
ABA Mediates Plant Development and Abiotic Stress via Alternative Splicing


Abstract
:1. Introduction
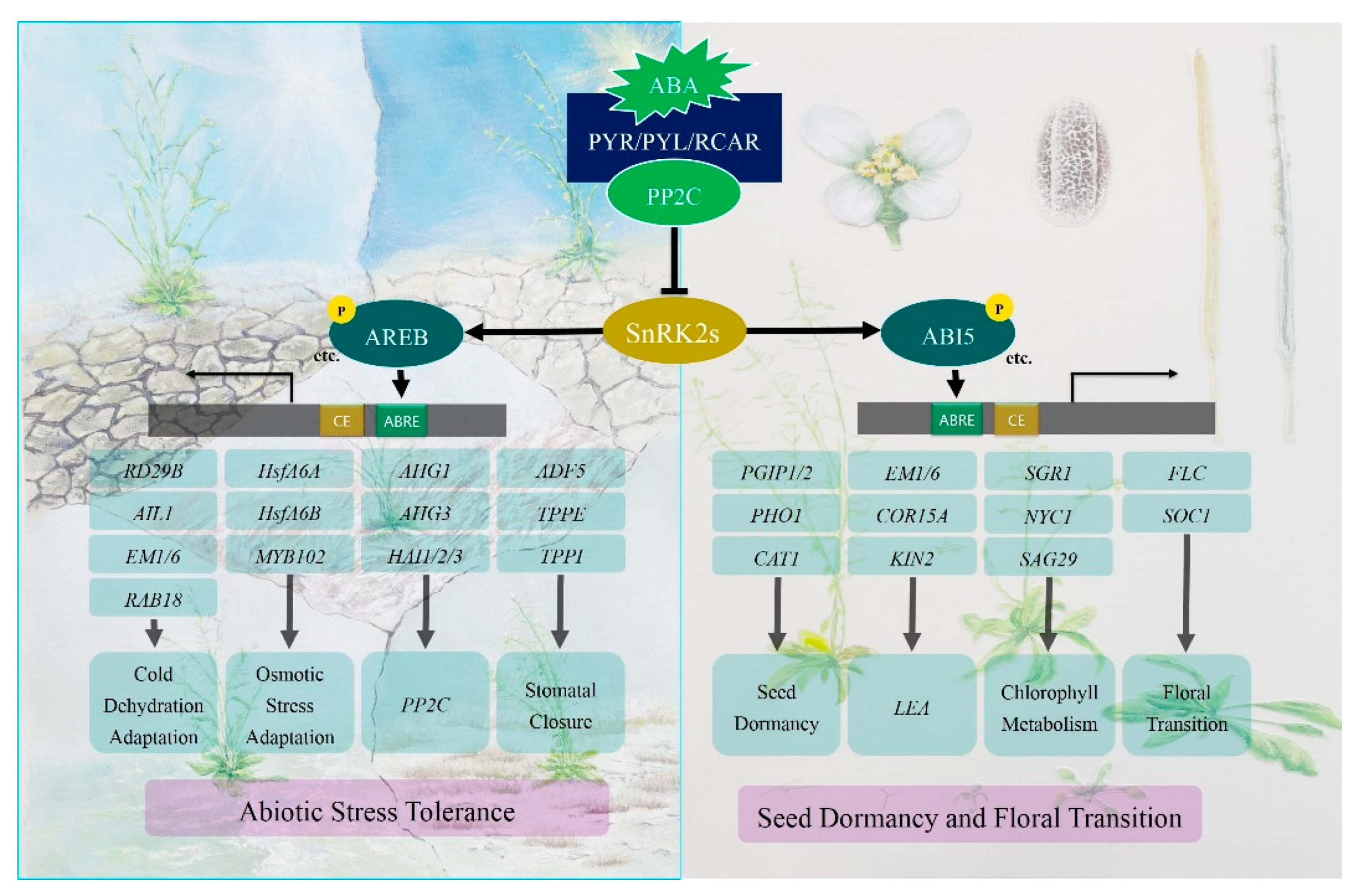
2. ABA Mediates Plant Development and Abiotic Stress
2.1. ABA in Responses to Abiotic Stress
2.2. ABA Mediates Plant Development
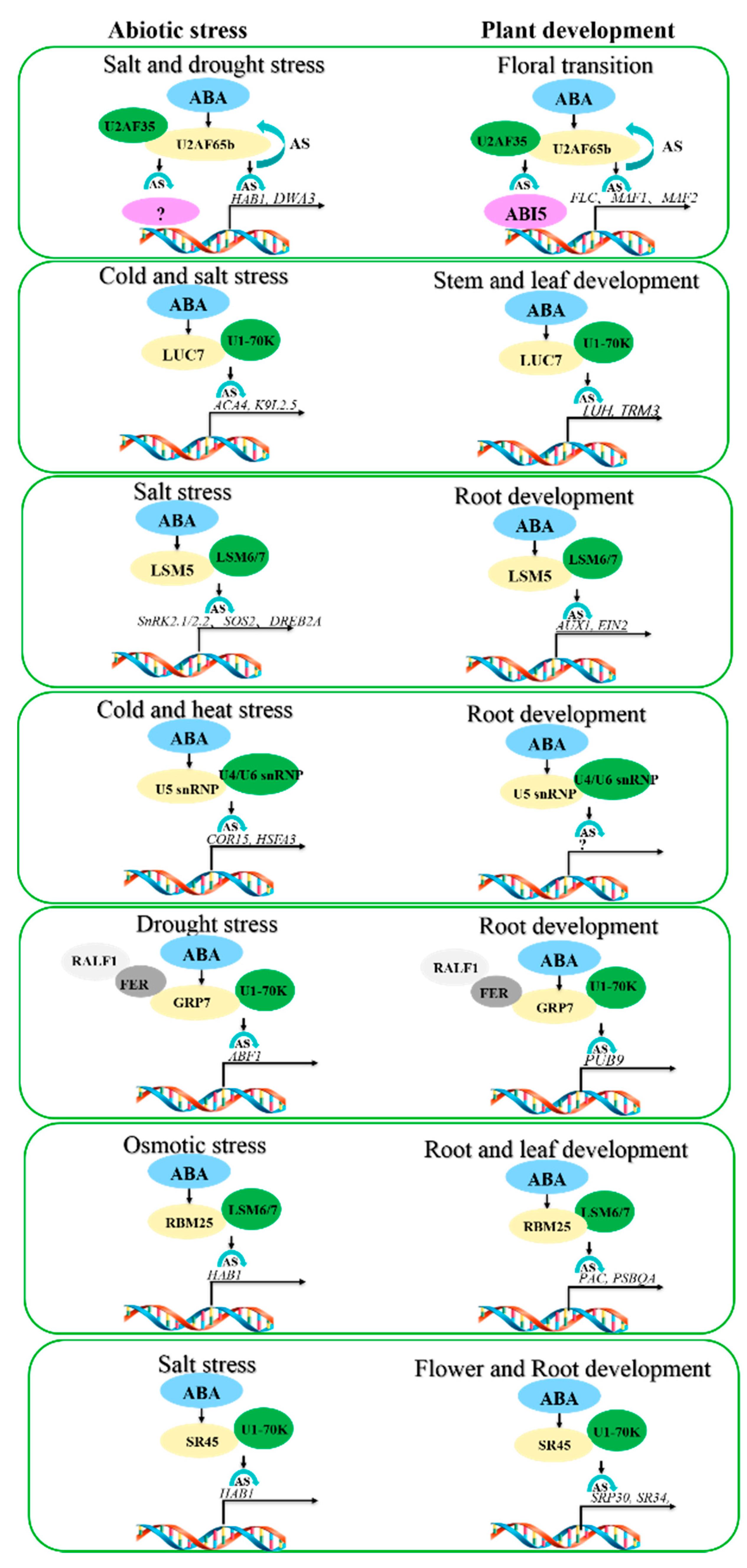
3. ABA Mediates Plant Development and Abiotic Stress via Alternative Splicing
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hada, A.; Jaabir, M.; Singh, N.; Changwal, C.; Kumar, A. Functional genomics approaches for combating the effect of abiotic stresses. In Stress Tolerance in Horticultural Crop; Woodhead Publishing: Sawston, UK, 2021; Volume 1, pp. 1–19. [Google Scholar]
- Chi, D.N.; Chen, J.; Clark, D.; Perez, H.; Huo, H.A. Effects of Maternal Environment on Seed Germination and Seedling Vigor of Petunia × hybrida under Different Abiotic Stresses. Plants 2021, 10, 581. [Google Scholar]
- Zhang, A.; Yang, X.; Lu, J.; Song, F.; Zhao, B. OsIAA20, an Aux/IAA protein, mediates abiotic stress tolerance in rice through an ABA pathway. Plant Sci. 2021, 308, 110903. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Xu, Q.; Zhao, Y.; Zhou, D.X. Histone deacetylase HDA710 controls salt tolerance by regulating ABA signaling in rice. J. Integr. Plant Biol. 2021, 63, 451–467. [Google Scholar] [CrossRef] [PubMed]
- Min, M.K.; Kim, R.; Hong, W.J.; Jung, K.H.; Kim, B.G. OsPP2C09 Is a Bifunctional Regulator in Both ABA-Dependent and Independent Abiotic Stress Signaling Pathways Citation. Int. J. Mol. Sci. 2021, 22, 393. [Google Scholar] [CrossRef]
- Shi, X.; Tian, Q.; Deng, P.; Zhang, W.; Jing, W. The rice aldehyde oxidase OsAO3 gene regulates plant growth, grain yield, and drought tolerance by participating in ABA biosynthesis. Biochem. Biophys. Res. Commun. 2021, 548, 189–195. [Google Scholar] [CrossRef]
- Black, D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.H.; Montez, J.M.; Carroll, K.M.; Darvishzadeh, J.G.; Jm, L.J.F.; Proenca, R. Abnormal splicing of the leptin receptor in diabetic mice. Nature 1996, 379, 632–635. [Google Scholar] [CrossRef]
- Punzo, P.; Grillo, S.; Batelli, G. Alternative splicing in plant abiotic stress responses. Biochem. Soc. Trans. 2020, 48, 2117–2126. [Google Scholar] [CrossRef]
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nature 2010, 463, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Stamm, S.; Ben-Ari, S.; Rafalska, I.; Tang, Y.; Zhang, Z.; Toiber, D.; Thanaraj, T.A.; Soreq, H. Function of alternative splicing. Gene 2005, 344, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Keren, H.; Lev-Maor, G.; Ast, G. Alternative splicing and evolution: Diversification, exon definition and function. Nat. Rev. Genet. 