alioua, A.; Heintz, D.; Heitz, T.; Shen, W.-H. Arabidopsis histone methyltransferase SET DOMAIN GROUP8 mediates induction of the jasmonate/ethylene pathway genes in plant defense response to necrotrophic fungi. Plant Physiol. 2010, 154, 1403–1414. [Google Scholar] [CrossRef] [Green Version]
- Raynaud, C.; Sozzani, R.; Glab, N.; Domenichini, S.; Perennes, C.; Cella, R.; Kondorosi, E.; Bergounioux, C. Two cell-cycle regulated SET-domain proteins interact with proliferating cell nuclear antigen (PCNA) in Arabidopsis. Plant J. 2006, 47, 395–407. [Google Scholar] [CrossRef]
- Pien, S.; Fleury, D.; Mylne, J.S.; Crevillen, P.; Inzé, D.; Avramova, Z.; Dean, C.; Grossniklaus, U. ARABIDOPSIS TRITHORAX1 dynamically regulates FLOWERING LOCUS C activation via histone 3 lysine 4 trimethylation. Plant Cell 2008, 20, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Saleh, A.; Alvarez-Venegas, R.; Yilmaz, M.; Le, O.; Hou, G.; Sadder, M.; Al-Abdallat, A.; Xia, Y.; Lu, G.; Ladunga, I. The highly similar Arabidopsis homologs of trithorax ATX1 and ATX2 encode proteins with divergent biochemical functions. Plant Cell 2008, 20, 568–579. [Google Scholar] [CrossRef] [Green Version]
- Tamada, Y.; Yun, J.-Y.; Woo, S.C.; Amasino, R.M. ARABIDOPSIS TRITHORAX-RELATED7 is required for methylation of lysine 4 of histone H3 and for transcriptional activation of FLOWERING LOCUS C. Plant Cell 2009, 21, 3257–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krichevsky, A.; Gutgarts, H.; Kozlovsky, S.V.; Tzfira, T.; Sutton, A.; Sternglanz, R.; Mandel, G.; Citovsky, V. C2H2 zinc finger-SET histone methyltransferase is a plant-specific chromatin modifier. Dev. Biol. 2007, 303, 259–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Yu, Y.; Meyer, D.; Wu, C.; Shen, W.-H. Prevention of early flowering by expression of FLOWERING LOCUS C requires methylation of histone H3 K36. Nat. Cell Biol. 2005, 7, 1256–1260. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zhang, J.; Li, T.; Fu, X.; Satheesh, V.; Niu, Q.; Lang, Z.; Zhu, J.; Lei, M. A group of SUVH methyl-DNA binding proteins regulate expression of the DNA demethylase ROS1 in Arabidopsis. J. Integr. Plant Biol. 2019, 61, 110–119. [Google Scholar] [CrossRef]
- Serre, N.B.C.; Alban, C.; Bourguignon, J.; Ravanel, S. An outlook on lysine methylation of non-histone proteins in plants. J. Exp. Bot. 2018, 69, 4569–4581. [Google Scholar] [CrossRef]
- Cazzonelli, C.I.; Nisar, N.; Roberts, A.C.; Murray, K.D.; Borevitz, J.O.; Pogson, B.J. A chromatin modifying enzyme, SDG8, is involved in morphological, gene expression, and epigenetic responses to mechanical stimulation. Front. Plant Sci. 2014, 5, 533. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Avramova, Z.; Fromm, M. The Arabidopsis trithorax-like factor ATX1 functions in dehydration stress responses via ABA-dependent and ABA-independent pathways. Plant J. 2011, 66, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Fromm, M.; Avramova, Z. Multiple exposures to drought’train’transcriptional responses in Arabidopsis. Nat. Commun. 2012, 3, 740. [Google Scholar] [CrossRef]
