
Arrow of Time, Entropy, and Protein Folding: Holistic View on Biochirality



Abstract
:1. Introduction
2. Space-Time Symmetry and Entropy
3. Biological Language of Protein Conformation
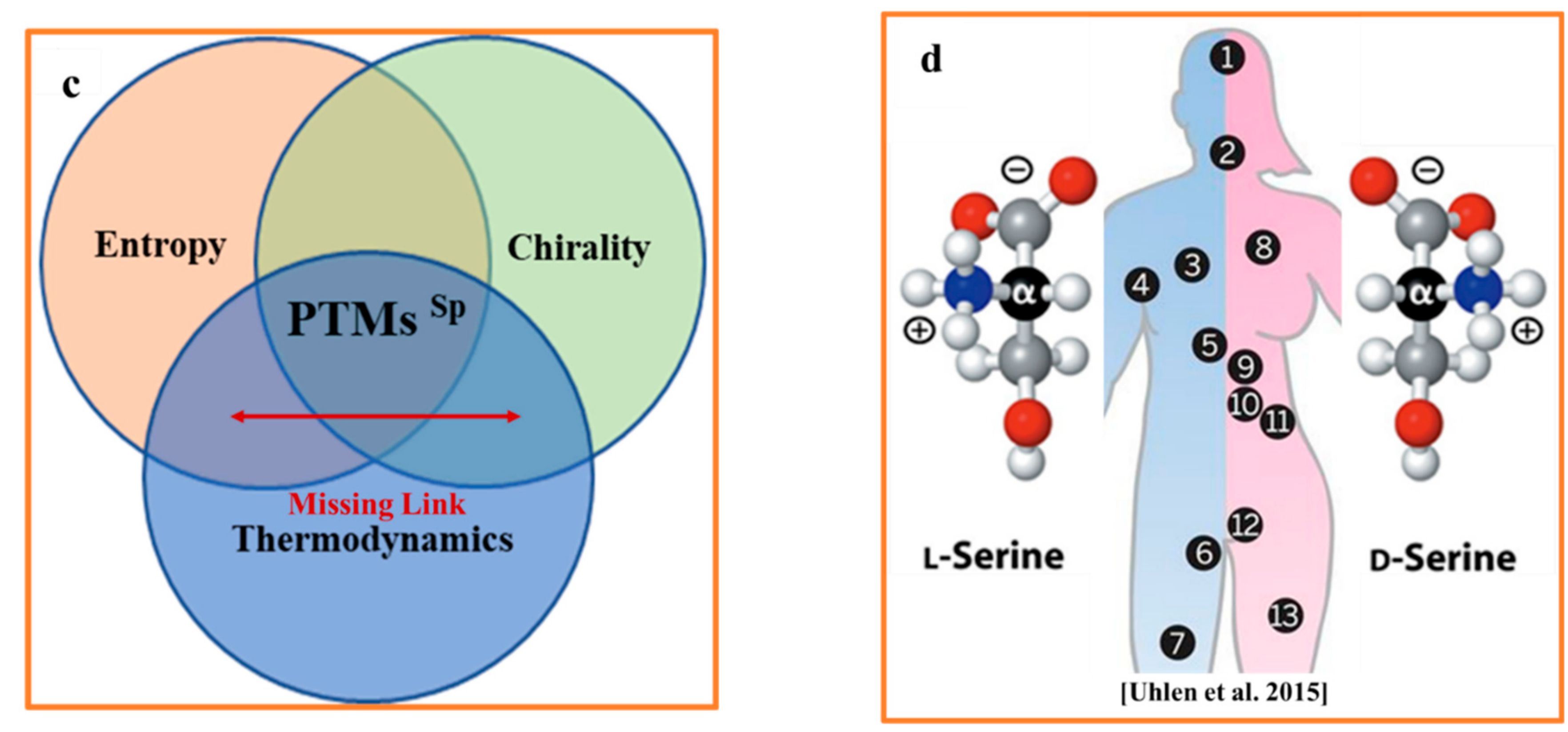
3.1. Protein Homochirality
3.2. Fundamental Roots of Homochirality
3.3. Spontaneous Racemization
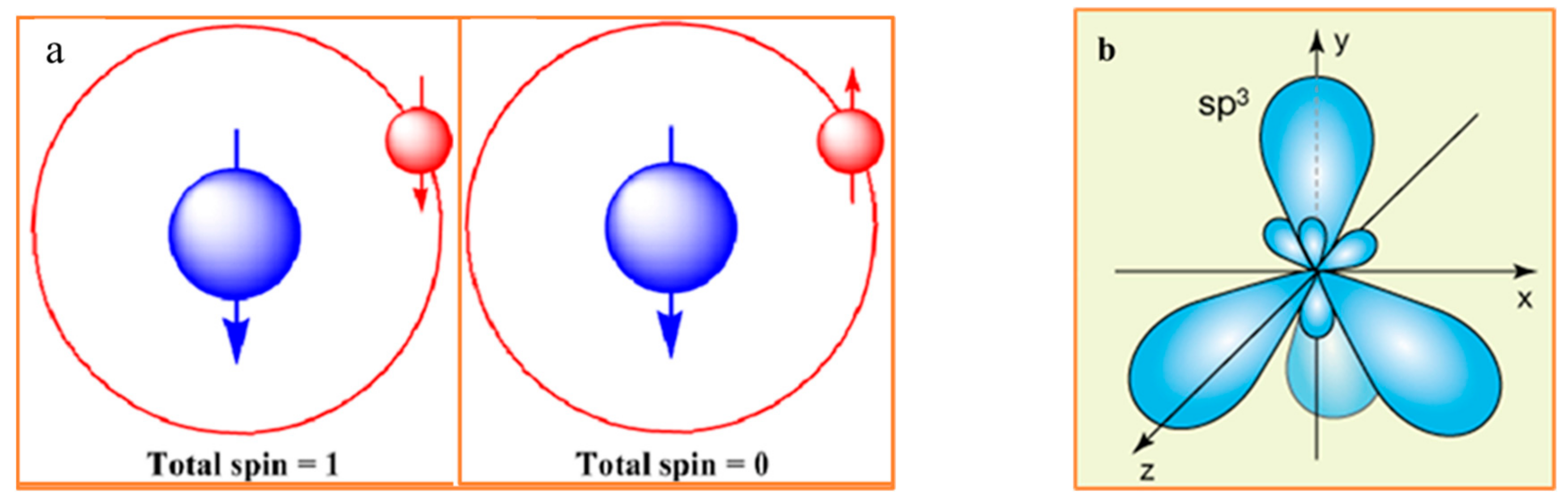
4. Entropy
5. Epilogue
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AAs | Amino acids |
| DNA | Deoxyribonucleic acid |
| GRNs | Gene regulatory networks |
| PTMsEnz | Enzymatic post translational modifications |
| L-AAs | L- amino acids |
| D-AAs | D-Amino acids |
| NS | Native state |
| NE | Non-equilibrium |
| PDB | Protein Data Bank |
| RNAs | Ribonucleic acids |
| PTMsSp | Spontaneous post-translational modifications |
| PhTsSp | Spontaneous phase transitions |
| Swiss-ProtFraction | Swiss-Prot Database |
References
- Jaakkola, S.; Sharma, V.; Annila, A. Cause of Chirality Consensus. Curr. Chem. Biol. 2008, 2, 153–158. [Google Scholar] [CrossRef]
- Cheng, L.; Liu, K.; Duan, Y.; Duan, H.; Li, Y.; Gao, M.; Cao, L. Adaptive Chirality of an Achiral Cage: Chirality Transfer, Induction, and Circularly Polarized Luminescence through Aqueous Host–Guest Complexation. CCS Chem. 2021, 3, 2749–2763. [Google Scholar] [CrossRef]
- Liu, X.; Lu, J.; Chen, J.; Zhang, M.; Chen, Y.; Xing, F.; Feng, L. Chiral Self-Assembly of Porphyrins Induced by Chiral Carbon Dots. Front. Chem. 2020, 8, 670. [Google Scholar] [CrossRef]
- Michiya Fujiki, M. Mirror symmetry breaking of silicon polymers—from weak bosons to artificial helix. Chem. Rec. 2009, 9, 271–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poppitz, E.; Shang, Y. Chiral Lattice Gauge Theories Via Mirror-Fermion Decoupling: A Mission (im)Possible? Int. J. Mod. Phys. A 2010, 25, 2761–2813. [Google Scholar] [CrossRef] [Green Version]
- Wang, W. Hypothesis. Electron Spin and the Origin of Bio-homochirality I. Extant Enzymatic Reaction Model. arXiv 2013, arXiv:1309.1229. [Google Scholar]
- Chi, P.B.; Liberles, D.A. Selection on protein structure, interaction, and sequence. Prot. Sci. 2016, 25, 1168–1178. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, R.A.; Pollock, D.D. Sequence entropy of folding and the absolute rate of amino acid substitutions. Nat. Ecol. Evol. 2017, 12, 1923–1930. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Cheng, K.; Mao, J.; Liu, F.; Liu, J.; Ye, M.; Zou, H. Quantitative proteomics reveals the kinetics of trypsin-catalyzed protein digestion. Anal. Bioanal. Chem. 2014, 406, 6247–6256. [Google Scholar] [CrossRef]
- Shen, S.; Kai, B.; Ruan, J.; Huzil, J.T.; Carpenter, E.; Tuszynski, J.A. Probabilistic analysis of the frequencies of amino acid pairs within characterized protein sequences. Physica A 2006, 370, 651–662. [Google Scholar] [CrossRef]
- Guo, A.-M.; Sun, Q.-F. Spin-dependent electron transport in protein-like single-helical molecules. Proc. Natl. Acad. Sci. USA 2014, 111, 11658–11662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaeli, K.; Kantor-Uriel, N.; Naaman, R.; Waldeck, D.H. The Electron’s Spin and Molecular Chirality-How are they Related and How do they Affect Life Processes? Chem Soc. Rev. 2016, 45, 6478–6487. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Mishra, S.; Avigad, E.; Bloom, B.P.; Baczewski, L.T.; Yochelis, S.; Paltiel, Y.; Naaman, R.; Waldeck, D.H. Effect of Chiral Molecules on the Electron’s Spin Wavefunction at Interfaces. J. Phys. Chem. Lett. 2020, 11, 1550–1557. [Google Scholar] [CrossRef]
