
Genome-Wide Identification, Classification, and Expression Analysis of the HD-Zip Transcription Factor Family in Apple (Malus domestica Borkh.)

Abstract
:1. Introduction
2. Results
2.1. Identification of MdHDZ Genes
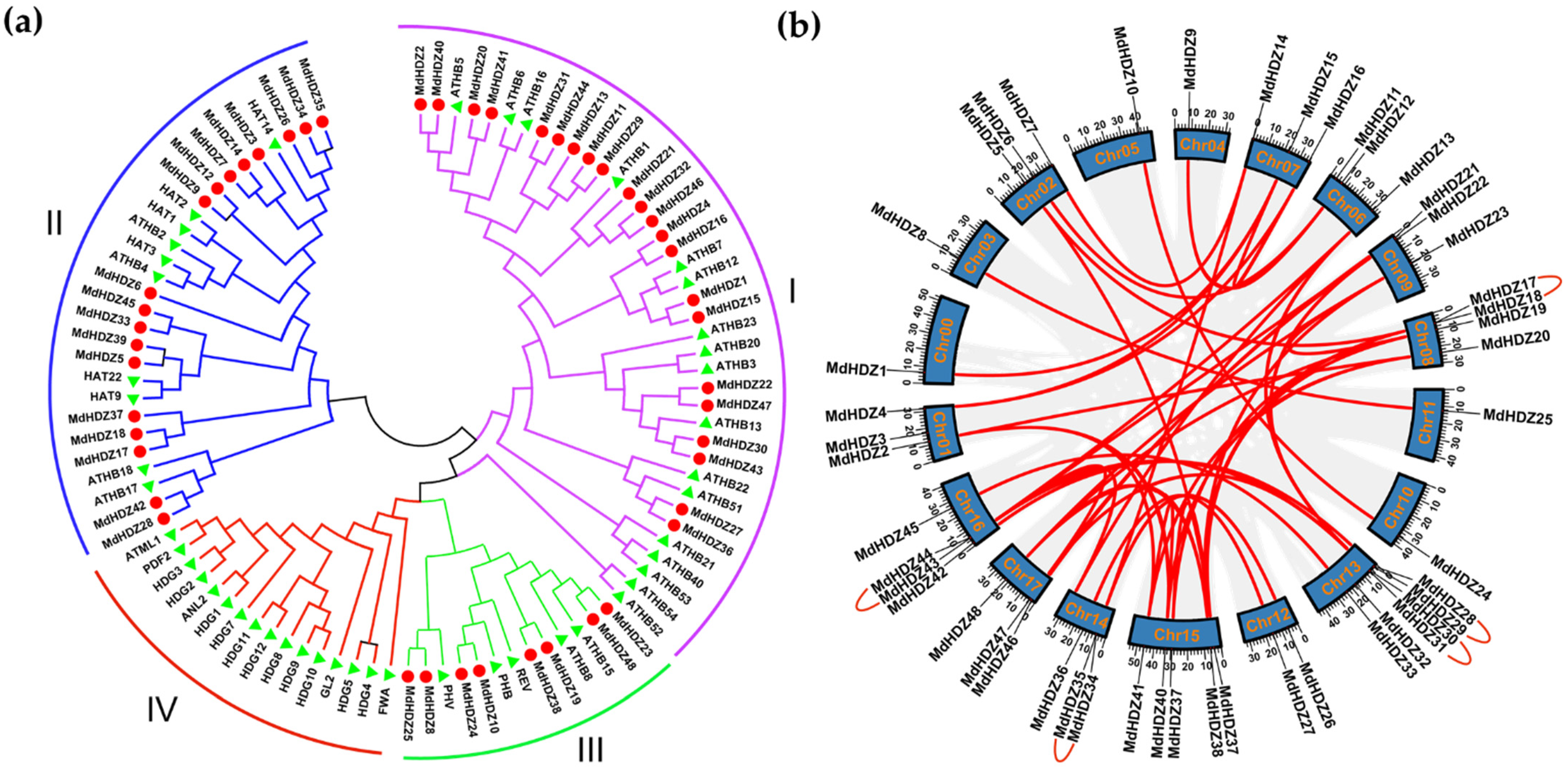
2.2. Phylogenetic and Synteny Analyses of MdHDZ Genes
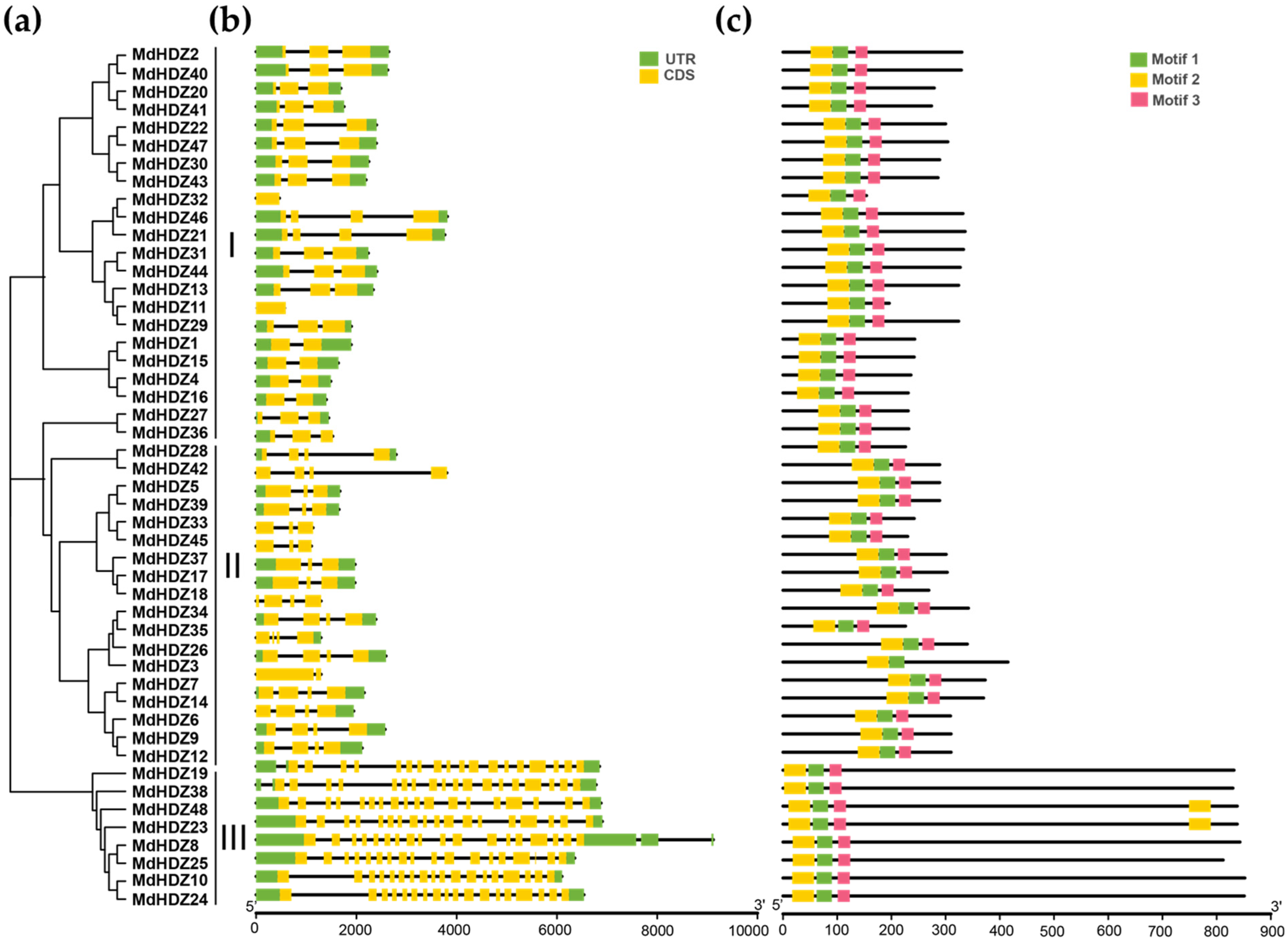
2.3. Gene Structure and Conserved Motif Analyses of MdHDZ Genes
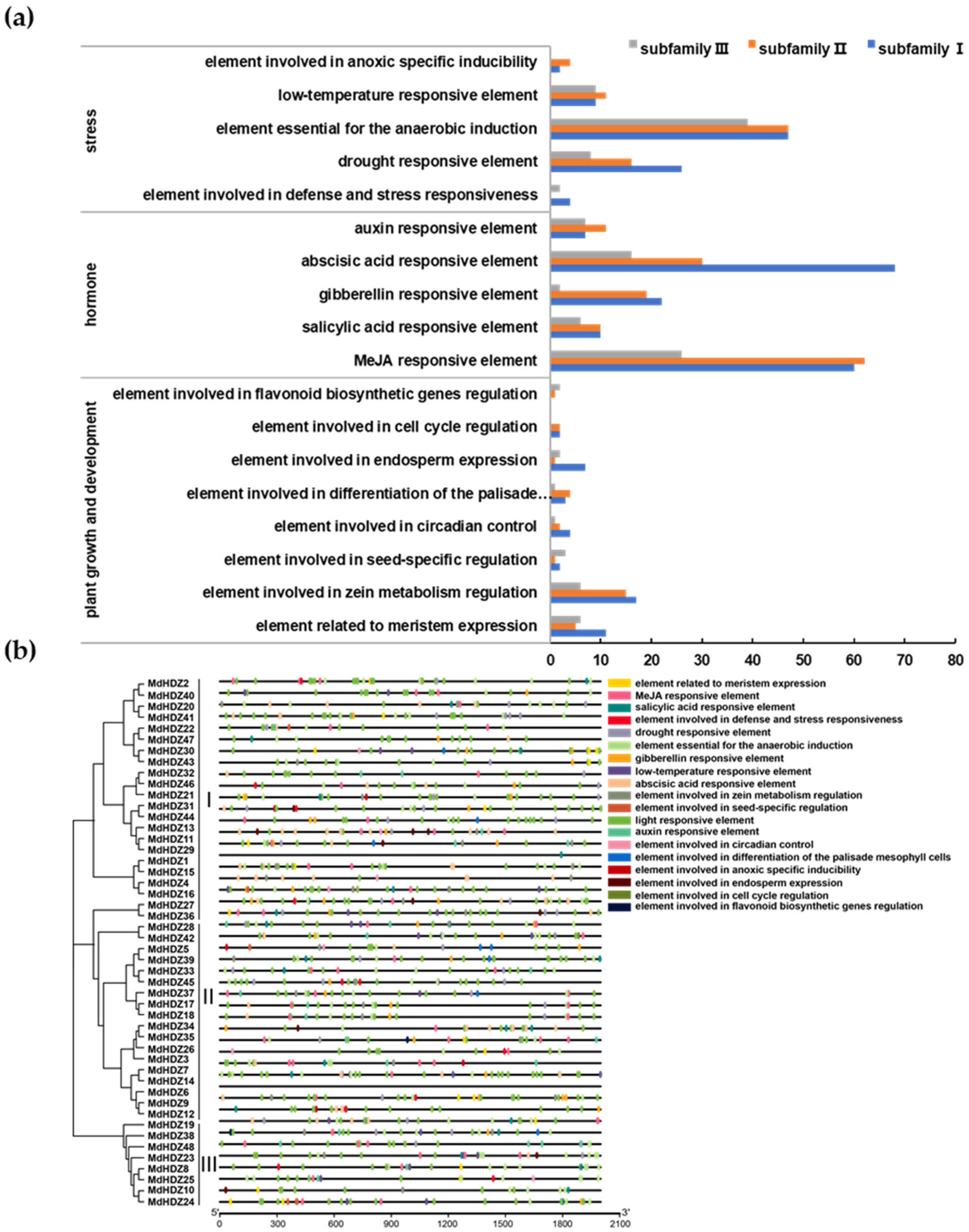
2.4. Cis-Regulatory Element Analysis of MdHDZ Genes