- Liu, N.; Fromm, M.; Avramova, Z. H3K27me3 and H3K4me3 chromatin environment at super-induced dehydration stress memory genes of Arabidopsis thaliana. Mol. Plant 2014, 7, 502–513. [Google Scholar] [CrossRef] [Green Version]
- Palma, K.; Thorgrimsen, S.; Malinovsky, F.G.; Fiil, B.K.; Nielsen, H.B.; Brodersen, P.; Hofius, D.; Petersen, M.; Mundy, J. Autoimmunity in Arabidopsis acd11 is mediated by epigenetic regulation of an immune receptor. PLoS Pathog. 2010, 6, e1001137. [Google Scholar] [CrossRef]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Flagel, L.E.; Wendel, J.F. Gene duplication and evolutionary novelty in plants. New Phytol. 2009, 183, 557–564. [Google Scholar] [CrossRef]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. TIG 2002, 18, 486. [Google Scholar] [CrossRef]
- Song, X.; Wei, Y.; Xiao, D.; Gong, K.; Sun, P.; Ren, Y.; Yuan, J.; Wu, T.; Yang, Q.; Li, X. Brassica carinata genome characterization clarifies U’s triangle model of evolution and polyploidy in Brassica. Plant Physiol. 2021, 186, 388–406. [Google Scholar] [CrossRef]
- Springer, N.M.; Napoli, C.A.; Selinger, D.A.; Pandey, R.; Cone, K.C.; Chandler, V.L.; Kaeppler, H.F.; Kaeppler, S.M. Comparative analysis of SET domain proteins in maize and Arabidopsis reveals multiple duplications preceding the divergence of monocots and dicots. Plant Physiol. 2003, 132, 907–925. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Venegas, R.; Pien, S.; Sadder, M.; Witmer, X.; Grossniklaus, U.; Avramova, Z. ATX-1, an Arabidopsis homolog of trithorax, activates flower homeotic genes. Curr. Biol. 2003, 13, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Yu, Y.; Law, J.A.; Zhang, X. SET DOMAIN GROUP2 is the major histone H3 lysine 4 trimethyltransferase in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 18557–18562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staiger, D.; Apel, K. Circadian clock-regulated expression of an RNA-binding protein in Arabidopsis: Characterisation of a minimal promoter element. Mol. Gen. Genet. 1999, 261, 811–819. [Google Scholar] [CrossRef]
- Menkens, A.E.; Schindler, U.; Cashmore, A.R. The G-box: A ubiquitous regulatory DNA element in plants bound by the GBF family of bZIP proteins. Trends Biochem. Sci. 1995, 20, 506–510. [Google Scholar] [CrossRef]
- Guiltinan, M.J. Erratum: A plant leucine zipper protein that recognizes an abscisic acid element (Science (267–269)). Science 1990, 250, 612. [Google Scholar] [CrossRef]
- Ding, Y.; Zhu, J.; Zhao, D.; Liu, Q.; Yang, Q.; Zhang, T. Targeting cis-regulatory elements for rice grain quality improvement. Front. Plant Sci. 2021, 12, 705834. [Google Scholar] [CrossRef]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface 289660. bioRxiv 2018, 13, 1194–1202. [Google Scholar]