- Schaible, M.J.; Rosenberg, R.A.; Kundu, S.; Orlando, T.M. Electron Spin-Polarization Dependent Damage to Chiral Amino Acid L-Histidine. J. Phys. Chem. Lett. 2020, 11, 10182–10187. [Google Scholar] [CrossRef] [PubMed]
- Almheiri, A.; Hartman, T.; Maldacena, J.; Shaghoulian, S.; Tajdini, A. The entropy of Hawking radiation. Rev. Mod. Phys. 2021, 93, 35002. [Google Scholar] [CrossRef]
- He, D.; Cai, Q. Area Entropy and Quantized Mass of Black Holes from Information Theory. Entropy 2021, 23, 858. [Google Scholar] [CrossRef]
- Poulikakos, L.V.; Dionne, J.A.; García-Etxarri, A. Perspective. Optical Helicity and Optical Chirality in Free Space and in the Presence of Matter. Symmetry 2019, 11, 1113. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Zhou, Y.; Chen, Y. Stimulated chiral light-matter interactions in biological micro-lasers. ACS Nano 2021, 15, 8965–8975. [Google Scholar] [CrossRef]
- Pritišanac, I.; Vernon, R.M.; Moses, A.M.; Forman Kay, J.D. Entropy and Information within Intrinsically Disordered Protein Regions. Entropy 2019, 21, 662. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, K.J.; Rovelli, C. Transitions in Brain Evolution: Space, Time and Entropy. Trends Neurosci. 2020, 43, 467–474. [Google Scholar] [CrossRef]
- Rongjia Yang, R. Is Gravity Entropic Force? Entropy 2014, 16, 4483–4488. [Google Scholar] [CrossRef] [Green Version]
- García-Ariza, M.A.; Montesinos, M.; del Castillo, G.F.T. Geometric Thermodynamics: Black Holes and the Meaning of the Scalar Curvature. Entropy 2014, 16, 6515–6523. [Google Scholar] [CrossRef] [Green Version]
- Meyers, R.E.; Keith, S.; Deacon, K.S. Space-Time Quantum Imaging. Entropy 2015, 17, 1508–1534. [Google Scholar] [CrossRef] [Green Version]
- Mathur, A.D. Losing Information Outside the Horizon. Entropy 2015, 17, 4083–4109. [Google Scholar] [CrossRef] [Green Version]
- Schrödinger, E. What is Life? In The Physical Aspect of the Living Cell; Cambridge University Press: Cambridge, UK, 1944; ISBN 978-0-521-42708-1. [Google Scholar]
- Hayflick, L. Entropy Explains Aging, Genetic Determinism Explains Longevity, and Undefined Terminology Explains Misunderstanding Both. PLoS Genet. 2007, 3, e220. [Google Scholar] [CrossRef] [PubMed]
- Gołos, J. Entropy and the Direction of Time. Entropy 2021, 23, 388. [Google Scholar] [CrossRef] [PubMed]
- Dyakin, V.V.; Wisniewski, T.M.; Lajtha, A. Racemization in Post-Translational Modifications Relevance to Protein Aging, Aggregation and Neurodegeneration: Tip of the Iceberg. Symmetry 2021, 13, 455. [Google Scholar] [CrossRef] [PubMed]
- Carhart-Harris, R.L.; Leech, R.; Hellyer, P.J.; Shanahan, M.; Feilding, A.; Tagliazucchi, E. The entropic brain: A theory of conscious states informed by neuroimaging research with psychedelic drugs. Front. Hum. Neurosci. 2014, 8, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carhart-Harris, R.L. The entropic brain-revisited. Neuropharmacology 2018, 142, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Cieri, F.; Zhuang, X.; Caldwell, J.Z.K.; Cordes, D. Hypothesis and Theory Article. Brain Entropy During Aging Through a Free Energy Principle Approach. Front. Hum. Neurosci. 2021, 15, 139. [Google Scholar] [CrossRef]