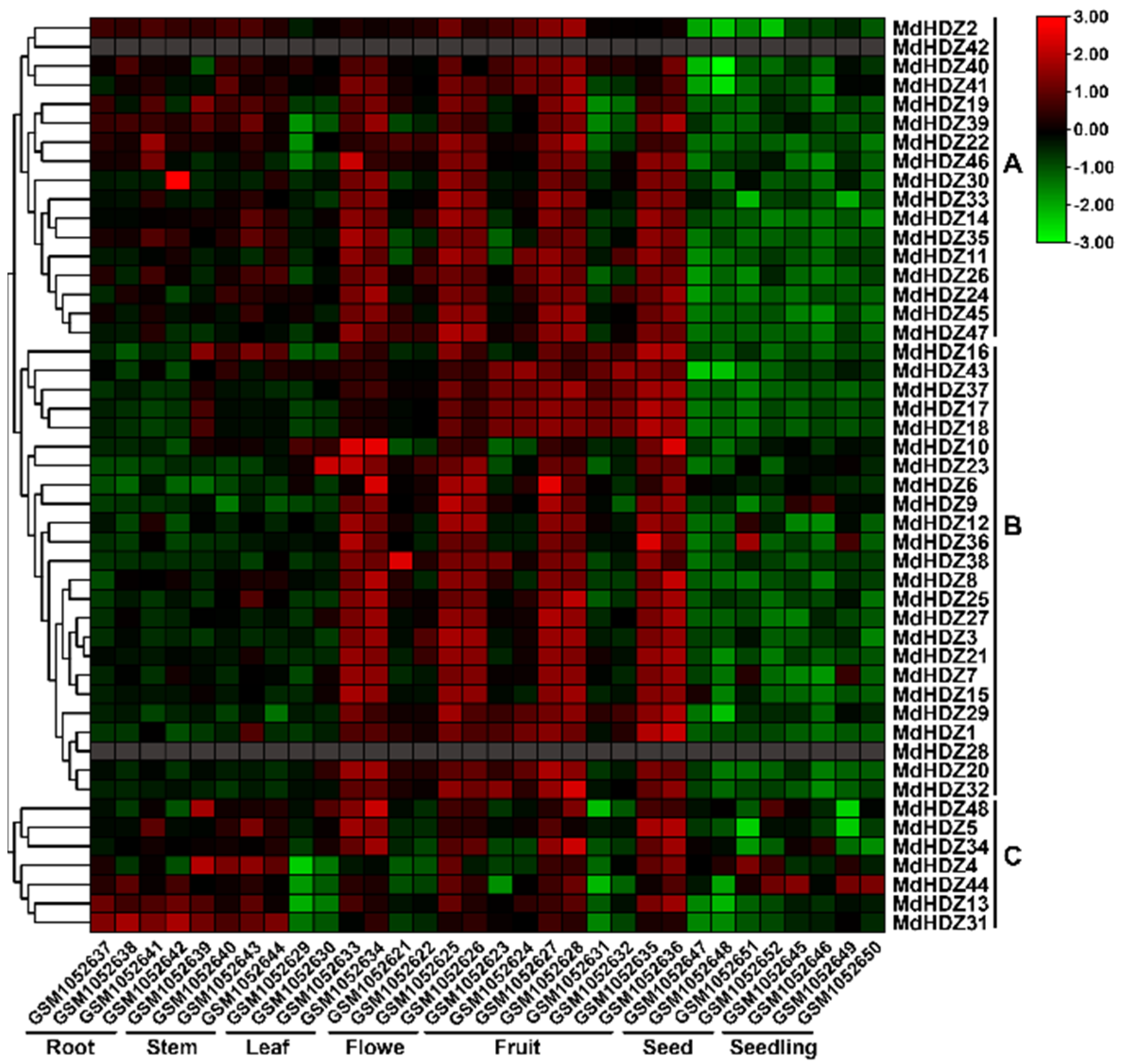
2.5. Tissue-Specific Expression Pattern of MdHDZ Genes
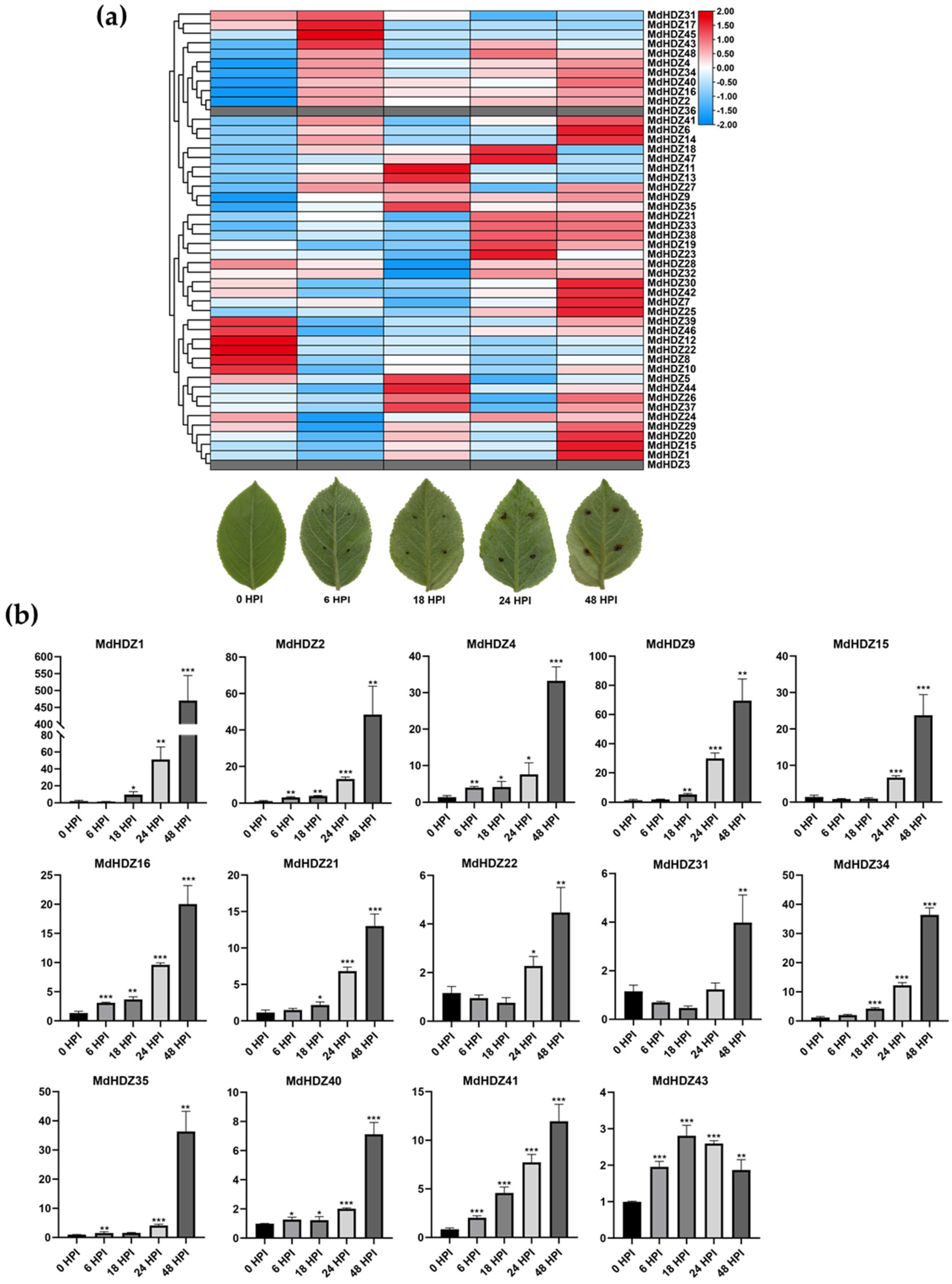
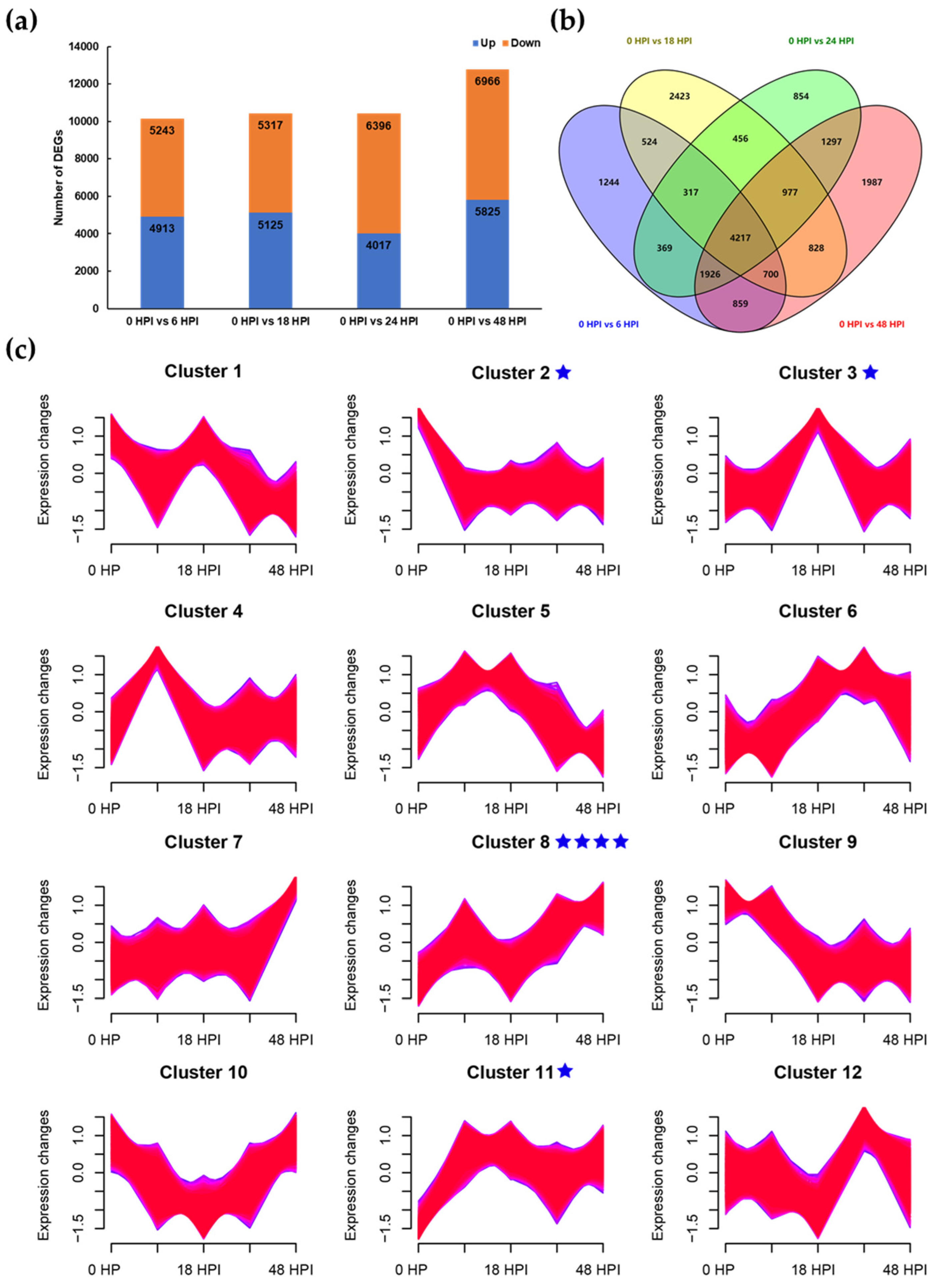
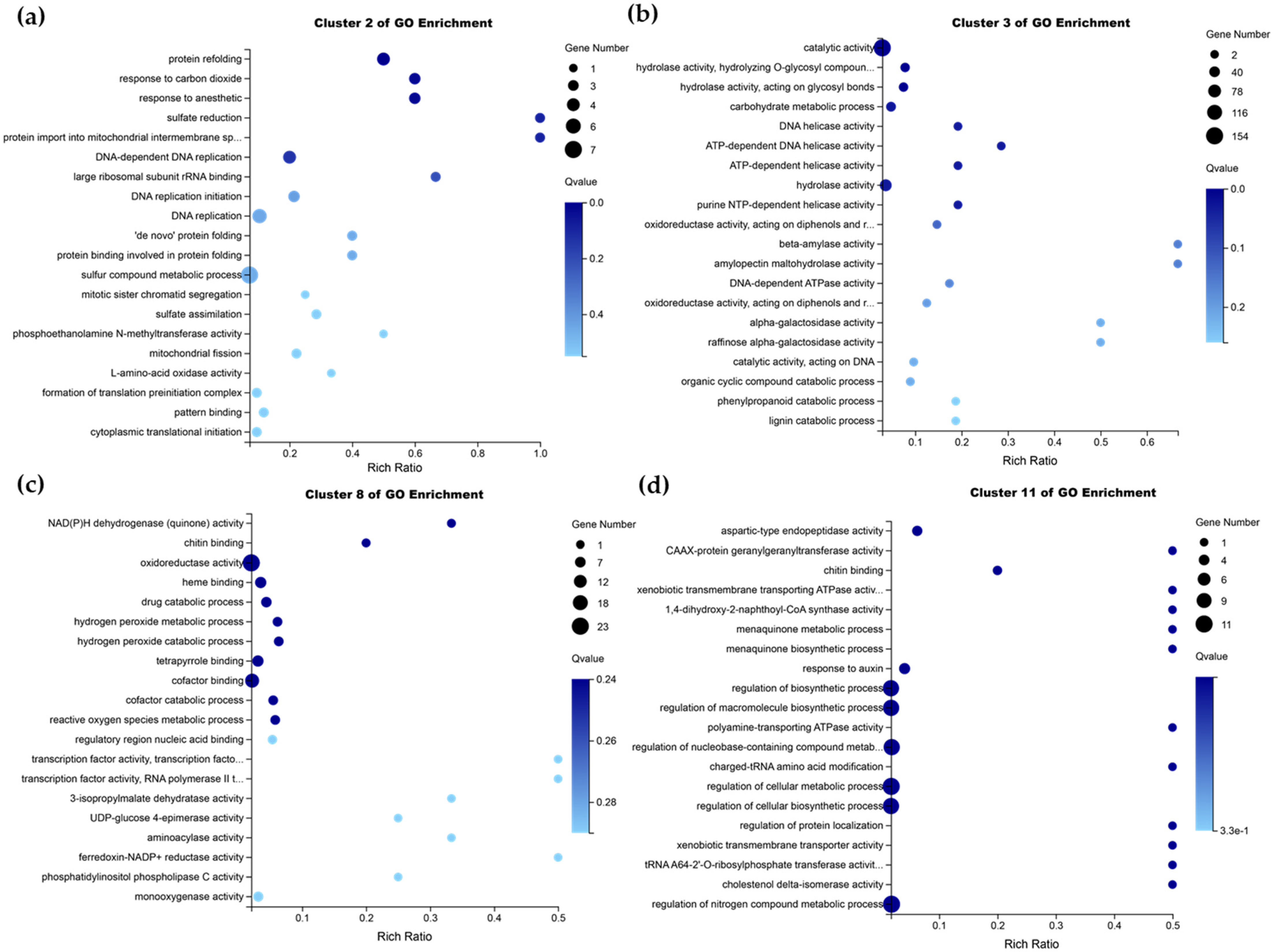
2.6. Transcriptome Analysis of Apple Leaf Response to AAAP Infection
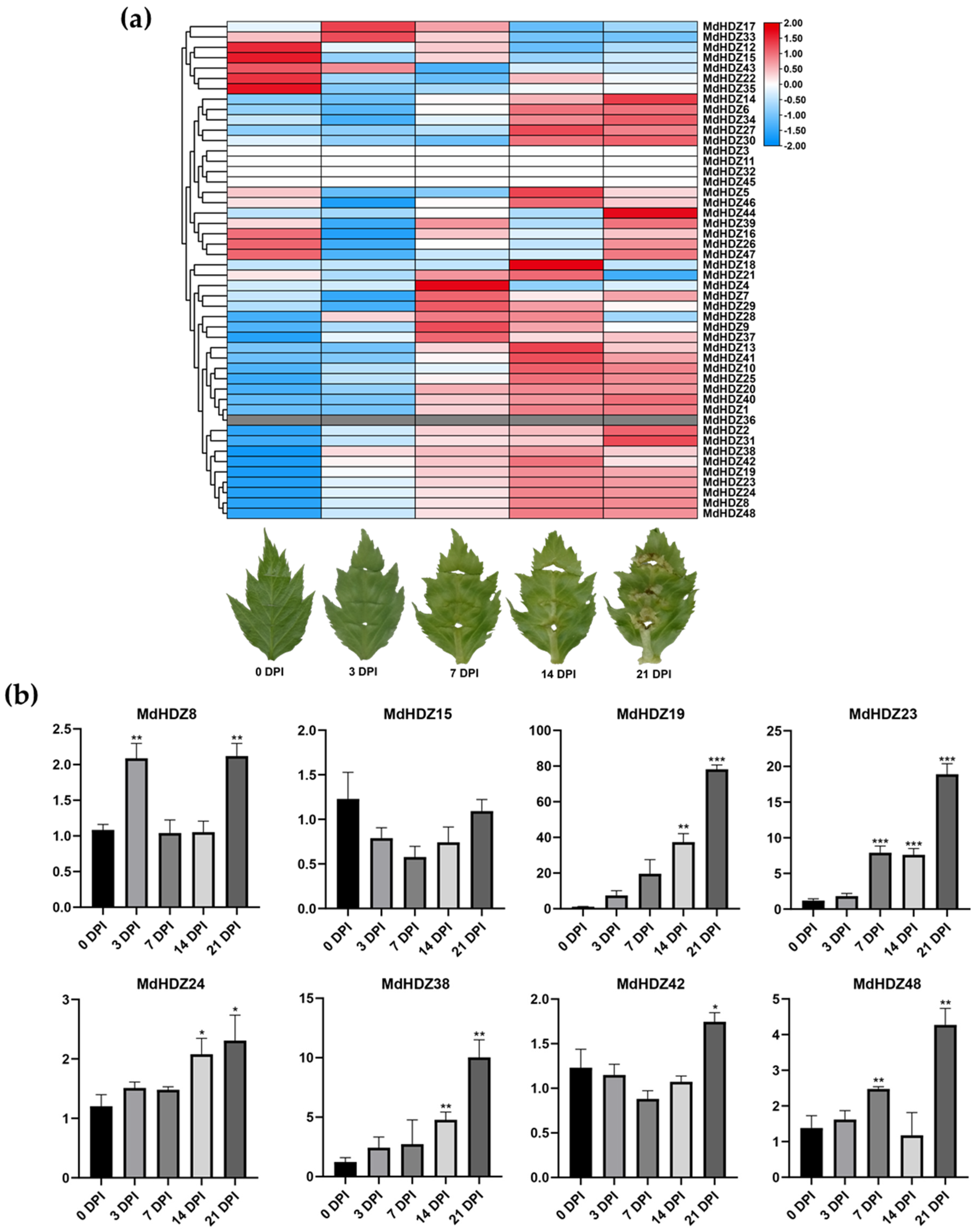
2.7. Expression Profiles of MdHDZ Genes in Adventitious Bud Regeneration from Apple Leaves In Vitro
3. Discussion
4. Materials and Methods