- Wei, L.; Zhu, Y.; Liu, R.; Zhang, A.; Zhu, M.; Xu, W.; Lin, A.; Lu, K.; Li, J. Genome wide identification and comparative analysis of glutathione transferases (GST) family genes in Brassica napus. Sci. Rep. 2019, 9, 9196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Chen, Y.; Lu, C.; Hwang, J. Prediction of protein subcellular localization. Proteins Struct. Funct. Bioinforma. 2006, 64, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ali, U.; Zhang, G.; Yu, L.; Fang, S.; Iqbal, S.; Li, H.; Lu, S.; Guo, L. Transcriptome analysis reveals genes commonly responding to multiple abiotic stresses in rapeseed. Mol. Breed. 2019, 39, 158. [Google Scholar] [CrossRef]
- Raza, A.; Su, W.; Gao, A.; Mehmood, S.S.; Hussain, M.A.; Nie, W.; Lv, Y.; Zou, X.; Zhang, X. Catalase (CAT) Gene Family in Rapeseed (Brassica napus L.): Genome-Wide Analysis, Identification, and Expression Pattern in Response to Multiple Hormones and Abiotic Stress Conditions. Int. J. Mol. Sci. 2021, 22, 4281. [Google Scholar] [CrossRef] [PubMed]
- Swift, M.L. GraphPad prism, data analysis, and scientific graphing. J. Chem. Inf. Comput. Sci. 1997, 37, 411–412. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript ID | Gene Name | Class | Chromosome: Location Start: Location End: Strand | Gene Size (bp) | CDS (bp) | Protein Length (aa) | PI | GRAVY | No. of Exons-Introns | Cellular Localization | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | BnaA09g51240D | BnMEA.1A | A09:33715032:33719248:− | 4217 | 1932 | 643 | 6.3 | −0.77 | 17–16 | Nuclear | |
| 2 | BnaC08g46050D | BnMEA.1B | I | C08:38367870:38372089:− | 4220 | 1995 | 664 | 6.71 | −0.757 | 17–16 | Nuclear |
| 3 | BnaA10g01220D | BnMEA.2A | A10:640151:647345:+ | 7195 | 1917 | 638 | 8.65 | −0.71 | 18–17 | Nuclear | |
| 4 | BnaC05g01310D | BnMEA.2B | C05:686198:689713:+ | 3516 | 1332 | 443 | 8.61 | −0.671 | 15–14 | Nuclear | |
| 5 | BnaA04g13630D | BnCLF.1A | A04:11524589:11529333:+ | 4745 | 2724 | 907 | 9.05 | −0.876 | 17–16 | Nuclear | |
| 6 | BnaC04g35880D | BnCLF.1B | C04:37428287:37432998:+ | 4712 | 2733 | 910 | 9.07 | −0.851 | 17–16 | Nuclear | |
| 7 | BnaA09g00500D | BnSWN.1A | A09:243397:248630:+ | 5234 | 2559 | 852 | 5.76 | −0.757 | 17–16 | Nuclear | |
| 8 | BnaCnng01170D | BnSWN.1B | Cnn_random:1341220:1346669:− | 5450 | 2571 | 856 | 5.78 | −0.797 | 18–17 | Nuclear | |
| 9 | BnaA10g10150D | BnSWN.2A | A10:8730712:8735228:+ | 4517 | 2526 | 841 | 5.61 | −0.707 | 16–15 | Nuclear | |
| 10 | BnaA03g20680D | BnASHH3.1A | II | A03:9804836:9807462:+ | 2627 | 1110 | 369 | 5.35 | −0.507 | 12–11 | Nuclear |
| 11 | BnaC03g24710D | BnASHH3.1B | C03:13880644:13883273:+ | 2630 | 1098 | 365 | 5.61 | −0.549 | 12–11 | Nuclear | |
