
The Protective Effects of n-Butylidenephthalide on Retinal Ganglion Cells during Ischemic Injury


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
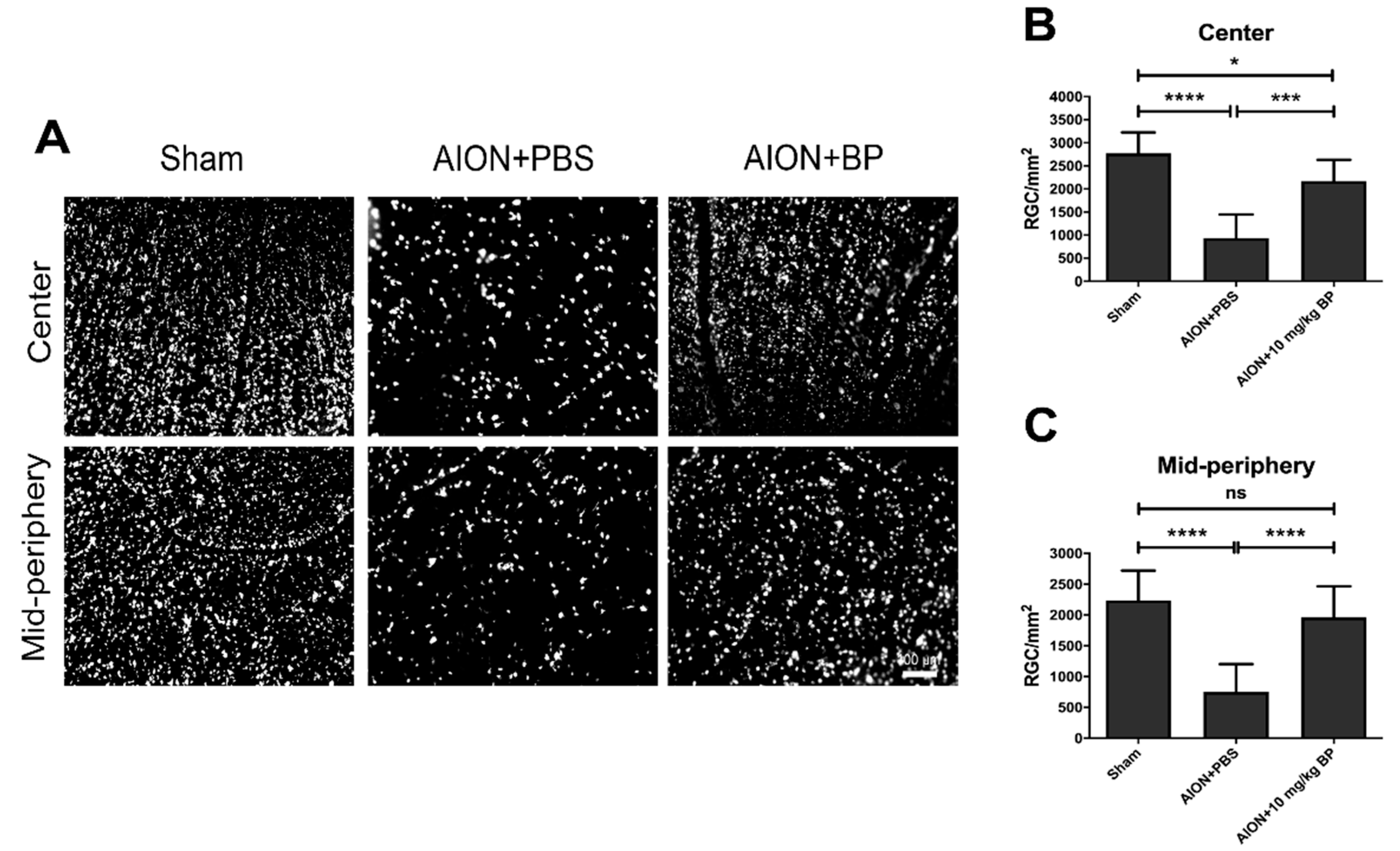
2.1. BP Rescued RGC Survival Rates
2.2. BP Preserved the Visual Function
2.3. BP Mitigated Optic Disc Edema and Preserved RNFL Thickness
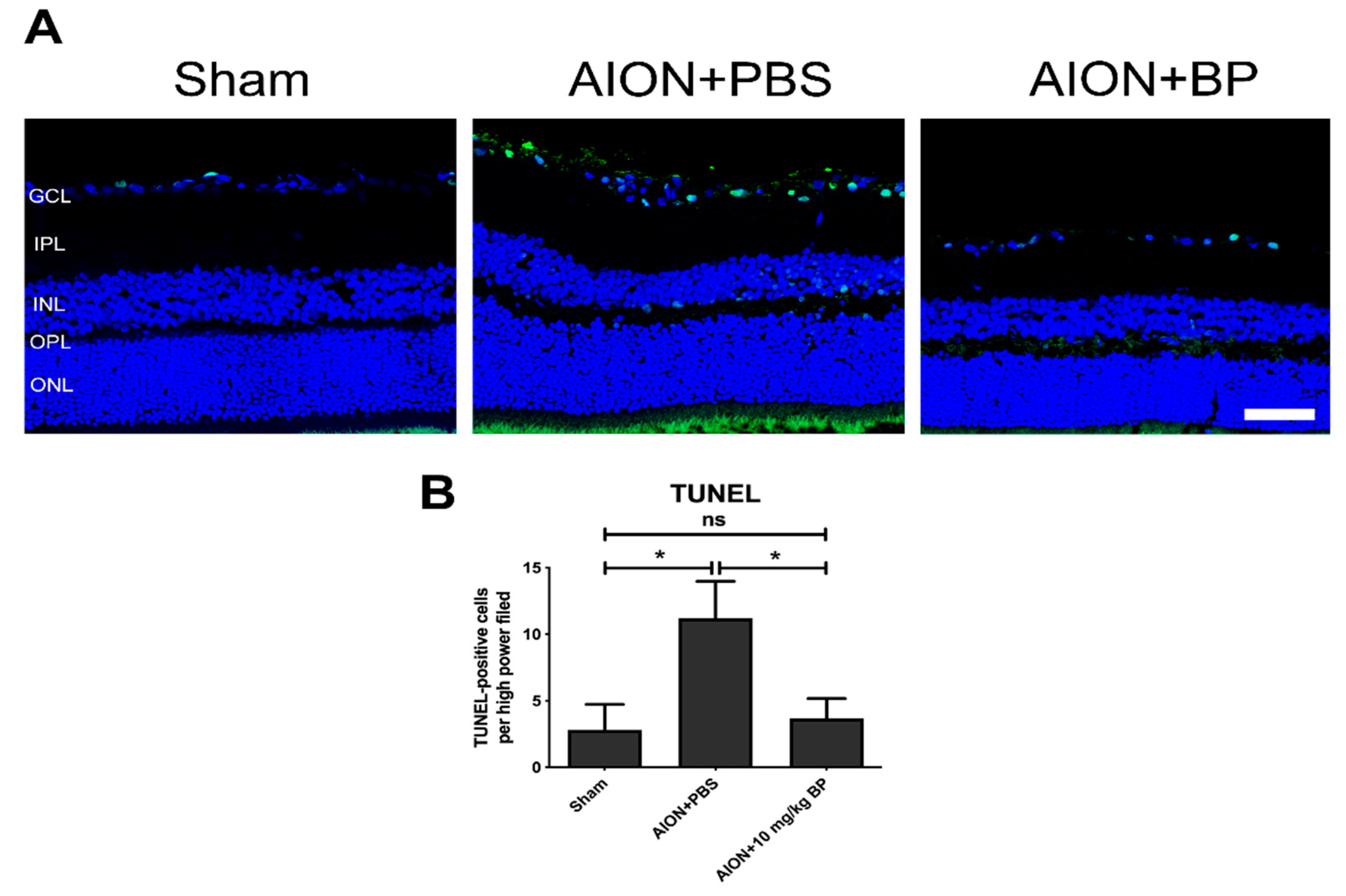
2.4. BP Reduced Apoptotic Cells in the RGC Layer
2.5. BP Reduced Macrophage Infiltration into the Optic Nerve from the Blood
2.6. BP Preserved the Completeness of the Myelin Sheath in the rAION Model
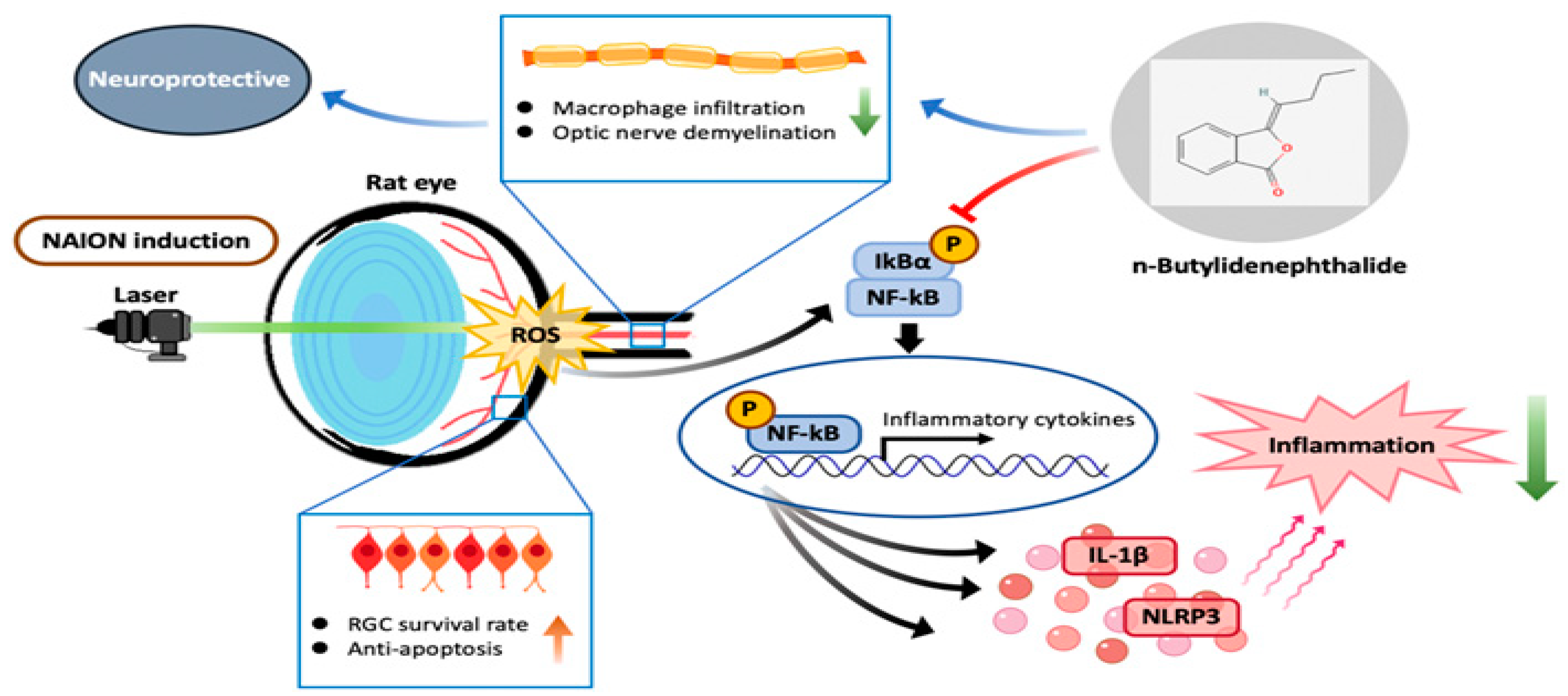
2.7. BP Inhibited the Inflammatory Response through the NF-κB Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. AION Induction
4.3. Retrograde Labeling of RGCs with Fluoro-Gold (FG)
4.4. Flash Visual Evoked Potential (FVEP)
4.5. Immunohistochemistry (IHC)
4.6. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL) Assay
4.7. Optical Coherence Tomography (OCT) Imaging
4.8. Western Blotting Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hayreh, S.S. Ischemic optic neuropathy. Prog. Retin. Eye Res. 2009, 28, 34–62. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.R.; Arnold, A.C. Current concepts in the diagnosis, pathogenesis and management of nonarteritic anterior ischaemic optic neuropathy. Eye 2015, 29, 65–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behbehani, R.; Ali, A.; Al-Moosa, A. Risk factors and visual outcome of Non-Arteritic Ischemic Optic Neuropathy (NAION): Experience of a tertiary center in Kuwait. PLoS ONE 2021, 16, e0247126. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.; Lin, W.V.; Sadaka, A.; Lee, A.G. Nonarteritic anterior ischemic optic neuropathy: Cause, effect, and management. Eye Brain 2017, 9, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Hayreh, S.S. Management of ischemic optic neuropathies. Indian J. Ophthalmol. 2011, 59, 123–136. [Google Scholar] [CrossRef]
- Bernstein, S.L.; Guo, Y.; Kelman, S.E.; Flower, R.W.; Johnson, M.A. Functional and Cellular Responses in a Novel Rodent Model of Anterior Ischemic Optic Neuropathy. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4153–4162. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Mehrabian, Z.; Bernstein, S.L. The Rodent Model of Nonarteritic Anterior Ischemic Optic Neuropathy (rNAION). J. Vis. Exp. 2016, 117, 54504. [Google Scholar] [CrossRef]