2010, 11, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, G.; Fu, C.; Duan, S.; Guo, X. Genome-wide identification, transcriptome analysis and alternative splicing events of Hsf family genes in maize. Sci. Rep. 2020, 10, 8073. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Rao, X.; Wang, Y.; Liu, Y. Roles of alternative splicing in modulating transcriptional regulation. BMC Syst. Biol. 2017, 11, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanowski, A.; Schlaen, R.G.; Perez-Santangelo, S.; Mancini, E.; Yanovsky, M.J. Global transcriptome analysis reveals circadian control of splicing events in Arabidopsis thaliana. Plant J. 2020, 103, 889–902. [Google Scholar] [CrossRef]
- Ganie, S.A.; Reddy, A. Stress-Induced Changes in Alternative Splicing Landscape in Rice: Functional Significance of Splice Isoforms in Stress Tolerance. Biology 2021, 10, 309. [Google Scholar] [CrossRef]
- Wang, T.Y.; Liu, Q.; Ren, Y.; Alam, S.K.; Yang, R. A pan-cancer transcriptome analysis of exitron splicing identifies novel cancer driver genes and neoepitopes. Mol. Cell 2021, 81, 2246–2260. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, Y.; Zhang, M.; Zhu, F.; Sun, M.; Lian, X.; Zhao, G.; Duan, D. Transcriptome and metabolome analysis of stress tolerance to aluminium in Vitis quinquangularis. Planta 2021, 254, 105. [Google Scholar] [CrossRef]
- Holliday, M.; Singer, E.S.; Ross, S.B.; Lim, S.; Bagnall, R.D. Transcriptome Sequencing of Patients with Hypertrophic Cardiomyopathy Reveals Novel Splice-altering Variants in MYBPC3. Circ. Genom. Precis. Med. 2021, 14, e003202. [Google Scholar] [CrossRef]
- Noam, L.; Noam, A.; Dena, L.; Robert, F.; Shin-Han, S. Genome-Wide Survey of Cold Stress Regulated Alternative Splicing in Arabidopsis thaliana with Tiling Microarray. PLoS ONE 2013, 8, e66511. [Google Scholar] [CrossRef] [Green Version]
- Mario, K.; Hu, Y.; Anida, M.; Sotirios, F.; Enrico, S.; Stefan, S. Alternative splicing in tomato pollen in response to heat stress. DNA Res. 2017, 24, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Seo, P.J.; Park, M.J.; Park, C.M. Alternative splicing of transcription factors in plant responses to low temperature stress: Mechanisms and functions. Planta 2013, 237, 1415–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takechi, H.; Hosokawa, N.; Hirayoshi, K.; Nagata, K. Alternative 5′ splice site selection induced by heat shock. Mol. Cell. Biol. 1994, 14, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barashkov, N.A.; Dzhemileva, L.U.; Fedorova, S.A.; Teryutin, F.M.; Posukh, O.L.; Fedotova, E.E.; Lobov, S.L.; Khusnutdinova, E.K. Autosomal recessive deafness 1A (DFNB1A) in Yakut population isolate in Eastern Siberia: Extensive accumulation of the splice site mutation IVS1+1G>A in GJB2 gene as a result of founder effect. J. Hum. Genet. 2011, 56, 631–639. [Google Scholar] [CrossRef]