| 12 | BnaA05g03780D | BnASHH3.2A | A05:2018708:2020905:+ | 2198 | 1098 | 365 | 6.37 | −0.519 | 11–10 | Nuclear | |
| 13 | BnaC04g03390D | BnASHH3.2B | C04:2408832:2411322:+ | 2491 | 1134 | 377 | 6.87 | −0.516 | 12–11 | Nuclear | |
| 14 | BnaA07g18580D | BnASHH4.1A | A07:15109224:15112095:+ | 2872 | 966 | 321 | 9.32 | −0.679 | 9–8 | Nuclear | |
| 15 | BnaC06g17610D | BnASHH4.1B | C06:20191156:20193591:+ | 2436 | 966 | 321 | 9.2 | −0.666 | 9–8 | Nuclear | |
| 16 | BnaA09g38210D | BnASHH4.2A | A09:27345059:27347078:+ | 2020 | 1044 | 347 | 8.94 | −0.493 | 11–10 | Nuclear | |
| 17 | BnaC08g30180D | BnASHH4.2B | C08:30147366:30149394:+ | 2029 | 1083 | 360 | 9.1 | −0.521 | 12–11 | Nuclear | |
| 18 | BnaA08g12860D | BnASHR3.1A | A08:11310674:11313478:− | 2805 | 1485 | 494 | 8.24 | −0.487 | 11–10 | Extracellular | |
| 19 | BnaC03g67870D | BnASHR3.1B | C03:57480155:57482886:− | 2732 | 1488 | 495 | 8.59 | −0.464 | 11–10 | Extracellular | |
| 20 | BnaA07g33020D | BnASHH1.1A | A07:22719804:22721972:− | 2169 | 1386 | 461 | 5.07 | −0.682 | 8–7 | Nuclear | |
| 21 | BnaC06g37540D | BnASHH1.1B | C06:35510189:35512505:− | 2317 | 1386 | 461 | 5.03 | −0.656 | 8–7 | Nuclear | |
| 22 | BnaA07g17090D | BnASHH1.2A | A07:14363518:14367606:− | 4089 | 774 | 257 | 5.1 | −0.434 | 5–4 | Extracellular | |
| 23 | BnaA07g33460D | BnASHH2.1A | A07:22947562:22954942:− | 7381 | 4974 | 1657 | 5.69 | −0.765 | 17–16 | Nuclear | |
| 24 | BnaC06g38010D | BnASHH2.1B | C06:35772518:35779988:− | 7471 | 4974 | 1657 | 5.94 | −0.761 | 18–17 | Nuclear | |
| 25 | BnaA04g18180D | BnATX1.1A | III | A04:14637597:14643717:+ | 6121 | 3093 | 1030 | 8.54 | −0.477 | 24–23 | Nuclear |
| 26 | BnaC04g42250D | BnATX1.1B | C04:42853666:42859699:+ | 6034 | 3105 | 1034 | 8.31 | −0.48 | 24–23 | Nuclear | |
| 27 | BnaA09g50210D | BnATX2.1A | A09:33252913:33259540:− | 6628 | 3189 | 1062 | 8.38 | −0.514 | 26–25 | Nuclear | |
| 28 | BnaC08g44440D | BnATX2.1B | C08:37672655:37679549:+ | 6895 | 3342 | 1113 | 8.59 | −0.504 | 25–24 | Nuclear | |
| 29 | BnaA07g19000D | BnATX3.1A | A07:15365794:15371466:- | 5673 | 2895 | 964 | 7.91 | −0.534 | 26–25 | Nuclear | |
| 30 | BnaC06g18140D | BnATX3.1B | C06:20544144:20550243:- | 6100 | 2934 | 977 | 8.14 | −0.577 | 25–24 | Nuclear | |
| 31 | BnaA03g48900D | BnATX4.1A | A03:25126743:25131753:+ | 5011 | 2874 | 957 | 8.98 | −0.493 | 22–21 | Nuclear | |
| 32 | BnaC07g41090D | BnATX4.1B | C07:41096336:41101563:+ | 5228 | 2751 | 916 | 9 | −0.524 | 26–25 | Nuclear | |
| 33 | BnaA10g06740D | BnATX5.1A | A10:5263397:5269324:+ | 5928 | 2925 | 974 | 8.76 | −0.595 | 23–22 | Nuclear | |
| 34 | BnaC09g29770D | BnATX5.1B | C09:32512813:32518626:+ | 5814 | 2925 | 974 | 8.78 | −0.61 | 23–22 | Nuclear | |
| 35 | BnaA01g18730D | BnATXR3.1A | A01:10213817:10224101:− | 10,285 | 6897 | 2298 | 7.63 | −0.773 | 21–20 | Nuclear | |
| 36 | BnaC01g42140D | BnATXR3.1B | C01_random:1186565:1187739:- | 1175 | 882 | 293 | 4.76 | −0.626 | 4–3 | Cytoplasm | |