- Kapupara, K.; Wen, Y.-T.; Tsai, R.-K.; Huang, S.-P. Soluble P-selectin promotes retinal ganglion cell survival through activation of Nrf2 signaling after ischemia injury. Cell Death Dis. 2017, 8, e3172. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.C. Pathogenesis of Nonarteritic Anterior Ischemic Optic Neuropathy. J. Neuro Ophthalmol. 2003, 23, 157–163. [Google Scholar] [CrossRef]
- Atkins, E.J.; Bruce, B.B.; Newman, N.J.; Biousse, V. Treatment of Nonarteritic Anterior Ischemic Optic Neuropathy. Surv. Ophthalmol. 2010, 55, 47–63. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, S.L.; Johnson, M.A.; Miller, N.R. Nonarteritic anterior ischemic optic neuropathy (NAION) and its experimental models. Prog. Retin. Eye Res. 2011, 30, 167–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, J.D.; Leiba, H.; Goldenberg-Cohen, N. Translational Preclinical Research may Lead to Improved Medical Management of Non-Arteritic Anterior Ischemic Optic Neuropathy. Front. Neurol. 2014, 5, 122. [Google Scholar] [CrossRef] [Green Version]
- Birer, S.; Arda, H.; Kilic, D.; Baskol, G. Systemic oxidative stress in non-arteritic anterior ischemic optic neuropathy. Eye 2019, 33, 1140–1144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Guo, Y.; Miller, N.R.; Bernstein, S.L. Optic nerve infarction and post-ischemic inflammation in the rodent model of anterior ischemic optic neuropathy (rAION). Brain Res. 2009, 1264, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.-T.; Huang, C.-W.; Liu, C.-P.; Chen, C.-H.; Tu, C.-M.; Hwang, C.-S.; Chen, Y.-H.; Chen, W.-R.; Lin, K.-L.; Ho, Y.-C.; et al. Inhibition of Retinal Ganglion Cell Loss by a Novel ROCK Inhibitor (E212) in Ischemic Optic Nerve Injury Via Antioxidative and Anti-Inflammatory Actions. Investig. Ophthalmol. Vis. Sci. 2021, 62, 21. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Huang, J.; Hua, S.; Zhang, Y.; Zhang, Y.; Li, T.; Dong, L.; Gao, Q.; Fu, X. The ethnopharmacology, phytochemistry and pharmacology of Angelica biserrata—A review. J. Ethnopharmacol. 