- Li, S.; Yamada, M.; Han, X.; Ohler, U.; Benfey, P. High-Resolution Expression Map of the Arabidopsis Root Reveals Alternative Splicing and lincRNA Regulation. Dev. Cell 2016, 39, 508–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarup, R.; Crespi, M.; Bennett, M.J. One Gene, Many Proteins: Mapping Cell-Specific Alternative Splicing in Plants. Dev. Cell 2016, 39, 383–385. [Google Scholar] [CrossRef] [Green Version]
- Sallam, N.; Moussa, M. DNA methylation changes stimulated by drought stress in ABA-deficient maize mutant vp10. Plant Physiol. Biochem. 2021, 160, 218–224. [Google Scholar] [CrossRef]
- Zhan, X.; Qian, B.; Cao, F.; Wu, W.; Yang, L.; Guan, Q.; Gu, X.; Wang, P.; Okusolubo, T.A.; Dunn, S.L.; et al. An Arabidopsis PWI and RRM motif-containing protein is critical for pre-mRNA splicing and ABA responses. Nat. Commun. 2015, 6, 8139. [Google Scholar] [CrossRef] [Green Version]
- Ling, Y.; Alshareef, S.; Butt, H.; Lozano-Juste, J.; Mahfouz, M.M. Pre-mRNA splicing repression triggers abiotic stress signaling in plants. Plant J. 2016, 89, 291–309. [Google Scholar] [CrossRef]
- Punkkinen, M.; Mahfouz, M.M.; Fujii, H. Chemical activation of Arabidopsis SnRK2.6 by pladienolide B. Plant Signal. Behav. 2021, 16, 1885165. [Google Scholar] [CrossRef]
- Xue, X.; Jiao, F.; Xu, H.; Jiao, Q.; Wang, M. The role of RNA-binding protein, microRNA and alternative splicing in seed germination: A field need to be discovered. BMC Plant Biol. 2021, 21, 194. [Google Scholar] [CrossRef]
- Laloum, T.; Martín, G.; Duque, P. Alternative Splicing Control of Abiotic Stress Responses. Trends Plant Sci. 2017, 23, 140–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, T.; Carvalho, R.F.; Richardson, D.N.; Duque, P. Abscisic Acid (ABA) Regulation of Arabidopsis SR Protein Gene Expression. Int. J. Mol. Sci. 2014, 15, 17541–17564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.C.; Sheng, L. ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 2012, 35, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Hai, A.T.; Lee, S.; Cao, S.T.; Lee, W.J.; Lee, H. Overexpression of the HDA15 Gene Confers Resistance to Salt Stress by the Induction of NCED3, an ABA Biosynthesis Enzyme. Front. Plant Sci. 2021, 12, 631. [Google Scholar]
- Toups, H.S.; Cochetel, N.; Galdamez, K.; Deluc, L.; Cramer, G.R. Abscisic Acid Metabolism in Leaves and Roots of Four Vitis Species in Response to Water Deficit; Research Square: Durham, NC, USA, 2021. [Google Scholar]
- Hwang, S.G.; Chen, H.C.; Huang, W.Y.; Chu, Y.C.; Shii, C.T.; Cheng, W.H. Ectopic expression of rice OsNCED3 in Arabidopsis increases ABA level and alters leaf morphology. Plant Sci. 2010, 178, 12–22. [Google Scholar] [CrossRef]