| 37 | BnaA05g13380D | BnATXR3.2A | A05:8123162:8131857:+ | 8696 | 5154 | 1717 | 6.05 | −0.576 | 20–19 | Nuclear | |
| 38 | BnaC06g11340D | BnATXR3.2B | C06:13248380:13258468:− | 10,089 | 6261 | 2086 | 6.29 | −0.796 | 21–20 | Nuclear | |
| 39 | BnaA09g16090D | BnATXR7.1A | A09:9526683:9533922:− | 7240 | 3867 | 1288 | 8.5 | −0.562 | 18–17 | Nuclear | |
| 40 | BnaC09g16850D | BnATXR7.1B | C09:13648653:13652429:− | 3777 | 1470 | 489 | 9.55 | −0.665 | 10–9 | Nuclear | |
| 41 | BnaA02g00160D | BnATXR5.1A | IV | A02:59526:61901:- | 2376 | 1170 | 389 | 8.45 | −0.46 | 5–4 | Nuclear |
| 42 | BnaC02g00720D | BnATXR5.1B | C02:298303:300331:+ | 2029 | 1068 | 355 | 7.04 | −0.316 | 6–5 | Nuclear | |
| 43 | BnaA10g22360D | BnATXR5.2A | A10:15105735:15107523:+ | 1789 | 1164 | 387 | 8.84 | −0.636 | 6–5 | Nuclear | |
| 44 | BnaC09g46870D | BnATXR5.2B | C09:46520440:46522899:+ | 2460 | 1158 | 385 | 8.91 | −0.669 | 6–5 | Nuclear | |
| 45 | BnaA02g37130D | BnATXR6.1A | A02_random:1596786:1601614:+ | 4829 | 1653 | 550 | 8.92 | −0.391 | 10–9 | Nuclear | |
| 46 | BnaCnng48300D | BnATXR6.1B | Cnn_radom:47593540:47595192:+ | 1653 | 1029 | 342 | 9 | −0.492 | 6–5 | Nuclear | |
| 47 | BnaA06g26870D | BnATXR6.2A | A06:18437953:18439411:− | 1459 | 1038 | 345 | 8.83 | −0.446 | 6–5 | Nuclear | |
| 48 | BnaC07g30100D | BnATXR6.2B | C07:34718609:34720078:+ | 1470 | 1038 | 345 | 8.75 | −0.487 | 6–5 | Nuclear | |
| 49 | BnaA03g01550D | BnSUVH1.1A | V | A03:700506:703243:− | 2738 | 1938 | 645 | 8.48 | −0.432 | 1–0 | Nuclear |
| 50 | BnaC03g01840D | BnSUVH1.1B | C03:886615:888912:− | 2298 | 1935 | 644 | 8.63 | −0.44 | 1–0 | Nuclear | |
| 51 | BnaA10g25310D | BnSUVH1.2A | A10:16309620:16314421:+ | 4802 | 3039 | 1012 | 7.06 | −0.398 | 5–4 | Nuclear | |
| 52 | BnaC09g50310D | BnSUVH1.2B | C09:48207580:48210227:+ | 2648 | 1842 | 613 | 6.31 | −0.566 | 4–3 | Nuclear | |
| 53 | BnaC02g02520D | BnSUVH1.3B | C02:1111629:1114084:+ | 2456 | 2169 | 722 | 8.68 | −0.43 | 3–2 | Chloroplast | |
| 54 | BnaC09g50300D | BnSUVH1.4B | C09:48204638:48207123:+ | 2486 | 1968 | 655 | 8.64 | −0.458 | 3–2 | Nuclear | |
| 55 | BnaA07g30600D | BnSUVH3.1A | A07:21627723:21630514:+ | 2792 | 1974 | 657 | 8.31 | −0.415 | 2–1 | Nuclear | |
| 56 | BnaC06g43880D | BnSUVH3.1B | C06_random:3270281:3273083:+ | 2803 | 1980 | 659 | 8.59 | −0.421 | 2–1 | Nuclear | |
| 57 | BnaA07g22850D | BnSUVH3.2A | A07:17311770:17314665:− | 2896 | 2001 | 666 | 8.31 | −0.454 | 1–0 | Nuclear | |
| 58 | BnaC06g23810D | BnSUVH3.2B | C06:25579994:25583205:− | 3212 | 2010 | 669 | 8.31 | −0.465 | 1–0 | Nuclear | |
| 59 | BnaC06g43300D | BnSUVH3.3B | C06_random:2755627:2758345:− | 2719 | 2010 | 669 | 8.31 | −0.456 | 1–0 | Nuclear | |
| 60 | BnaAnng14120D | BnSUVH7.1A | Ann_random:15255512:15258054:− | 2543 | 2382 | 793 | 5.44 | −0.558 | 2–1 | Nuclear | |