4.1. Identification of HD-Zip Genes in The Apple Genome
4.2. Phylogenetic and Synteny Analyses
4.3. Gene Structure and Conserved Motif Analysis
4.4. Promoter Analysis
4.5. Tissue-Specific Expression Pattern Analysis
4.6. Plant Materials and Fungus Inoculation Method
4.7. RNA Sequencing and Data Analysis
4.8. qRT-PCR Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eva Henriksson, A.S.B.O. Homeodomain Leucine Zipper Class I Genes in Arabidopsis. Expression Patterns and Phylogenetic Relationships. Plant Physiol. 2005, 139, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.A.; Medford, J.I.; Barton, M.K.; Moan, E.I. A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Chan, R.L.; Gago, G.M.; Palena, C.M.; Gonzalez, D.H. Homeoboxes in plant development. Biochim. Biophys. Acta (BBA)—Gene Struct. Expr. 1998, 1442, 1–19. [Google Scholar] [CrossRef]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Appiano, M.; Huibers, R.P.; Chen, X.; Bai, Y. Activation tagging of ATHB13 in Arabidopsis thaliana confers broad-spectrum disease resistance. Plant Mol. Biol. 2014, 86, 641–653. [Google Scholar] [CrossRef]
- Romani, F.; Ribone, P.A.; Capella, M.; Miguel, V.N.; Chan, R.L. A matter of quantity: Common features in the drought response of transgenic plants overexpressing HD-Zip I transcription factors. Plant Sci. 2016, 139–154. [Google Scholar] [CrossRef]
- Sur2 mutations of Arabidopsis thaliana define a new locus involved in the control of auxin homeostasis. Plant J. 2010, 14, 603–611.