2019, 231, 152–169. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.-w.; Lin, C.-c.; Yu, Y.-l.; Lin, C.-y.; Lin, P.-c.; Wu, M.-t.; Chen, C.-j.; Chang, W.; Lin, S.-z.; Chen, Y.-l.S.; et al. n-Butylidenephthalide induced apoptosis in the A549 human lung adenocarcinoma cell line by coupled down-regulation of AP-2alpha and telomerase activity. Acta Pharmacol. Sin. 2009, 30, 1297–1306. [Google Scholar] [CrossRef]
- Su, Y.-J.; Huang, S.-Y.; Ni, Y.-H.; Liao, K.-F.; Chiu, S.-C. Anti-Tumor and Radiosensitization Effects of N-Butylidenephthalide on Human Breast Cancer Cells. Molecules 2018, 23, 240. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-Y.; Chen, Y.-C. Riboflavin immobilized Fe3O4 magnetic nanoparticles carried with n-butylidenephthalide as targeting-based anticancer agents. Artif. Cells Nanomed. Biotechnol. 2019, 47, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.-M.; Zhang, J.-J.; Li, S.; Chen, S.; Le, W.-D. n-butylidenephthalide treatment prolongs life span and attenuates motor neuron loss in SOD1(G93A) mouse model of amyotrophic lateral sclerosis. CNS Neurosci. Ther. 2017, 23, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.-M.; Harn, H.-J.; Chiou, T.-W.; Chuang, M.-H.; Chen, C.-H.; Chuang, C.-H.; Lin, P.-C.; Lin, S.-Z. Preconditioned adipose-derived stem cells ameliorate cardiac fibrosis by regulating macrophage polarization in infarcted rat hearts through the PI3K/STAT3 pathway. Lab. Investig. 2019, 99, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-M.; Harn, H.-J.; Chiou, T.-W.; Chuang, M.-H.; Chen, C.-H.; Chuang, C.-H.; Lin, P.-C.; Lin, S.-Z. Host pre-conditioning improves human adipose-derived stem cell transplantation in ageing rats after myocardial infarction: Role of NLRP3 inflammasome. J. Cell. Mol. Med. 2020, 24, 12272–12284. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.-H.; Harn, H.-J.; Liu, S.-P.; Chen, C.-S.; Chang, W.-L.; Chen, Y.-M.; Huang, J.-E.; Li, R.-J.; Tsai, S.-Y.; Hung, H.-S.; et al. n-butylidenephthalide protects against dopaminergic neuron degeneration and α-synuclein accumulation in Caenorhabditis elegans models of Parkinson’s disease. PLoS ONE 2014, 9, e85305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajamani, K.; Liu, J.