- Zivieri, R.; Pacini, N.; Finocchio, G. Rate of entropy model for irreversible processes in living systems. Sci. Rep. 2017, 7, 9134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondepudi, D.; Kapcha, L. Entropy production in chiral symmetry breaking transitions. Chirality 2008, 20, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.C.; Rieper, E.; Tuszynski, J.A. Self-organization and entropy reduction in a living cell. Bio. Syst. 2013, 111, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyakin, V.V.; Lucas, J.; Dyakina-Fagnano, N.V.; Posner, E.V.; Vadasz, C. The Chain of Chirality Transfer as Determinant of Brain Functional Laterality. Breaking the Chirality Silence: Search for New Generation of Biomarkers; Relevance to Neurodegenerative Diseases, Cognitive Psychology, and Nutrition Science. Neurol. Neurosci. Res. 2017, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Holló, G. Demystification of animal symmetry: Symmetry is a response to mechanical forces. Biol. Direct. 2017, 12, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamads, A. Chiral Neuronal Motility: The Missing Link between Molecular Chirality and Brain Asymmetry. Symmetry 2019, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Laurent, G.; Lacoste, D.; Gaspard, P. Emergence of homochirality in large molecular systems. Proc. Natl. Acad. Sci. USA 2021, 118, e2012741118. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Yaku, K.; Nakagawa, T. Simultaneous Measurement of Amino Acid Enantiomers in Aged Mouse Brain Samples by LC/MS/MS Combined with Derivatization Using Nα-(5-Fluoro-2,4-dinitrophenyl)-l-leucinamide (l-FDLA). Metabolites 2021, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- MacNeil, L.T.; Walhout, A.J.M. Gene regulatory networks and the role of robustness and stochasticity in the control of gene expression. Genome Res. 2011, 21, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, I.R. The evolution of universal adaptations of life is driven by universal properties of matter: Energy, entropy, and interaction. F1000Research 2020, 18, 626. [Google Scholar] [CrossRef]
- Ayon, N.J. Features, roles and chiral analyses of proteinogenic amino acids. AIMS Mol. Sci. 2020, 7, 229–268. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Bauer, D.; Hamacher, K.; Bröer, S.; Pauleit, D.; Palm, C.; Zilles, K.; Coenen, H.H.; Langen, K.-J. Preferred stereoselective brain uptake of d-serine--a modulator of glutamatergic neurotransmission Comparative Study. Nucl. Med. Biol. 2005, 32, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Novotný, M.; Kleywegt, G.J. A Survey of Left-handed Helices in Protein Structures. J. Mol. Biol. 2005, 347, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, N.E.; Hoang, H.N.; Abbenante, G.; Fairlie, D.P. Left- and right-handed alpha-helical turns in homo- and 03-hetero-chiral helical scaffolds. J. Am. Chem. Soc. 2009, 131, 15877–15886. [Google Scholar] [CrossRef] [PubMed]
- Jacco, J.A.J.; Bastings, J.A.J.; Hans, M.; van Damink, E.S.W.O.; Rensen, S.S. D-amino Acids in Health and Disease: A Focus on Cancer. Nutrients 2019, 11, 2205. [Google Scholar] [CrossRef] [Green Version]
- Ebeling, W.; Volkenstein, M.V. Entropy and the evolution of biological information. Physica A 1990, 63, 398–402. [Google Scholar] [CrossRef]
- Bishop, R.C. Nonequilibrium statistical mechanics Brussels-Austinstyle. Stud. Hist. Philos. Mod. Phys. 2004, 35, 1–30. [Google Scholar] [CrossRef]