- Rueda, E.C. Hahb-10, a sunflower homeobox-leucine zipper gene, is regulated by light quality and quantity, and promotes early flowering when expressed in Arabidopsis. Plant Cell Physiol. 2005, 46, 1954–1963. [Google Scholar] [CrossRef]
- Prigge, M.J. Class III Homeodomain-Leucine Zipper Gene Family Members Have Overlapping, Antagonistic, and Distinct Roles in Arabidopsis Development. Plant Cell Online 2005, 17, 61–76. [Google Scholar] [CrossRef] [Green Version]
- Baima, S. The Arabidopsis ATHB-8 HD-Zip Protein Acts as a Differentiation-Promoting Transcription Factor of the Vascular Meristems. Plant Physiol. 2001, 126, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Mattsson, J.; Ckurshumova, W.; Berleth, T. Auxin Signaling in Arabidopsis Leaf Vascular Development. Plant Physiol. 2003, 131, 1327–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, M.; Katsumata, H.; Abe, M.; Yabe, N.; Kome Da, Y.; Yamamoto, K.T.; Takahashi, T. Characterization of the class IV homeodomain-Leucine Zipper gene family in Arabidopsis. Plant Physiol. 2006, 141, 1363–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wang, S.; Xiao, Y.; Li, Z.; Yang, J. Functional characterization of a HD-ZIP IV transcription factor NtHDG2 in regulating flavonols biosynthesis in Nicotiana tabacum. Plant Physiol. Bioch. 2019, 146, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Kuai, B.; Hu, P.; Ding, Y. Ectopic-overexpression of an HD-Zip IV transcription factor from Ammopiptanthus mongolicus (Leguminosae) promoted upward leaf curvature and non-dehiscent anthers in Arabidopsis thaliana. Plant Cell Tissue Organ Cult. 2012, 110, 299–306. [Google Scholar] [CrossRef]
- Oshima, R.; Dagallier, B.; Kearns, P.W.E. OECD Consensus Document on the Biology of Apple (Malus domestica Borkh.). Ser. Harmon. Regul. Overs. Biotechnol. 2019, 23, 1015–1023. [Google Scholar]
- Chen, Z.; Yu, L.; Liu, W.; Zhang, J.; Chen, X. Research Progress of Fruit Color Development in Apple (Malus domestica Borkh.). Plant Physiol. Bioch 2021, 162, 267–279. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, C.; Zhang, Y.; Gu, Z.; Li, T. CORRECTION: A Single-Nucleotide Polymorphism in the Promoter of a Hairpin RNA Contributes to Alternaria alternata Leaf Spot Resistance in Apple (Malus × domestica). Plant Cell 2019, 30, 1924–1942. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Ni, W.; Liu, S.; Cai, B.; Han, X.; Wang, S. Transcriptomics Analysis of Apple Leaves in Response to Alternaria alternata Apple Pathotype Infection. Front. Plant Sci. 2017, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Valdés, A.E.; Verns, E.; Johansson, H.; Rada-Iglesias, A.; Engstrm, P. The homeodomain-leucine zipper (HD-Zip) class I transcription factors ATHB7 and ATHB12 modulate abscisic acid signalling by regulating protein phosphatase 2C and abscisic acid receptor gene activities. Plant Mol. Biol. 2012, 80, 405–418. [Google Scholar] [CrossRef]
- Zhang, S.; Haider, I.; Kohlen, W.; Li, J.; Ouwerkerk, P. Function of the HD-Zip I gene OsHOX22 in ABA-mediated drought and salt tolerances in rice. Plant Mol. Biol. 2012, 80, 571–585. [Google Scholar] [CrossRef]
- Zhao, S.; Gao, H.; Jia, X.; Li, X.; Ma, F. The γ-clade HD-Zip I transcription factor MdHB-7 regulates salt tolerance in transgenic apple (Malus domestica). Plant Soil 2021, 172, 1452–1464. [Google Scholar] [CrossRef]
- Mou, S.; Liu, Z.; Gao, F.; Yang, S.; Su, M.; Shen, L.; Wu, Y.; He, S. CaHDZ27, a Homeodomain-Leucine Zipper I (HD-Zip I) Protein, Positively Regulates the Resistance to Ralstonia solanacearum Infection in Pepper. Mol. Plant Microbe Interact. 2017, 30, 960–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, H.; Luo, X.; Wang, T.; Liu, S.; Zhu, L. GhHB12 negatively regulates abiotic stress tolerance in Arabidopsis and cotton. Environ. Exp. Bot. 2020, 176, 104087. [Google Scholar]
- Wang; Hong; Li; Xiao; Gang; Lin; Jing; Chang; Youhong, Genome-wide identification of pear HD-Zip gene family and expression patterns under stress induced by drought, salinity, and pathogen. Acta Physiol. Plant 2015, 37, 189. [CrossRef] [Green Version]
- Lin, X.; Kaul, S.; Rounsley, S.; Shea, T.P.; Benito, M.I.; Town, C.D.; Fujii, C.Y.; Mason, T.; Bowman, C.L.; Barnstead, M. Sequence and analysis of chromosome 2 of the plant. Nature 1999, 402, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.A.; Kingston, R.E. Mechanisms of polycomb gene silencing: Knowns and unknowns. Nat. Rev. Mol. Cell Biol. 2009, 10, 697. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 2010, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- James, D.J.; Passey, A.J.; Barbara, D.J.; Bevan, M.W. Genetic transformation of apple (Malus pumila Mill.) using a disarmed Ti-binary vector. Plant Cell Rep. 1989, 7, 658–661. [Google Scholar]
- Zhang, Z.; Jing, S.; Wang, G.; Fang, H. Genetic Transformation of the Commercial Apple Cultivar New Jonagold and Regeneration of Its Transgenic Plants. Acta Hortic. Sin. 1997, 24, 378–380. [Google Scholar]
- Seong, E.S.; Song, K.J.; Jegal, S.; Chang, Y.Y.; Chung, I.M. Silver nitrate and aminoethoxyvinylglycine affect Agrobacterium-mediated apple transformation. Plant Growth Regul. 2005, 45, 75–82. [Google Scholar] [CrossRef]
- Mayer, K.F.; Schoof, H.; Haecker, A.; Lenhard, M.; Jürgens, G.; Laux, T. Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 1998, 95, 805. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.Q.; Lian, H.; Zhou, C.M.; Xu, L.; Jiao, Y.; Wang, J.W. A Two-Step Model for de novo Activation of WUSCHEL during Plant Shoot Regeneration. Plant Cell 2017, 1073, 1073–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roodbarkelari, F.; Du, F.; Truernit, E.; Laux, T. ZLL/AGO10 maintains shoot meristem stem cells during Arabidopsis embryogenesis by down-regulating ARF2-mediated auxin response. BMC Biol. 2015, 13, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarbelli, A.R.; Ciolfi, A.; Salvucci, S.; Ruzza, V.; Possenti, M.; Carabelli, M.; Fruscalzo, A.; Sessa, G.; Morelli, G.; Ruberti, I. The Arabidopsis Homeodomain-leucine Zipper II gene family: Diversity and redundancy. Plant Mol. Biol. 2008, 68, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, Z.; Zhao, H.; Zhao, Y.; Cheng, B.; Xiang, Y. Genome-wide analysis of soybean HD-Zip gene family and expression profiling under salinity and drought treatments. PLoS ONE 2014, 9, e87156. [Google Scholar]