-W.; Wu, C.-H.; Chiang, I.T.; You, D.-H.; Lin, S.-Y.; Hsieh, D.-K.; Lin, S.-Z.; Harn, H.-J.; Chiou, T.-W. n-Butylidenephthalide exhibits protection against neurotoxicity through regulation of tryptophan 2,3 dioxygenase in spinocerebellar ataxia type 3. Neuropharmacology 2017, 117, 434–446. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, Y.; Xu, Y.; Chen, Z.; Liu, N.; Ke, C.; Liu, B.; Wu, W. Sodium ferulate and n-butylidenephthalate combined with bone marrow stromal cells (BMSCs) improve the therapeutic effects of angiogenesis and neurogenesis after rat focal cerebral ischemia. J. Transl. Med. 2016, 14, 223. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.-H.; Liu, N.-W.; Ke, C.-C.; Liu, B.-W.; Chen, Y.-A.; Luo, C.; Zhang, Q.; Xia, Z.-Y.; Liu, R.-S. Combined treatment of sodium ferulate, n-butylidenephthalide, and ADSCs rehabilitates neurovascular unit in rats after photothrombotic stroke. J. Cell. Mol. Med. 2019, 23, 126–142. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-C.; Chen, S.-Y.; Lien, H.-Y.; Lin, S.-Z.; Lee, T.-M. Targeting the PI3K/STAT3 axis modulates age-related differences in macrophage phenotype in rats with myocardial infarction. J. Cell. Mol. Med. 2019, 23, 6378–6392. [Google Scholar] [CrossRef] [Green Version]
- Salgado, C.; Vilson, F.; Miller, N.R.; Bernstein, S.L. Cellular Inflammation in Nonarteritic Anterior Ischemic Optic Neuropathy and Its Primate Model. Arch. Ophthalmol. 2011, 129, 1583–1591. [Google Scholar] [CrossRef] [Green Version]
- Sprinkle, T.J. 2’,3’-cyclic nucleotide 3’-phosphodiesterase, an oligodendrocyte-Schwann cell and myelin-associated enzyme of the nervous system. Crit. Rev. Neurobiol. 1989, 4, 235–301. [Google Scholar]
- Lee, J.; Gravel, M.; Zhang, R.; Thibault, P.; Braun, P.E. Process outgrowth in oligodendrocytes is mediated by CNP, a novel microtubule assembly myelin protein. J. Cell Biol. 2005, 170, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Dratviman-Storobinsky, O.; Hasanreisoglu, M.; Offen, D.; Barhum, Y.; Weinberger, D.; Goldenberg-Cohen, N. Progressive damage along the optic nerve following induction of crush injury or rodent anterior ischemic optic neuropathy in transgenic mice. Mol. Vis. 2008, 14, 2171–2179. [Google Scholar] [PubMed]