| 61 | BnaC06g28920D | BnSUVH7.1B | C06:30089672:30096121:+ | 6450 | 5571 | 1466 | 4.6 | −0.469 | 3–2 | Nuclear | |
| 62 | BnaA06g11960D | BnSUVH7.2A | A06:6207143:6216147:+ | 9005 | 2688 | 895 | 5.37 | −0.582 | 5–4 | Nuclear | |
| 63 | BnaA06g11990D | BnSUVH7.3A | A06:6229288:6231537:+ | 2250 | 2097 | 698 | 5.41 | −0.577 | 3–2 | Nuclear | |
| 64 | BnaAnng21540D | BnSUVH7.4A | Ann_radom:24010960:24013125:+ | 2166 | 2145 | 714 | 5.77 | −0.564 | 2–1 | Nuclear | |
| 65 | BnaA03g04610D | BnSUVH4.1A | A03:2148052:2152060:+ | 4009 | 1815 | 604 | 8.1 | −0.594 | 14–13 | Nuclear | |
| 66 | BnaC03g06140D | BnSUVH4.1B | C03:2974553:2979472:+ | 4920 | 1812 | 603 | 7.71 | −0.615 | 14–13 | Nuclear | |
| 67 | BnaA05g10320D | BnSUVH2.1A | A05:5614940:5616640:− | 1701 | 1701 | 566 | 8.71 | −0.227 | 1–0 | Mitochondria | |
| 68 | BnaC04g11240D | BnSUVH2.1B | C04:8711809:8713467:− | 1659 | 1659 | 552 | 6.48 | −0.278 | 1–0 | Nuclear | |
| 69 | BnaA04g19330D | BnSUVH2.2A | A04:15264117:15266030:+ | 1914 | 1914 | 637 | 7.15 | −0.347 | 1–0 | Nuclear | |
| 70 | BnaC04g43670D | BnSUVH2.2B | C04:43996787:43998700:+ | 1914 | 1914 | 637 | 7.84 | −0.351 | 1–0 | Nuclear | |
| 71 | BnaA09g34050D | BnSUVH9.1A | A09:25038028:25040666:- | 2639 | 1896 | 631 | 5.46 | −0.382 | 2–1 | Nuclear | |
| 72 | BnaC08g24970D | BnSUVH9.1B | C08:26899934:26902602:− | 2669 | 1902 | 633 | 5.44 | −0.382 | 2–1 | Nuclear | |
| 73 | BnaA05g32760D | BnSUVR3.1A | A05:22388231:22390304:- | 2074 | 1011 | 336 | 8.18 | −0.185 | 2–1 | Extracellular | |
| 74 | BnaC05g48060D | BnSUVR3.1B | C05:42732413:42734037:− | 1625 | 1011 | 336 | 8.32 | −0.144 | 2–1 | Nuclear | |
| 75 | BnaA05g08770D | BnSUVH5.1A | A05:4846708:4848861:− | 2154 | 2154 | 717 | 8.72 | −0.593 | 1–0 | Nuclear | |
| 76 | BnaC04g10170D | BnSUVH5.1B | C04:7828228:7830906:− | 2679 | 2166 | 721 | 8.88 | −0.609 | 1–0 | Nuclear | |
| 77 | BnaA04g13190D | BnSUVH6.1A | A04:11068672:11071386:− | 2715 | 2241 | 746 | 8.64 | −0.653 | 3–2 | Nuclear | |
| 78 | BnaC04g35290D | BnSUVH6.1B | C04:36693689:36695998:− | 2310 | 2277 | 758 | 8.7 | −0.656 | 2–1 | Nuclear | |
| 79 | BnaA02g22450D | BnSUVR2.1A | A02:14963608:14967065:+ | 3458 | 1923 | 640 | 5.18 | −0.492 | 11–10 | Nuclear | |
| 80 | BnaC02g30730D | BnSUVR2.1B | C02:32562070:32566295:+ | 4226 | 2355 | 784 | 5.68 | −0.449 | 12–11 | Nuclear | |
| 81 | BnaA06g36410D | BnSUVR2.2A | A06:23853703:23857119:− | 3417 | 2118 | 705 | 5.27 | −0.465 | 11–10 | Nuclear | |
| 82 | BnaC07g17530D | BnSUVR2.2B | C07:23844342:23848092:+ | 3751 | 2058 | 685 | 5.32 | −0.501 | 11–10 | Nuclear | |
| 83 | BnaA03g28550D | BnSUVR4.1A | A03:13930218:13933493:+ | 3276 | 1404 | 467 | 7.94 | −0.622 | 8–7 | Nuclear | |
| 84 | BnaC03g33690D | BnSUVR4.1B | C03:20516321:20518949:+ | 2629 | 1419 | 472 | 7.1 | −0.632 | 8–7 | Nuclear | |
| 85 | BnaA05g33030D | BnSUVR4.2A | A05:22506210:22509857:− | 3648 | 1674 | 557 | 5.82 | −0.582 | 9–8 | Nuclear | |