- Chuong, N.N.; Xuan, L.; Nghia, D.; Dai, T.; Thao, N.P. Protein Phosphatase Type 2C Functions in Phytohormone-Dependent Pathways and in Plant Responses to Abiotic Stresses. Curr. Protein Pept. Sci. 2021, 22, 430–440. [Google Scholar] [CrossRef]
- Collin, A.; Daszkowska-Golec, A.; Szarejko, I. Updates on the Role of ABSCISIC ACID INSENSITIVE 5 (ABI5) and ABSCISIC ACID-RESPONSIVE ELEMENT BINDING FACTORs (ABFs) in ABA Signaling in Different Developmental Stages in Plants. Cells 2021, 10, 1996. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef] [Green Version]
- Soma, F.; Takahashi, F.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Cellular Phosphorylation Signaling and Gene Expression in Drought Stress Responses: ABA-Dependent and ABA-Independent Regulatory Systems. Plants 2021, 10, 756. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Rep. 2013, 32, 959–970. [Google Scholar] [CrossRef]
- Yoshida, T.; Fujita, Y.; Maruyama, K.; Mogami, J.; Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Four Arabidopsis AREB/ABF transcription factors function predominantly in gene expression downstream of SnRK2 kinases in abscisic acid signalling in response to osmotic stress. Plant Cell Environ. 2015, 38, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Li, Y.; Wang, Y.; Liu, X.; Wang, P. Initiation and amplification of SnRK2 activation in abscisic acid signaling. Nat. Commun. 2021, 12, 2456. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, Y.; Zhang, W.; Zheng, Y.; Yuan, B.; Li, Q.; Leng, P. Tomato SlPP2C5 Is Involved in the Regulation of Fruit Development and Ripening. Plant Cell Physiol. 2021, 11, 1760–1769. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hu, J.; Fan, W.; Zhu, P.; Cao, B.; Zheng, S.; Xia, Z.; Zhu, Y.; Zhao, A. Heterotrimeric G-protein γ subunits regulate ABA signaling in response to drought through interacting with PP2Cs and SnRK2s in mulberry (Morus alba L.). Plant Physiol. Biochem. 2021, 161, 210–221. [Google Scholar] [CrossRef]
- Geiger, D.; Scherzer, S.; Mumm, P.; Marten, I.; Ache, P. Guard cell anion channel SLAC1 is regulated by CDPK protein kinases with distinct Ca2+ affinities. Proc. Natl. Acad. Sci. USA 2010, 107, 8023–8028. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.; Panzeri, D.; Okuma, E.; Tldsepp, K.; Zimmerli, L. STRESS INDUCED FACTOR 2 Regulates Arabidopsis Stomatal Immunity through Phosphorylation of the Anion Channel SLAC1. Plant Cell 2020, 32, 2216–2236. [Google Scholar] [CrossRef]
- Sato, A.; Sato, Y.; Fukao, Y.; Fujiwara, M.; Uozumi, N. Threonine at position 306 of the KAT1 potassium channel is essential for channel activity and is a target site for ABA-activated SnRK2/OST1/SnRK2.6 protein kinase. Biochem. J. 2009, 424, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Kuromori, T.; Seo, M.; Shinozaki, K. ABA Transport and Plant Water Stress Responses. Trends Plant Sci. 2018, 23, 513–522. [Google Scholar] [CrossRef]