- Ettle, B.; Schlachetzki, J.C.M.; Winkler, J. Oligodendroglia and Myelin in Neurodegenerative Diseases: More Than Just Bystanders? Mol. Neurobiol. 2016, 53, 3046–3062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mot, A.I.; Depp, C.; Nave, K.-A. An emerging role of dysfunctional axon-oligodendrocyte coupling in neurodegenerative diseases. Dialogues Clin. Neurosci. 2018, 20, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Raffaele, S.; Boccazzi, M.; Fumagalli, M. Oligodendrocyte Dysfunction in Amyotrophic Lateral Sclerosis: Mechanisms and Therapeutic Perspectives. Cells 2021, 10, 565. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z.-G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, A.; Namekata, K.; Guo, X.; Noro, T.; Harada, C.; Harada, T. Targeting Oxidative Stress for Treatment of Glaucoma and Optic Neuritis. Oxidative Med. Cell. Longev. 2017, 2017, 2817252. [Google Scholar] [CrossRef]
- Masuda, T.; Shimazawa, M.; Hara, H. Retinal Diseases Associated with Oxidative Stress and the Effects of a Free Radical Scavenger (Edaravone). Oxidative Med. Cell. Longev. 2017, 2017, 9208489. [Google Scholar] [CrossRef]
- Dogru, M.; Kojima, T.; Simsek, C.; Tsubota, K. Potential Role of Oxidative Stress in Ocular Surface Inflammation and Dry Eye Disease. Investig. Ophthalmol. Vis. Sci. 2018, 59, DES163–DES168. [Google Scholar] [CrossRef] [Green Version]
- Cejka, C.; Cejkova, J. Oxidative Stress to the Cornea, Changes in Corneal Optical Properties, and Advances in Treatment of Corneal Oxidative Injuries. Oxidative Med. Cell. Longev. 2015, 2015, 591530. [Google Scholar] [CrossRef] [Green Version]
- Chiras, D.; Kitsos, G.; Petersen, M.B.; Skalidakis, I.; Kroupis, C. Oxidative stress in dry age-related macular degeneration and exfoliation syndrome. Crit. Rev. Clin. Lab. Sci. 2015, 52, 12–27. [Google Scholar] [CrossRef]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS Crosstalk in Inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, J.; Wang, J.; Yang, B.; Weng, Q.; He, Q. Targeting Microglia and Macrophages: A Potential Treatment Strategy for Multiple Sclerosis. Front. Pharmacol. 2019, 10, 286. [Google Scholar] [CrossRef] [Green Version]