- Berry, S. Entropy irreversibility and evolution. J. Theor. Biol. 1995, 175, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Mangel, M.; Kindsvater, H.K.; Bon-sall, M.B. Evolutionary analysis of life span, competition, and adaptive radiation, motivated by the pacific rockfishes (sebastes). Evolution 2007, 61, 1208–1224. [Google Scholar] [CrossRef]
- Geiger, T.; Clarke, S. Deamidation, isomerization, and racemization at asparaginyl and aspartyl residues in peptides. Succinimide-linked reactions that contribute to protein degradation. J. Biol Chem. 1987, 262, 785–794. [Google Scholar] [CrossRef]
- McCudden, C.R.; Kraus, V.B. Biochemistry of amino acid racemization and clinical application to musculoskeletal disease. Clin. Biochem. 2006, 39, 1112–1130. [Google Scholar] [CrossRef]
- Gorisse, L.; Pietrement, C.; Vuiblet, V.; Schmelzer, C.E.H.; Köhler, M.; Duca, L.; Debelle, L.; Fornès, P.; Jaisson, S.; Gillery, F. Protein carbamylation is a hallmark of aging. Proc. Natl. Acad. Sci. USA 2016, 113, 1191–1196. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, C.; Jaisson, S.; Gorisse, L. Carbamylation and glycation compete for collagen molecular aging in vivo. Sci. Rep. 2019, 9, 18291. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, R.; Defelipe, L.A.; Aliperti, L.; Niebling, S.; Custódio, T.F.; Löw, C.; Schwarz, J.J.; Remans, K.; Craig, P.O.; Otero, L.H.; et al. Deamidation drives molecular aging of the SARS-CoV-2 spike protein receptor-binding motif. J. Biol. Chem. 2021, 297, 101175. [Google Scholar] [CrossRef] [PubMed]
- Dyakin, V.V.; Dyakina-Fagnano, N.V.; Mcintire, L.B.; Uversky, V.N. Fundamental Clock of Biological Aging: Convergence of Molecular, Neurodegenerative, Cognitive and Psychiatric Pathways: Non-Equilibrium Thermodynamics Meet Psychology. Int. J. Mol. Sci. 2022, 23, 285. [Google Scholar] [CrossRef] [PubMed]
- Toyama, B.H.; Savas, J.N.; Park, S.K.; Harris, M.S.; Ingolia, N.T.; Yates, J.R.; Hetzer, M.W. Identification of long-lived proteins reveals exceptional stability of essential cellular structures. Cell 2013, 54, 971–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.; He, J.; Zou, J. Phosphorylated lipid-conjugated oligonucleotide selectively anchors on cell membranes with high alkaline phosphatase expression. Nat. Commun. 2019, 10, 2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Ma, W. The origin of biological homochirality along with the origin of life. PLoS Comput. Biol. 2020, 16, e1007592. [Google Scholar] [CrossRef]
- Calvo, O.; García, A. RNA Polymerase II Phosphorylation and Gene Expression Regulation. In Protein Phosphorylation in Human Health; InTech Open: Rijeka, Croatia, 2012; Chapter 5; pp. 151–194. [Google Scholar] [CrossRef] [Green Version]
- Bada, J.L.; Cleaves, H.J. Amino Acid Racemization and Homochirality on Earth and Elsewhere. In Proceedings of the Astrobiology Science Conference 2015, Chicago, IL, USA, 15–19 June 2015. [Google Scholar]
- Malyshko, E.V.; Semenova, E.V.; Bagrova, O.E.; Murtazina, A.R.; Tverdislov, V.A. Chiral Dualism as a Unifying Principle in Molecular Biophysics. Biophysica 2021, 1, 3. [Google Scholar] [CrossRef]
- Aberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2003. [Google Scholar]
- Peter, G. Vekilova. Phase transitions of folded proteins. Soft Matter 2010, 21, 5254–5272. [Google Scholar]
- Uversky, V.N.; Finkelstein, A.V. Life in Phases: Intra-and Inter-Molecular Phase. Transitions in Protein Solutions. Biomolecules 2019, 9, 842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuruyama, T. Non-equilibrium thermodynamics of biological signal transduction predicts conservation of entropy production rate. J. Theor. Biol. 2019, 472, 84–87. [Google Scholar] [CrossRef]