- Nian, L.; Zhang, X.; Yi, X.; Liu, X.; Yang, Y.; Li, X.; Haider, F.U.; Zhu, X. Genome-wide identification of ABA receptor PYL/RCAR gene family and their response to cold stress in Medicago sativa L. Physiol. Mol. Biol. Plants 2021, 27, 1979–1995. [Google Scholar] [CrossRef]
- Ibrahim, A.K.; Xu, Y.; He, Q.; Niyitanga, S.; Zhang, L. Comparative Transcriptomes and Genome-Wide Identification Reveal Salt Stress-Responsive PP2C in Jute (Corchorus capsularis); Research Square: Durham, NC, USA, 2021. [Google Scholar]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 27–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, Y.; Yokohama, R.; Reed, J.J.; Liu, H.Y.; Lu, G. Citation: The Arabidopsis SMALL AUXIN UP RNA32 Protein Regulates ABA-Mediated Responses to Drought Stress. Front. Plant Sci. 2021, 12, 259. [Google Scholar]
- Wu, H.; Li, H.; Zhang, W.; Tang, H.; Yang, L. Transcriptional regulation and functional analysis of Nicotiana tabacum under salt and ABA stress—ScienceDirect. Biochem. Biophys. Res. Commun. 2021, 570, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Adams, S.; Grundy, J.; Veflingstad, S.R.; Dyer, N.P.; Hannah, M.A.; Ott, S.; Carré, I.A. Circadian control of abscisic acid biosynthesis and signalling pathways revealed by genome-wide analysis of LHY binding targets. New Phytol. 2018, 220, 893–907. [Google Scholar] [CrossRef]
- Baek, D.; Kim, W.Y.; Cha, J.Y.; Park, H.J.; Yun, D.J. The GIGANTEA-ENHANCED EM LEVEL complex enhances drought tolerance via regulation of abscisic acid synthesis. Plant Physiol. 2020, 184, 443–458. [Google Scholar] [CrossRef]
- Wang, Z.; Ren, Z.; Cheng, C.; Wang, T.; Ji, H.; Zhao, Y.; Deng, Z.; Zhi, L.; Lu, J.; Wu, X.; et al. Counteraction of ABA-Mediated Inhibition of Seed Germination and Seedling Establishment by ABA Signaling Terminator in Arabidopsis. Mol. Plant 2020, 13, 1284–1297. [Google Scholar] [CrossRef]
- Hong, J.H.; Seah, S.W.; Xu, J. The root of ABA action in environmental stress response. Plant Cell Rep. 2013, 32, 971–983. [Google Scholar] [CrossRef]
- Shu, K.; Qian, C.; Wu, Y.; Liu, R.; Zhang, H.; Wang, S.; Tang, S.; Yang, W.; Xie, Q. ABSCISIC ACID-INSENSITIVE 4 negatively regulates flowering through directly promoting Arabidopsis FLOWERING LOCUS C transcription. J. Exp. Bot. 2016, 67, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Do, V.G.; Kim, S.; Kweon, H.; Mcghie, T.K. Cold stress triggers premature fruit abscission through ABA-dependent signal transduction in early developing apple. PLoS ONE 2021, 16, e0249975. [Google Scholar] [CrossRef]