| 86 | BnaC05g47290D | BnSUVR4.2B | C05:42343619:42349146:− | 5528 | 2487 | 828 | 5.82 | −0.382 | 12–11 | Extracellular | |
| 87 | BnaA04g13850D | BnSUVR5.1A | A04:11704476:11710607:+ | 6132 | 4101 | 1366 | 6.25 | −0.473 | 11–10 | Nuclear | |
| 88 | BnaA09g41880D | BnSUVR5.2A | A09:29184943:29188659:− | 3717 | 1479 | 492 | 5.6 | −0.481 | 11–10 | Nuclear | |
| 89 | BnaC04g56280D | BnSUVR5.1B | C04_random:3971855:3978126:+ | 6272 | 4101 | 1366 | 6.25 | −0.486 | 11–10 | Nuclear | |
| 90 | BnaA07g02410D | BnASHR1.1A | VI | A07:2033534:2037030:+ | 3497 | 1452 | 483 | 7.73 | −0.269 | 14–13 | Nuclear |
| 91 | BnaCnng42550D | BnASHR1.1B | Cnn_random:41655054:41658605:− | 3552 | 1452 | 483 | 7.97 | −0.259 | 14–13 | Nuclear | |
| 92 | BnaA09g09920D | BnASHR2.1A | A09:5047113:5048261:+ | 1149 | 1149 | 382 | 4.49 | −0.368 | 1–0 | Nuclear | |
| 93 | BnaA09g29220D | BnATXR1.1A | A09:21881217:21882842:- | 1626 | 1626 | 541 | 6.16 | −0.34 | 1–0 | Cytoplasmic | |
| 94 | BnaCnng33180D | BnATXR1.1B | Cnn_radom:31543831:31545456:+ | 1626 | 1626 | 541 | 6.97 | −0.354 | 1–0 | Cytoplasmic | |
| 95 | BnaA05g19420D | BnATXR2.1A | A05:14796293:14799111:+ | 2819 | 1311 | 436 | 4.8 | −0.258 | 14–13 | Nuclear | |
| 96 | BnaCnng26330D | BnATXR2.1B | Cnn_random:24846860:24849927:− | 3068 | 1416 | 471 | 8.84 | −0.195 | 15–14 | Nuclear | |
| 97 | BnaAnng0177D | BnATXR4.1A | Ann_random:988229:990161:+ | 1933 | 990 | 329 | 7.44 | −0.094 | 7–6 | Extracellular | |
| 98 | BnaC02g01930D | BnATXR4.1B | C02:841583:843636:- | 2054 | 990 | 329 | 6.96 | −0.082 | 7–6 | Extracellular | |
| 99 | BnaA05g17880D | BnaA05g17880D | A05:12965688:12970193:+ | 4506 | 2361 | 786 | 6.59 | −0.141 | 14–13 | Nuclear | |
| 100 | BnaCnng44980D | BnaCnng44980D | A09_random:246310:248642:− | 5360 | 2748 | 915 | 6.68 | −0.101 | 16–15 | Nuclear | |
| 101 | BnaA09g52100D | BnaA09g52100D | Cnn_random:1772446:1777817:− | 2333 | 1677 | 558 | 5.83 | −0.027 | 4–3 | PM | |
| 102 | BnaCnng01720D | BnaCnng01720D | Cnn_radom:44087053:44092412:+ | 5372 | 2298 | 765 | 7.81 | −0.37 | 7–6 | Nuclear/PM | |
| 103 | BnaCnng08960D | BnSETD1.1B | VII | Cnn_random:8235749:8239114:+ | 3366 | 1716 | 571 | 4.78 | −0.203 | 15–14 | Cytoplasmic |
| 104 | BnaA09g45900D | BnSETD2.1A | A09:31291564:31293629:+ | 2066 | 1455 | 484 | 4.82 | −0.171 | 6–5 | Chloroplast | |
| 105 | BnaC08g39970D | BnSETD2.1B | C08:35400386:35402532:+ | 2147 | 1458 | 485 | 4.78 | −0.161 | 6–5 | chloroplast | |
| 106 | BnaA09g28320D | BnSETD3.1A | A09:21211771:21213718:− | 1948 | 1428 | 475 | 5.81 | −0.101 | 5–4 | Mitochondria/Cytoplasmic | |
| 107 | BnaAnng22450D | BnSETD3.2A | Ann_radom:25207576:25208943:+ | 1368 | 1101 | 366 | 5.06 | −0.119 | 4–3 | Cytoplasmic | |
| 108 | BnaC05g20890D | BnSETD3.1B | C05:14486930:14488943:+ | 2014 | 1416 | 471 | 4.87 | −0.294 | 5–4 | Mitochondria | |