- Ge, X.X.; Liu, Z.; Wu, X.M.; Chai, L.J.; Guo, W.W. Genome-wide identification, classification and analysis of HD-ZIP gene family in citrus, and its potential roles in somatic embryogenesis regulation. Gene 2015, 574, 61–68. [Google Scholar] [CrossRef]
- Sharif, R.; Xie, C.; Wang, J.; Cao, Z.; Zhang, H.; Chen, P.; Yuhong, L. Genome wide identification, characterization and expression analysis of HD-ZIP gene family in Cucumis sativus L. under biotic and various abiotic stresses. Int. J. Biol. Macromol. 2020, 158, 502–520. [Google Scholar] [CrossRef]
- Li, W.; Dong, J.; Cao, M.; Gao, X.; Wang, D.; Liu, B.; Chen, Q. Genome-wide identification and characterization of HD-ZIP genes in potato. Gene 2019, 697, 103–117. [Google Scholar] [CrossRef]
- Yue, H.; Shu, D.; Wang, M.; Xing, G.; Zhan, H.; Du, X.; Song, W.; Nie, X. Genome-Wide Identification and Expression Analysis of the HD-Zip Gene Family in Wheat (Triticum aestivum L.). Genes 2018, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Gao, Z.; Li, R.; Xu, Y.; Kong, Y.; Zhou, G.; Meng, C.; Hu, R. Genome-wide identification and expression profiling of HD-ZIP gene family in Medicago truncatula. Genomics 2020, 112, 3624–3635. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, C.; Guo, Y.; Niu, W.; Wang, Y.; Xu, Y. Evolution and expression analysis reveal the potential role of the HD-Zip gene family in regulation of embryo abortion in grapes (Vitis vinifera L.). Bmc Genom. 2017, 18, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Pruss, D. The genome of the domesticated apple (Malus [times] domestica Borkh.). Nat Genet 2010, 42, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Turchi, L.; Carabelli, M.; Ruzza, V.; Possenti, M.; Sassi, M.; Penalosa, A.; Sessa, G.; Salvi, S.; Forte, V.; Morelli, G. Arabidopsis HD-Zip II transcription factors control apical embryo development and meristem function. Development 2013, 140, 2118–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, L.; ShanShan, H.; CaiXia, Z.; LiYi, Z.; ShuXun, B.; GaoPeng, Y.; WuXing, L.; LiQun, K.; PeiHua, C.; XiaoLei, H. Identification and analysis of differentially expressed genes in adventitious shoot regeneration in leaves of apple. Sci. Agric. Sin. 2021, 54, 3488–3501. [Google Scholar]
- Long, M. Gene Duplication and Evolution. Science 2001, 293, 1551. [Google Scholar] [CrossRef] [Green Version]
- Meyers, B.C.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R.W. Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis. Plant Cell Online 2003, 15, 809–834. [Google Scholar] [CrossRef] [Green Version]
- Capella, M.; Ribone, P.A.; Arce, A.L.; Chan, R.L. Arabidopsis thaliana HomeoBox 1 (AtHB1), a Homedomain-Leucine Zipper I (HD-Zip I) transcription factor, is regulated by Phytochrome-Interacting Factor 1 to promote hypocotyl elongation. New Phytol. 2015, 207, 669–682. [Google Scholar] [CrossRef]
- Haas, B.J.; Salzberg, S.L.; Wei, Z.; Pertea, M. Automated eukaryotic gene structure annotation using EVidenceModeler and the Program to Assemble Spliced Alignments. Genome Biol. 2008, 9, R7. [Google Scholar] [CrossRef] [Green Version]
- Lai, E.C.; Burks, C.; Posakony, J.W. The K box, a conserved 3′ UTR sequence motif, negatively regulates accumulation of enhancer of split complex transcripts. Development 1998, 125, 4077–4088. [Google Scholar] [CrossRef]
- Harris, J.C.; Hrmova, M.; Lopato, S.; Langridge, P. Modulation of plant growth by HD-Zip class I and II transcription factors in response to environmental stimuli. New Phytol. 2011, 190, 823–837. [Google Scholar] [CrossRef]
- Agalou, A.; Purwantomo, S.; Overnas, E.; Johannesson, H.; Zhu, X.; Estiati, A.; de Kam, R.J.; Engstrom, P.; Slamet-Loedin, I.H.; Zhu, Z.; et al. A genome-wide survey of HD-Zip genes in rice and analysis of drought-responsive family members. Plant Mol. Biol. 2008, 66, 87–103. [Google Scholar] [CrossRef]
- Samarskiĭ, A. Claverie JM: The significance of digital gene expression profiles. Genome Res. Dokl. Akad. Nauk. Sssr. 1997, 631–634. [Google Scholar]
- Ebrahimian-Motlagh, S.; Ribone, P.A.; Thirumalaikumar, V.P.; Allu, A.D.; Chan, R.L.; Mueller-Roeber, B.; Balazadeh, S. JUNGBRUNNEN1 Confers Drought Tolerance Downstream of the HD-Zip I Transcription Factor AtHB13. Front. Plant Sci. 2017, 8, 2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabello, J.V.; Giacomelli, J.I.; Piattoni, C.V.; Iglesias, A.A.; Chan, R.L. The sunflower transcription factor HaHB11 improves yield, biomass and tolerance to flooding in transgenic Arabidopsis plants. J. Biotechnol. 2016, 222, 73–83. [Google Scholar] [CrossRef]
- Manavella, P.A.; Dezar, C.A.; Bonaventure, G.; Baldwin, I.T.; Chan, R.L. HAHB4, a sunflower HD-Zip protein, integrates signals from the jasmonic acid and ethylene pathways during wounding and biotic stress responses. Plant J. Cell Mol. Biol. 2010, 56, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.K.; Yoon, J.; Choi, G.J.; Jang, H.A.; Kwon, S.Y.; Choi, D. Capsicum annuum homeobox 1 (CaHB1) is a nuclear factor that has roles in plant development, salt tolerance, and pathogen defense. Biochem. Biophys. Res. Commun. 2013, 442, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shen, F.; Xu, X.; Zheng, Q.; Wang, Y.; Wu, T.; Li, W.; Qiu, C.; Xu, X.; Han, Z.; et al. An HD-ZIP transcription factor, MxHB13, integrates auxin-regulated and juvenility-determined control of adventitious rooting in Malus xiaojinensis. Plant J. 2021, 107, 1663–1680. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F. Basic local alignment search tool (BLAST). J. Mol. Biol. 2012, 215, 403–410. [Google Scholar] [CrossRef]