- Jacobson, S.J.; Willson, C.G.; Rapoport, H. Mechanism of cystine racemization in strong acid. J. Org. Chem. 1974, 39, 1074–1077. [Google Scholar] [CrossRef] [PubMed]
- Kasai, A.; Yamashita, N.; Utsunomiya-Tate, N. Collagen Racemization and Deposition in the Lungs of Aged Rats. Biochem. Insights 2010, 3, 25–33. [Google Scholar] [CrossRef]
- Dyakin, V.V.; Lajtha, A.; Dyakina-Fagnano, N.V. Racemization hypothesis of neurodegeneration (RHND). Alsheimer’s Denentia. Suppl. Basic Sci. Pathog. Part 2 2020, 16, e047697. [Google Scholar] [CrossRef]
- Jayant, B.; Ungaonkar, J.B. Entropy in Biology. Resonance 2001, 6, 61–66. [Google Scholar] [CrossRef]
- Binyamin, I.; Meidan-Shani, S.; Ashkenaz, N. Synthesis of γ-hydroxypropyl P-chirogenic (±)-phosphorus oxide derivatives by regioselective ring-opening of oxaphospholane 2-oxide precursors. Beilstein J. Org. Chem. 2015, 11, 1332–1339. [Google Scholar] [CrossRef] [Green Version]
- Dutartre, M.; Bayardon, J.; Juge, S. Applications and stereoselective syntheses of P-chirogenic phosphorus compounds. Chem. Soc. Rev. 2016, 45, 5771–5794. [Google Scholar] [CrossRef]
- Krasowska, D.; Chrzanowski, J.; Kiełbasiński, P.; Drabowicz, J. Chiral Hypervalent, Pentacoordinated Phosphoranes. Molecules 2016, 21, 1573. [Google Scholar] [CrossRef]
- Mondal, A.; Thiel, N.O.; Dorel, R. P-chirogenic phosphorus compounds by stereoselective Pd-catalysed arylation of phosphoramidites. Nat. Catal. 2022, 5, 10–19. [Google Scholar] [CrossRef]
- Biswas, S.; Kinbara, K.; Niwa, T.; Taguchi, H.; Ishii, N.; Watanabe, S.; Miyata, K.; Kataoka, K.; Takuzo Aida, T. Biomolecular robotics for chemomechanically driven guest delivery fuelled by intracellular ATP. Nat. Chem. 2013, 5, 613–620. [Google Scholar] [CrossRef]
- Kumar, M.; Brocorens, P.; Tonnele, C.; Beljonne, D.; Surin, M.; George, S.J. A dynamic supramolecular polymer with stimuli-responsive handedness for in situ probing of enzymatic ATP hydrolysis. Nat. Commun. 2014, 5, 5793. [Google Scholar] [CrossRef] [PubMed]
- Termonia, Y. Entropy-driven self-assembly of nanoparticles into strings. Colloids Surf. A Physicochem. Eng. Asp. 2014, 447, 23–27. [Google Scholar] [CrossRef]
- Baxa, M.C.; Haddadian, E.J.; Jumper, J.M.; Freed, K.F.; Sosnick, T.R. Loss of conformational entropy in protein folding calculated using realistic ensembles and its implications for NMR-based calculations. Proc. Natl. Acad. Sci. USA 2014, 111, 15396–15401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frausto-Solis, J.; Liñán-García, E.; Sánchez-Hernández, J.P.; González-Barbosa, J.J.; González-Flores, C.; Castilla-Valdez, G. Multiphase Simulated Annealing Based on Boltzmann and Bose-Einstein Distribution Applied to Protein Folding Problem. Adv. Bioinform. 2016, 2016, 7357123. [Google Scholar] [CrossRef] [Green Version]
- Kadler, K.E.; Prockop, Y.H.D.J. Assembly of collagen fibrils de novo by cleavage of the type I pC-collagen with procollagen C-proteinase. Assay of critical concentration demonstrates that collagen self-assembly is a classic example of an entropy-driven process. J. Biol. Chem. 1987, 262, 15696–15701. [Google Scholar] [CrossRef]