- Lopez-Molina, L.; Mongrand, S.; Chua, N.H. A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis. Proc. Natl. Acad. Sci. USA 2001, 98, 4782–4787. [Google Scholar] [CrossRef] [Green Version]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The Role and Regulation of ABI5 (ABA-Insensitive 5) in Plant Development, Abiotic Stress Responses and Phytohormone Crosstalk. Front. Plant Sci. 2016, 7, 1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, L.; Ye, T.; Lu, Y.; Chen, X.; Wu, Y. The inhibitory effect of ABA on floral transition is mediated by ABI5 in Arabidopsis. J. Exp. Bot. 2013, 64, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Günter, T.E.; Heinz, S. The golden decade of molecular floral development (1990–1999): A cheerful obituary. Dev. Genet. 2015, 25, 181–193. [Google Scholar]
- Zhu, F.Y.; Chen, M.X.; Ye, N.H.; Shi, L.; Ma, K.L.; Yang, J.F.; Cao, Y.Y.; Zhang, Y.; Yoshida, T.; Fernie, A.R. Proteogenomic analysis reveals alternative splicing and translation as part of the abscisic acid response in Arabidopsis seedlings. Plant J. 2017, 91, 518–533. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Ji, L.; Wang, J.; Jin, J.; Yang, X.; Rao, P.; Gao, K.; Liao, W.; Ye, M.; An, X. Dynamic changes in the transcriptome of Populus hopeiensis in response to abscisic acid. Sci. Rep. 2017, 7, 42708. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Oubridge, C.; Van Roon, A.M.; Nagai, K. Crystal structure of human U1 snRNP, a small nuclear ribonucleoprotein particle, reveals the mechanism of 5′ splice site recognition. eLife 2015, 4, e04986. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Xiong, F.; Ren, Q.P.; Wang, X.L. Regulation of flowering transition by alternative splicing: The role of the U2 auxiliary factor. J. Exp. Bot. 2019, 71, 751–758. [Google Scholar] [CrossRef]
- Palusa, S.G.; Ali, G.S.; Reddy, A.S. Alternative splicing of pre-mRNAs of Arabidopsis serine/arginine-rich proteins: Regulation by hormones and stresses. Plant J. 2007, 49, 1091–1107. [Google Scholar] [CrossRef]
- Filichkin, S.A.; Priest, H.D.; Givan, S.A.; Shen, R.; Bryant, D.W.; Fox, S.E.; Wong, W.-K.; Mockler, T.C. Genome-wide mapping of alternative splicing in Arabidopsis thaliana. Genome Res. 2010, 20, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Albaqami, M.; Laluk, K.; Reddy, A. The Arabidopsis splicing regulator SR45 confers salt tolerance in a splice isoform-dependent manner. Plant Mol. Biol. 2019, 100, 379–390. [Google Scholar] [CrossRef]
- Zhang, X.N.; Mount, S.M. Two Alternatively Spliced Isoforms of the Arabidopsis SR45 Protein Have Distinct Roles during Normal Plant Development. Plant Physiol. 2009, 150, 1450–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, D.; Wang, Y.; Hamilton, M.; Ben-Hur, A.; Reddy, A. Transcriptome-Wide Identification of RNA Targets of Arabidopsis SERINE/ARGININE-RICH45 Uncovers the Unexpected Roles of This RNA Binding Protein in RNA Processing. Plant Cell 2015, 27, 3294–3308. [Google Scholar] [CrossRef] [PubMed]