- Block, M.L.; Zecca, L.; Hong, J.-S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms Underlying Inflammation in Neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef] [Green Version]
- Chitnis, T.; Weiner, H.L. CNS inflammation and neurodegeneration. J. Clin. Investig. 2017, 127, 3577–3587. [Google Scholar] [CrossRef] [Green Version]
- Hickman, S.; Izzy, S.; Sen, P.; Morsett, L.; El Khoury, J. Microglia in neurodegeneration. Nat. Neurosci. 2018, 21, 1359–1369. [Google Scholar] [CrossRef]
- Slater, B.J.; Vilson, F.L.; Guo, Y.; Weinreich, D.; Hwang, S.; Bernstein, S.L. Optic Nerve Inflammation and Demyelination in a Rodent Model of Nonarteritic Anterior Ischemic Optic Neuropathy. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7952–7961. [Google Scholar] [CrossRef]
- Guo, H.; Callaway, J.B.; Ting, J.P.Y. Inflammasomes: Mechanism of action, role in disease, and therapeutics. Nat. Med. 2015, 21, 677–687. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Pei, L.; Yao, S.; Wu, Y.; Shang, Y. NLRP3 Inflammasome in Neurological Diseases, from Functions to Therapies. Front. Cell. Neurosci. 2017, 11, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.; Barnett, M.H.; Yiannikas, C.; Parratt, J.; Matthews, J.; Graham, S.L.; Klistorner, A. Chronic demyelination exacerbates neuroaxonal loss in patients with MS with unilateral optic neuritis. Neurol. Neuroimmunol. Neuroinflamm. 2020, 7, e700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marmoy, O.R.; Viswanathan, S. Clinical electrophysiology of the optic nerve and retinal ganglion cells. Eye 2021, 35, 2386–2405. [Google Scholar] [CrossRef] [PubMed]
- Meyer, R.; Weissert, R.; Diem, R.; Storch, M.K.; de Graaf, K.L.; Kramer, B.; Bähr, M. Acute Neuronal Apoptosis in a Rat Model of Multiple Sclerosis. J. Neurosci. 2001, 21, 6214. [Google Scholar] [CrossRef]
- Shindler, K.S.; Ventura, E.; Dutt, M.; Rostami, A. Inflammatory demyelination induces axonal injury and retinal ganglion cell apoptosis in experimental optic neuritis. Exp. Eye Res. 2008, 87, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.-L.; Wen, Y.; Chang, C.; Chang, S.; Lin, K.; Tsai, R. Early Methylprednisolone Treatment Can Stabilize the Blood-Optic Nerve Barrier in a Rat Model of Anterior Ischemic Optic Neuropathy (rAION). Investig. Ophthalmol. Vis. Sci. 2017, 58, 1628–1636. [Google Scholar] [CrossRef] [Green Version]