- Otosu, T.; Ishii, K.; Oikawa, H.; Ara, M.; Takahashi, S.; Tahara, T. Highly Heterogeneous Nature of the Native and Unfolded States of the B Domain of Protein A Revealed by Two-Dimensional Fluorescence Lifetime Correlation Spectroscopy. J. Phys. Chem. B 2017, 121, 5463–5473. [Google Scholar] [CrossRef]
- Stollar, E.J.; Smith, D.P. Uncovering protein structure. Essays Biochem. 2020, 64, 649–680. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhang, K.; He, W.; Du, Y.; Hooi, M.; Lu, Y. Racemization at the Asp 58 residue in αA-crystallin from the lens of high myopic cataract patients. J. Cell Mol. Med. 2018, 22, 1118–1126. [Google Scholar] [CrossRef] [Green Version]
- Razavi, M.; Fathi, S.M.S.; Tuszynski, J.A. The Effect of the Protein Synthesis Entropy Reduction on the Cell Size Regulation and Division Size of Unicellular Organisms. Entropy 2022, 24, 94. [Google Scholar] [CrossRef] [PubMed]
- Demirel, Y. None-quilibrium Thermodynamics Modeling of Coupled Biochemical Cycles in Living Cells. J. Non Newton. Fluid Mech. 2010, 165, 953–972. [Google Scholar] [CrossRef] [Green Version]
- Towse, C.-L.; Akke, M.; Valerie Daggett, V. The Dynameomics Entropy Dictionary: A Large-Scale Assessment of Conformational Entropy Across Protein Fold Space. J. Phys. Chemistry. J. Phys. Chem. B 2017, 121, 3933–3945. [Google Scholar] [CrossRef] [PubMed]
- Bada, J.L. Amino Acid Racemization Dating of Fossil Bones. Annu. Rev. Earth Planet. Sci. 1985, 13, 241–268. [Google Scholar] [CrossRef]
- Balchin, D.; Hayer-Hartl, M.; Hartl, F.U. Recent advances in understanding catalysis of protein folding by molecular chaperones. FEBS Lett. 2020, 594, 2770–2781. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T. Conversion of psychological stress into cellular stress response: Roles of the sigma-1 receptor in the process PCN. Psychiatry Clin. Neuroscience. Front. Rev. 2015, 69, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Tsankova, T.; Renthal, W.; Kumar, A.; Eric, J.; Nestler, E.J. Epigenetic regulation in psychiatric disorders. Nat. Rev. Neurosci 2007, 8, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Zannas, A.S. Editorial Perspective: Psychological stress and epigenetic aging–what can we learn and how can we prevent? J. Child Psychol. Psychiatry 2016, 57, 67–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferracioli, N.G.M. Psychological aspects of aging and psychology’s contributions to gerontology: Theoretical and technical interface. MOJ Gerontol. Ger. 2018, 3, 115–116. [Google Scholar] [CrossRef] [Green Version]
- Schiele, M.A.; Gottschalk, M.G.; Domschke, K. The applied implications of epigenetics in anxiety, affective and stress-related disorders-A review and synthesis on psychosocial stress, psychotherapy and prevention. Clin. Psychol. Rev. 2020, 77, 101830. [Google Scholar] [CrossRef]
- Englander, M.T.; Avins, J.L.; Fleisher, R.C.; Liu, B.; Effraim, F.R.; Wang, J.; Schulten, K.; Leyh, T.S.; Gonzalez, R.L., Jr.; Cornish, V.W. The ribosome can discriminate the chirality of amino acids within its peptidyl-transferase center. Proc. Natl. Acad. Sci. USA 2015, 112, 6038–6043. [Google Scholar] [CrossRef] [Green Version]
- Nanda, V.; Andrianarijaona, A.; Narayan, C. The role of protein homochirality in shaping the energy landscape of folding. Protein Sci. 2007, 16, 1667–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Ramakrishnan, V.; Ranbhor, R.; Patel, K.; Durani, S. Homochiral Stereochemistry: The Missing Link of Structure to Energetics in Protein. Folding. J. Phys. Chem. B 2009, 113, 16435–16442. [Google Scholar] [CrossRef] [PubMed]