- Tillemans, V.; Dispa, L.; Remacle, C.; Collinge, M.; Motte, P. Functional distribution and dynamics of Arabidopsis SR splicing factors in living plant cells. Plant J. 2005, 41, 567–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, F.; Ren, J.J.; Yu, Q.; Wang, Y.Y.; Lu, C.C.; Kong, L.J.; Otegui, M.S.; Wang, X.L. AtU2AF65b functions in abscisic acid mediated flowering via regulating the precursor messenger RNA splicing of ABI5 and FLC in Arabidopsis. New Phytol. 2019, 223, 277–292. [Google Scholar] [CrossRef] [PubMed]
- Airoldi, C.A.; McKay, M.; Davies, B. MAF2 Is Regulated by Temperature-Dependent Splicing and Represses Flowering at Low Temperatures in Parallel with FLM. PLoS ONE 2015, 10, e0126516. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, S.K.; Kanwar, P.; Samtani, H.; Kaur, K.; Jha, S.K.; Pandey, G.K. Alternative Splicing of CIPK3 Results in Distinct Target Selection to Propagate ABA Signaling in Arabidopsis. Front. Plant Sci. 2017, 8, 1924. [Google Scholar] [CrossRef] [Green Version]
- Marcella, D.; Willing, E.M.; Szabo, E.X.; Francisco-Mangilet, A.G.; Droste-Borel, I.; Macek, B.; Schneeberger, K.; Laubinger, S. The U1 snRNP subunit LUC7 modulates plant development and stress responses via regulation of alternative splicing. Plant Cell 2018, 30, 2838–2854. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.; Zhang, S.; Feng, D.; Ali, S.; Xiong, L. Dynamic regulation of genome-wide pre-mRNA splicing and stress tolerance by the Sm-like protein LSm5 in Arabidopsis. Genome Biol. 2014, 15, R1. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.H.; Kapoor, A.; Zhu, J.; Zhu, J.K. STABILIZED1, a Stress-Upregulated Nuclear Protein, Is Required for Pre-mRNA Splicing, mRNA Turnover, and Stress Tolerance in Arabidopsis. Plant Cell 2006, 18, 1736–1749. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.D.; Cho, Y.H.; Lee, B.H.; Yoo, S.D. STABILIZED1 modulates pre-mRNA splicing for thermotolerance. Plant Physiol. 2017, 173, 2370–2382. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, T.; Wang, B.; Lin, Q.; Yu, F. RALF1-FERONIA complex affects splicing dynamics to modulate stress responses and growth in plants. Sci. Adv. 2020, 6, eaaz1622. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.K.; Jung, H.J.; Lee, H.J.; Kim, K.A.; Kang, H. Glycine-rich RNA-binding protein 7 affects abiotic stress responses by regulating stomata opening and closing in Arabidopsis thaliana. Plant J. 2010, 55, 455–466. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Kanwar, P.; Yadav, A.K.; Sharma, C.; Kumar, A.; Pandey, G.K. Arabidopsis CBL interacting protein kinase 3 interacts with ABR1, an APETALA2 domain transcription factor, to regulate ABA responses. Plant Sci. 2017, 254, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Harb, A.; Simpson, C.; Guo, W.; Govindan, G.; Kakani, V.G.; Sunkar, R. The Effect of Drought on Transcriptome and Hormonal Profiles in Barley Genotypes With Contrasting Drought Tolerance. Front. Plant Sci. 2020, 11, 618491. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Plants | Major DAS Type | References |