- Hayreh, S.S.; Zimmerman, M.B. Non-arteritic anterior ischemic optic neuropathy: Role of systemic corticosteroid therapy. Graefe Arch. Clin. Exp. Ophthalmol. 2008, 246, 1029–1046. [Google Scholar] [CrossRef]
- Black, R.L.; Oglesby, R.B.; von Sallmann, L.; Bunim, J.J. Posterior Subcapsular Cataracts Induced by Corticosteroids in Patients with Rheumatoid Arthritis. JAMA 1960, 174, 166–171. [Google Scholar] [CrossRef]
- Kersey, J.P.; Broadway, D.C. Corticosteroid-induced glaucoma: A review of the literature. Eye 2006, 20, 407–416. [Google Scholar] [CrossRef] [Green Version]
- McGhee, C.N.J.; Dean, S.; Danesh-Meyer, H. Locally Administered Ocular Corticosteroids. Drug Saf. 2002, 25, 33–52. [Google Scholar] [CrossRef]
- Zhou, Y.; Cui, C.; Ma, X.; Luo, W.; Zheng, S.G.; Qiu, W. Nuclear Factor κB (NF-κB)–Mediated Inflammation in Multiple Sclerosis. Front. Immunol. 2020, 11, 391. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, Y.-Y.; Chien, J.-Y.; Ciou, J.-W.; Huang, S.-P. The Protective Effects of n-Butylidenephthalide on Retinal Ganglion Cells during Ischemic Injury. Int. J. Mol. Sci. 2022, 23, 2095. https://doi.org/10.3390/ijms23042095
Chou Y-Y, Chien J-Y, Ciou J-W, Huang S-P. The Protective Effects of n-Butylidenephthalide on Retinal Ganglion Cells during Ischemic Injury. International Journal of Molecular Sciences. 2022; 23(4):2095. https://doi.org/10.3390/ijms23042095
Chicago/Turabian StyleChou, Yu-Yau, Jia-Ying Chien, Jhih-Wei Ciou, and Shun-Ping Huang. 2022. "The Protective Effects of n-Butylidenephthalide on Retinal Ganglion Cells during Ischemic Injury" International Journal of Molecular Sciences 23, no. 4: 2095. https://doi.org/10.3390/ijms23042095
APA StyleChou, Y. -Y., Chien, J. -Y., Ciou, J. -W., & Huang, S. -P. (2022). The Protective Effects of n-Butylidenephthalide on Retinal Ganglion Cells during Ischemic Injury. International Journal of Molecular Sciences, 23(4), 2095. https://doi.org/10.3390/ijms23042095