- Brady, G.P.; Sharp, K.A. Entropy in protein folding and in protein–protein interactions. Curr. Opin. Struct. Biol. 1997, 7, 215–221. [Google Scholar] [CrossRef]
- Parpura, V.; Fisher, E.S.; Lechleiter, J.D.; Schousboe, A.; Waagepetersen, H.S.; Brunet, S.; Baltan, S.; Verkhratsky, A. Glutamate and ATP at the Interface Between Signaling and Metabolism in Astroglia: Examples from Pathology. Neurochem. Res. 2017, 42, 19–34. [Google Scholar] [CrossRef]
- Uversky, V.N. Amyloidogenesis of Natively Unfolded Proteins. Curr. Alzheimer Res. 2008, 5, 260–287. [Google Scholar] [CrossRef] [PubMed]
- Raskatov, J.A.; Teplow, D.B. Using chirality to probe the conformational dynamics and assembly of intrinsically disordered amyloid proteins. Sci. Rep. 2017, 7, 12433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barykin, E.P.; Mitkevich, V.A.; Kozin, S.A.; Makarov, A.A. Amyloid β Modification: A Key to the Sporadic Alzheimer’s Disease? Front. Genet. 2017, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noether, E. Invariante Variationsprobleme. Nachrichten von der Gesellschaft der Wissenschaften zu Göttingen. Math. Phys. Klasse. 1918, 2, 235–257. [Google Scholar]
- Petitjean, M. About Chirality in Minkowski Spacetime. Symmetry 2019, 11, 1320. [Google Scholar] [CrossRef] [Green Version]
- Rocha, B.C.; Paul, S.; Vashisth, H. Role of Entropy in Colloidal Self-Assembly. Entropy 2020, 22, 877. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, H. Chemical reactions and the principle of maximal entropy production. J. Appl. Math. Phys. 1983, 34, 832–844. [Google Scholar] [CrossRef]
- Kleidon, A.; Malhi, Y.; Cox, P.M. Maximum entropy production in environmental and ecological systems. Philos. Trans. R Soc. Lond. B Biol. Sci. 2010, 365, 1297–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juretić, D.; Simunić, J.; Lošić, Z.B. Maximum Entropy Production Theorem for Transitions between Enzyme Functional States and Its Applications. Entropy 2019, 21, 743. [Google Scholar] [CrossRef] [Green Version]
- Toto, A.; Malagrinò, F.; Visconti, L.; Troilo, F.; Pagano, L.; Brunori, M.; Jemth, P.; Gianni, G. Reviews Templated Folding of Intrinsically Disordered Proteins. J. Biol. Chem. 2020, 295, P6586–P6593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, M.; Qin, W. Organic Chiral Spin-Optics: The Interaction between Spin and Photon in Organic Chiral Materials. Adv. Opt. Mater. 2021, 9, 2101201. [Google Scholar] [CrossRef]
- McManus, J.J.; Charbonneau, P.; Zaccarelli, E.; Asherie, N. The Physics of Protein Self-Assembly. Curr. Opin. Colloid Interface Sci. 2016, 22, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Nash, T. The Role of Entropy in Molecular Self-Assembly. J. Nanomed. Res. 2017, 5, 00126. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Alberstein, R.G.; De Yoreo, J.J. Assembly of a patchy protein into variable 2D lattices via tunable multiscale interactions. Nat. Commun. 2020, 11, 3770. [Google Scholar] [CrossRef] [PubMed]
- Liardon, R.; Hurrell, R.F. Amino acid racemization in heated and alkali-treated proteins. J. Agric. Food Chem. 1984, 31, 432–437. [Google Scholar] [CrossRef]
- Martinez, J.P.; De Oliveira, M.; Guiqin Cai, G.; Nachtschatt, M.; Navone, L.; Zhang, Z.; Robins, K.; Speight, R. Challenges and Opportunities in Identifying and Characterising Keratinases for Value-Added Peptide Production. Catalysts 2020, 10, 184. [Google Scholar] [CrossRef] [Green Version]