|---|---|---|
| Arabidopsis | AL > AF > AE > RI > SE > MX | [67] |
| Nicotiana tabacum | SE > RI > A3 > A5 > MX | [56] |
| Populus hopeiensis | RI > A3 > A5 > SE > MX | [68] |
| Locus | Gene | Transcript Type |
|---|---|---|
| AT1G17550 | HAB2 | HAB2-iso1,2,3 |
| AT1G45249 | ABRE1 | ABRE1-iso1,2,3 |
| AT4G34000 | ABF3 | ABF1-iso1,2,3 |
| AT5G25610 | RD22 | RD22-iso1,2 |
| AT4G27410 | RD26 | RD26-iso1,2,3,4 |
| AT1G20620 | CAT3 | CAT3-iso1,2,3,4 |
| AT5G62470 | MYB96 | MYB96-iso1,2 |
| AT1G20630 | CAT1 | CAT1-iso1,2 |
| AT4G46270 | GBF3 | GBF3-iso1,2,3,4,5,6 |
| AT4G19230 | CYP707A1 | CYP707A1-iso1,2 |
| AT4G26080 | ABI1 | ABI1-iso1,2 |
| AT5G57050 | ABI2 | ABI2-iso1,2,3 |
| Locus | Splicing Factors |
|---|---|
| AT1G02840 | SR34 |
| AT4G02430 | SR34b |
| AT1G09140 | SR30 |
| AT3G61860 | RS31 |
| AT2G46610 | RS31a |
| AT4G25500 | RS40 |
| AT5G52040 | RS41 |
| AT2G24590 | RSZ22A |
| AT3G53500 | RSZ32 |
| AT2G37340 | RSZ33 |
| AT5G64200 | SC35 |
| AT1G55310 | SCL33 |
| AT3G55460 | SCL30 |
| AT3G13570 | SCL30A |
| AT1G16610 | SR45 |
| AT3G50670 | U1-70K |
| AT1G27650 | U2AF35A |
| AT1G60900 | U2AF65B |
| AT2G43810 | LSM6B |
| Gene ID | Gene Name | AS Mediated Development and Abiotic Stress | References |
|---|---|---|---|
| AT2G26980 | AtCIPK3 | ABA, osmotic stress, and drought stress | [79,86] |
| At5g43270 | AtSPL2 | shoot maturation | [29] |
| At3g62600 | AtATERDJ3B | protein folding | [29] |
| At3g15970 | AtNUP50 | intracellular transport | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Jia, Z.; Pu, Q.; Tian, Y.; Zhu, F.; Liu, Y. ABA Mediates Plant Development and Abiotic Stress via Alternative Splicing. Int. J. Mol. Sci. 2022, 23, 3796. https://doi.org/10.3390/ijms23073796
Yang X, Jia Z, Pu Q, Tian Y, Zhu F, Liu Y. ABA Mediates Plant Development and Abiotic Stress via Alternative Splicing. International Journal of Molecular Sciences. 2022; 23(7):3796. https://doi.org/10.3390/ijms23073796
Chicago/Turabian StyleYang, Xue, Zichang Jia, Qiong Pu, Yuan Tian, Fuyuan Zhu, and Yinggao Liu. 2022. "ABA Mediates Plant Development and Abiotic Stress via Alternative Splicing" International Journal of Molecular Sciences 23, no. 7: 3796. https://doi.org/10.3390/ijms23073796
APA StyleYang, X., Jia, Z., Pu, Q., Tian, Y., Zhu, F., & Liu, Y. (2022). ABA Mediates Plant Development and Abiotic Stress via Alternative Splicing. International Journal of Molecular Sciences, 23(7), 3796. https://doi.org/10.3390/ijms23073796