- Sudhir, K.; Glen, S.; Koichiro, T. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, 1, 2–3. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Tae-Ho, L.; Jin, H.; Barry, M.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Rui, X.; Hao, C.; He, Y. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Dewey, C.N.; Li, B. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar]
- Likun; Wang; Zhixing; Feng; Xi; Wang; Xiaowo; Wang, DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [CrossRef]
- He, S.; Yuan, G.; Bian, S.; Han, X.; Zhang, C. Major Latex Protein MdMLP423 Negatively Regulates Defense against Fungal Infections in Apple. Int. J. Mol. Sci. 2020, 21, 1879. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data using Real-Time Quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID | Chromosome Location | Exon | Size (Amino Acids) | MW (kDa) | pI |
|---|---|---|---|---|---|---|
| MdHDZ1 | MD00G1036300 | Chr00:6430068..6432722 | 2 | 243 | 27.82 | 5.37 |
| MdHDZ2 | MD01G1036200 | Chr01:12167559..12171263 | 3 | 330 | 37.26 | 5.09 |
| MdHDZ3 | MD01G1069700 | Chr01:17372035..17373849 | 2 | 415 | 45.63 | 8.94 |
| MdHDZ4 | MD01G1226600 | Chr01:31679345..31681430 | 2 | 236 | 27.47 | 5.36 |
| MdHDZ5 | MD02G1192800 | Chr02:18264264..18266606 | 3 | 289 | 31.93 | 8.36 |
| MdHDZ6 | MD02G1216800 | Chr02:24770720..24774323 | 4 | 309 | 34.57 | 8.13 |
| MdHDZ7 | MD02G1318700 | Chr02:37304680..37307701 | 4 | 373 | 41.40 | 6.06 |
| MdHDZ8 | MD03G1118500 | Chr03:10745381..10758148 | 18 | 843 | 92.52 | 6.09 |
| MdHDZ9 | MD04G1061200 | Chr04:8078000..8080961 | 4 | 310 | 34.59 | 7.62 |
| MdHDZ10 | MD05G1273700 | Chr05:40852942..40861482 | 18 | 852 | 92.95 | 6.03 |
| MdHDZ11 | MD06G1032300 | Chr06:3869510..3870336 | 1 | 196 | 22.40 | 9.52 |
| MdHDZ12 | MD06G1054800 | Chr06:8046646..8049808 | 4 | 310 | 34.60 | 8.42 |
| MdHDZ13 | MD06G1187600 | Chr06:32505970..32509240 | 3 | 324 | 36.40 | 4.79 |
| MdHDZ14 | MD07G1002500 | Chr07:275492..278216 | 4 | 370 | 40.91 | 6.44 |
| MdHDZ15 | MD07G1156200 | Chr07:22690670..22692966 | 2 | 242 | 27.82 | 4.75 |
| MdHDZ16 | MD07G1297100 | Chr07:35700128..35702089 | 2 | 231 | 26.66 | 5.78 |
| MdHDZ17 | MD08G1075400 | Chr08:6123797..6126556 | 3 | 303 | 34.09 | 6.94 |
| MdHDZ18 | MD08G1075500 | Chr08:6137356..6139170 | 4 | 269 | 30.11 | 6.60 |
| MdHDZ19 | MD08G1112900 | Chr08:10008344..10017933 | 18 | 832 | 91.75 | 5.90 |
| MdHDZ20 | MD08G1188500 | Chr08:23799477..23801844 | 3 | 279 | 31.43 | 5.57 |
| MdHDZ21 | MD09G1035100 | Chr09:2143505..2148775 | 4 | 336 | 38.15 | 5.29 |
| MdHDZ22 | MD09G1049000 | Chr09:3226829..3230190 | 3 | 300 | 33.92 | 6.55 |
| MdHDZ23 | MD09G1205400 | Chr09:19576024..19585691 | 18 | 838 | 91.88 | 6.03 |
| MdHDZ24 | MD10G1253500 | Chr10:34599032..34608186 | 18 | 851 | 93.08 | 6.03 |
| MdHDZ25 | MD11G1136800 | Chr11:12604948..12613845 | 18 | 812 | 88.65 | 5.85 |
| MdHDZ26 | MD12G1055800 | Chr12:6297205..6300831 | 4 | 340 | 37.68 | 8.69 |
| MdHDZ27 | MD12G1100600 | Chr12:15668169..15670196 | 3 | 231 | 27.00 | 8.37 |
| MdHDZ28 | MD13G1025200 | Chr13:1775080..1778991 | 4 | 226 | 25.40 | 9.09 |
| MdHDZ29 | MD13G1030700 | Chr13:2201012..2203672 | 3 | 324 | 36.46 | 4.88 |
| MdHDZ30 | MD13G1074800 | Chr13:5277838..5280985 | 3 | 289 | 32.95 | 6.06 |
| MdHDZ31 | MD13G1079500 | Chr13:5590336..5593469 | 3 | 333 | 37.30 | 4.81 |
| MdHDZ32 | MD13G1196700 | Chr13:17146326..17146976 | 1 | 154 | 18.34 | 9.46 |
| MdHDZ33 | MD13G1236500 | Chr13:23981101..23982690 | 3 | 242 | 27.33 | 8.02 |
| MdHDZ34 | MD14G1056200 | Chr14:5791182..5794527 | 4 | 342 | 37.78 | 7.62 |
| MdHDZ35 | MD14G1056300 | Chr14:5813565..5815376 | 4 | 226 | 25.50 | 8.27 |
| MdHDZ36 | MD14G1094700 | Chr14:14187962..14190102 | 3 | 232 | 27.00 | 8.95 |
| MdHDZ37 | MD15G1062900 | Chr15:4328848..4331612 | 3 | 301 | 33.96 | 6.49 |
| MdHDZ38 | MD15G1092200 | Chr15:6404045..6413541 | 18 | 830 | 91.67 | 5.93 |
| MdHDZ39 | MD15G1302900 | Chr15:29349544..29351859 | 3 | 289 | 31.98 | 8.12 |
| MdHDZ40 | MD15G1319800 | Chr15:33163160..33166835 | 3 | 329 | 37.18 | 5.21 |
| MdHDZ41 | MD15G1374500 | Chr15:45816138..45818593 | 3 | 274 | 31.09 | 6.79 |
| MdHDZ42 | MD16G1027800 | Chr16:1954912..1960238 | 4 | 289 | 32.71 | 8.99 |
| MdHDZ43 | MD16G1076000 | Chr16:5322782..5325852 | 3 | 286 | 32.52 | 5.80 |
| MdHDZ44 | MD16G1079400 | Chr16:5571757..5575130 | 3 | 327 | 37.07 | 4.78 |
| MdHDZ45 | MD16G1241700 | Chr16:26132793..26134339 | 3 | 230 | 26.13 | 6.83 |
| MdHDZ46 | MD17G1035400 | Chr17:2532964..2538305 | 4 | 332 | 37.43 | 5.18 |
| MdHDZ47 | MD17G1049000 | Chr17:3572666..3576024 | 3 | 304 | 34.41 | 6.36 |
| MdHDZ48 | MD17G1185400 | Chr17:22000363..22009994 | 18 | 838 | 92.12 | 6.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Han, X.; Liang, Z.; Yan, J.; Cong, P.; Zhang, C. Genome-Wide Identification, Classification, and Expression Analysis of the HD-Zip Transcription Factor Family in Apple (Malus domestica Borkh.). Int. J. Mol. Sci. 2022, 23, 2632. https://doi.org/10.3390/ijms23052632
Liu K, Han X, Liang Z, Yan J, Cong P, Zhang C. Genome-Wide Identification, Classification, and Expression Analysis of the HD-Zip Transcription Factor Family in Apple (Malus domestica Borkh.). International Journal of Molecular Sciences. 2022; 23(5):2632. https://doi.org/10.3390/ijms23052632
Chicago/Turabian StyleLiu, Kai, Xiaolei Han, Zhaolin Liang, Jiadi Yan, Peihua Cong, and Caixia Zhang. 2022. "Genome-Wide Identification, Classification, and Expression Analysis of the HD-Zip Transcription Factor Family in Apple (Malus domestica Borkh.)" International Journal of Molecular Sciences 23, no. 5: 2632. https://doi.org/10.3390/ijms23052632
APA StyleLiu, K., Han, X., Liang, Z., Yan, J., Cong, P., & Zhang, C. (2022). Genome-Wide Identification, Classification, and Expression Analysis of the HD-Zip Transcription Factor Family in Apple (Malus domestica Borkh.). International Journal of Molecular Sciences, 23(5), 2632. https://doi.org/10.3390/ijms23052632