- Hameroff, S. Consciousness, the brain, and spacetime geometry. Ann. N. Y. Acad. Sci. 2001, 929, 74–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Song, Y.; Wang, W.; Tan, T.; Ji, Y.; Zhang, G.; Hu, J.; Zhang, Y. Macrochirality of Self-Assembled and Co-assembled Supramolecular Structures of a Pair of Enantiomeric Peptides. Front. Mol. Biosci. 2021, 8, 608. [Google Scholar] [CrossRef]
- Yu, J.; Yan, B.; Liu, C.-X. Model Hamiltonian and time reversal breaking topological phases of antiferromagnetic half-Heusler materials. Phys. Rev. B 2017, 95, 235158. [Google Scholar] [CrossRef] [Green Version]
- Watson, T.W.; Musielak, Z.E. Chiral Dirac Equation and Its Spacetime and CPT Symmetries. arXiv 2021, arXiv:2109.11015. [Google Scholar] [CrossRef]
- Grishin, D.V.; Zhdanov, D.D.; Pokrovskaya, M.V.; Sokolov, N.N. D-amino acids in nature, agriculture and biomedicine. All Life 2020, 13, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Seely, A.J.E. Optimizing Our Patients’ Entropy Production as Therapy? Hypotheses Originating from the Physics of Physiology. Entropy 2020, 22, 1095. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Proteino-Genic Amino Acids (AAs) | |||||||
|---|---|---|---|---|---|---|---|
| Amino Acid | H(1) | C(6) | N(7) | O(8) | S(16) | Swiss-PrtFr | |
| 1 | Alanine (Ala/A) | 7 | 3 | 1 | 2 | 0 | 0.0777 |
| 2 | Arginine (Arg/R) | 15 | 6 | 4 | 2 | 0 | 0.0526 |
| 3 | Asparagine (Asp/N) | 8 | 4 | 2 | 3 | 0 | 0.0437 |
| 4 | Aspartate (Asp/D) | 6 | 4 | 1 | 4 | 0 | 0.053 |
| 5 | Cysteine (Cys/C) | 7 | 3 | 1 | 2 | 1 | 0.0157 |
| 6 | Glutamate (Glu/E) | 8 | 5 | 1 | 4 | 0 | 0.0692 |
| 7 | Glutamine (Glx/Z) | 10 | 5 | 2 | 3 | 0 | 0.0532 |
| 8 | Glycine (Gly/G) | 5 | 2 | 1 | 2 | 0 | 0.0691 |
| 9 | Histidine (His/H) | 10 | 6 | 3 | 2 | 0 | 0.0227 |
| 10 | Isoleucine (Ile/I) | 13 | 6 | 1 | 2 | 0 | 0.0591 |
| 11 | Leucine (Leu/L) | 13 | 6 | 1 | 2 | 0 | 0.096 |
| 12 | Lysine (Lys/K) | 15 | 6 | 2 | 2 | 0 | 0.0595 |
| 13 | Methionine (Me/Mt) | 11 | 5 | 1 | 2 | 1 | 0.0238 |
| 14 | Phenylaninr (Phe/F) | 11 | 9 | 1 | 2 | 0 | 0.0405 |
| 15 | Proline (Pro/P) | 10 | 5 | 1 | 2 | 0 | 0.0469 |
| 16 | Serine (Ser/S) | 7 | 3 | 1 | 3 | 0 | 0.0694 |
| 17 | Threonine (Thr/T) | 9 | 4 | 1 | 3 | 0 | 0.055 |
| 18 | Tryptoohan(Trp/W) | 11 | 11 | 2 | 2 | 0 | 0.0118 |
| 19 | Tyrosin (Tyr/Y) | 11 | 9 | 1 | 3 | 0 | 0.0311 |
| 20 | Valine (Val/V) | 11 | 5 | 1 | 2 | 0 | 0.0667 |
| Non-Proteino-genic amino acids (AAs) | |||||||
| 1 | L-Carnitine | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dyakin, V.V.; Uversky, V.N. Arrow of Time, Entropy, and Protein Folding: Holistic View on Biochirality. Int. J. Mol. Sci. 2022, 23, 3687. https://doi.org/10.3390/ijms23073687
Dyakin VV, Uversky VN. Arrow of Time, Entropy, and Protein Folding: Holistic View on Biochirality. International Journal of Molecular Sciences. 2022; 23(7):3687. https://doi.org/10.3390/ijms23073687
Chicago/Turabian StyleDyakin, Victor V., and Vladimir N. Uversky. 2022. "Arrow of Time, Entropy, and Protein Folding: Holistic View on Biochirality" International Journal of Molecular Sciences 23, no. 7: 3687. https://doi.org/10.3390/ijms23073687
APA StyleDyakin, V. V., & Uversky, V. N. (2022). Arrow of Time, Entropy, and Protein Folding: Holistic View on Biochirality. International Journal of Molecular Sciences, 23(7), 3687. https://doi.org/10.3390/ijms23073687