| 109 | BnaA07g01600D | BnSETD4.1A | A07:1248812:1251691:+ | 2880 | 1617 | 538 | 4.87 | −0.294 | 10–9 | Cytoplasmic | |
| 110 | BnaC07g02950D | BnSETD4.1B | C07:3766179:3768855:− | 2677 | 1626 | 541 | 4.76 | −0.312 | 10–9 | Cytoplasmic | |
| 111 | BnaC01g39740D | BnSETD5.1B | C01:38283208:38286353:- | 3146 | 1524 | 507 | 8.66 | −0.388 | 12–11 | Chloroplast | |
| 112 | BnaA09g34820D | BnSETD6.1A | A09:25444044:25449106:− | 5063 | 1404 | 467 | 6.84 | −0.203 | 15–14 | Mitochondria/PM | |
| 113 | BnaC08g25980D | BnSETD6.1B | C08:27486308:27488874:− | 2567 | 930 | 309 | 5.12 | −0.121 | 11–10 | PM/Mitochondria/ Chloroplast | |
| 114 | BnaA09g35960D | BnSETD7.1A | A09:26136452:26138636:+ | 2185 | 1242 | 413 | 4.5 | −0.326 | 5–4 | Nuclear/Cytoplasmic/ER | |
| 115 | BnaA09g35970D | BnSETD7.2A | A09:26139780:26142012:+ | 2233 | 1548 | 515 | 4.9 | −0.311 | 5–4 | Nuclear/Cytoplasmic | |
| 116 | BnaC08g27460D | BnSETD7.1B | C08:28436643:28438862:+ | 2220 | 1461 | 486 | 4.72 | −0.291 | 5–4 | Nuclear/Cytoplasmic | |
| 117 | BnaA03g14450D | BnSETD8.1A | A03:6658037:6659546:+ | 1510 | 798 | 265 | 6.67 | −0.215 | 9–8 | Chloroplast | |
| 118 | BnaA04g22390D | BnSETD8.2A | A04:16863871:16865991:+ | 2121 | 999 | 332 | 5.55 | −0.297 | 12–11 | Nuclear/Cytoplasmic | |
| 119 | BnaA08g30390D | BnSETD8.3A | A08_random:1468364:1476519:+ | 8156 | 2034 | 677 | 5.74 | −0.378 | 17–16 | Nuclear | |
| 120 | BnaC03g63720D | BnSETD8.1B | C03:53197435:53200160:+ | 2726 | 1455 | 484 | 5.59 | −0.278 | 12–11 | Cytoplasmic/Mitochondrial/Chloroplast | |
| 121 | BnaA02g03480D | BnSETD10.1A | A02:1543050:1544888:+ | 1839 | 1446 | 481 | 5.16 | −0.338 | 6–5 | Cytoplasmic | |
| 122 | BnaC02g07170D | BnSETD10.1B | C02:3795126:3796969:+ | 1844 | 1440 | 479 | 5.32 | −0.375 | 6–5 | Cytoplasmic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sehrish, S.; Sumbal, W.; Xie, M.; Zhao, C.; Zuo, R.; Gao, F.; Liu, S. Genome-Wide Identification and Characterization of SET Domain Family Genes in Brassica napus L. Int. J. Mol. Sci. 2022, 23, 1936. https://doi.org/10.3390/ijms23041936
Sehrish S, Sumbal W, Xie M, Zhao C, Zuo R, Gao F, Liu S. Genome-Wide Identification and Characterization of SET Domain Family Genes in Brassica napus L. International Journal of Molecular Sciences. 2022; 23(4):1936. https://doi.org/10.3390/ijms23041936
Chicago/Turabian StyleSehrish, Sarfraz, Wahid Sumbal, Meili Xie, Chuanji Zhao, Rong Zuo, Feng Gao, and Shengyi Liu. 2022. "Genome-Wide Identification and Characterization of SET Domain Family Genes in Brassica napus L." International Journal of Molecular Sciences 23, no. 4: 1936. https://doi.org/10.3390/ijms23041936
APA StyleSehrish, S., Sumbal, W., Xie, M., Zhao, C., Zuo, R., Gao, F., & Liu, S. (2022). Genome-Wide Identification and Characterization of SET Domain Family Genes in Brassica napus L. International Journal of Molecular Sciences, 23(4), 1936. https://doi.org/10.3390/ijms23041936