
Identification of B-Cell Epitopes for Eliciting Neutralizing Antibodies against the SARS-CoV-2 Spike Protein through Bioinformatics and Monoclonal Antibody Targeting

 ,
,
Abstract
:1. Introduction
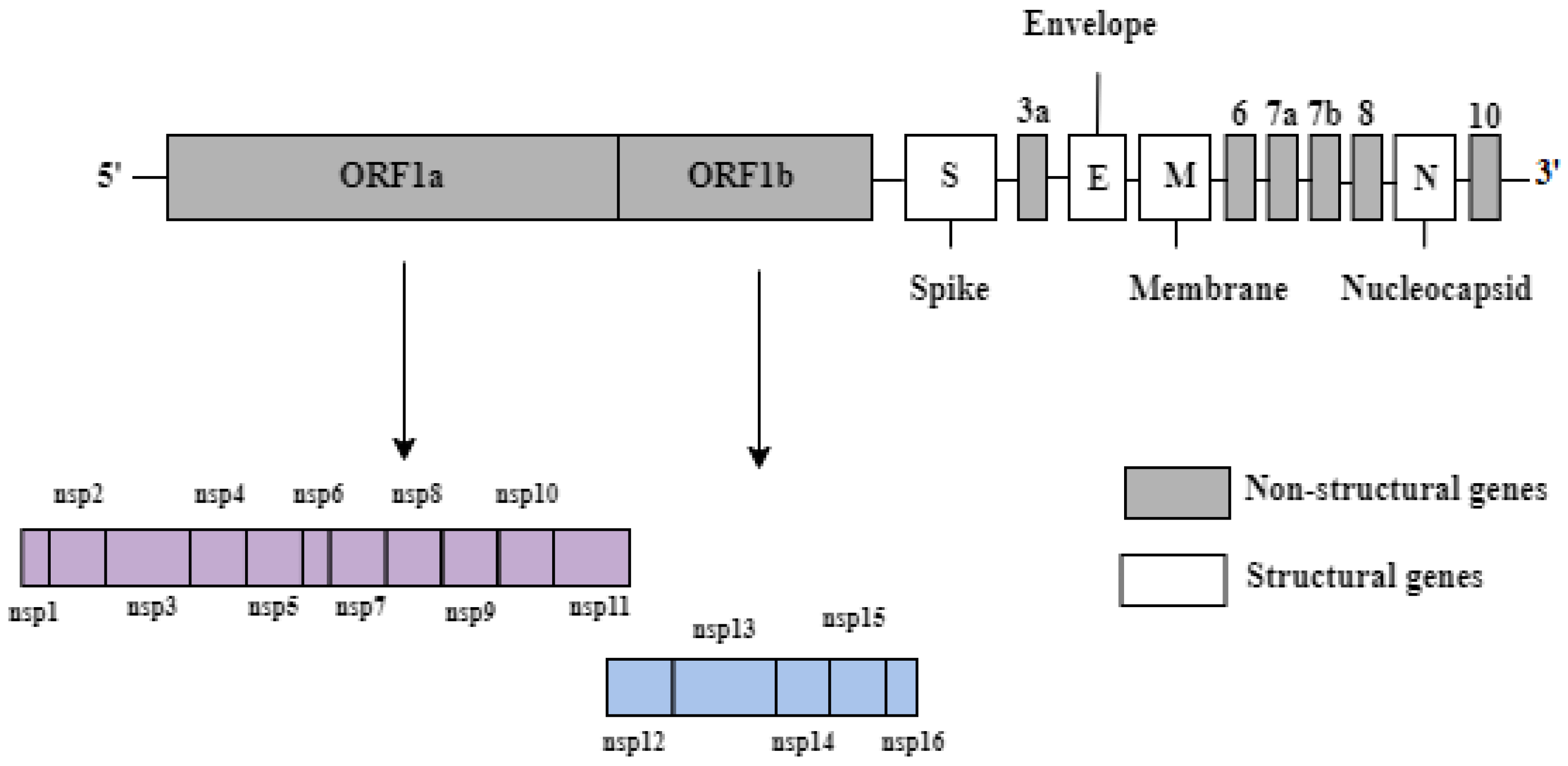
2. Genomic Structure of SARS-CoV-2
3. Viral Binding and Entry into Host Cell and the SARS-CoV-2 Lifecycle
4. Current Status of SARS-CoV-2 Vaccine
5. Approaches to Rational Design of Peptide Vaccines
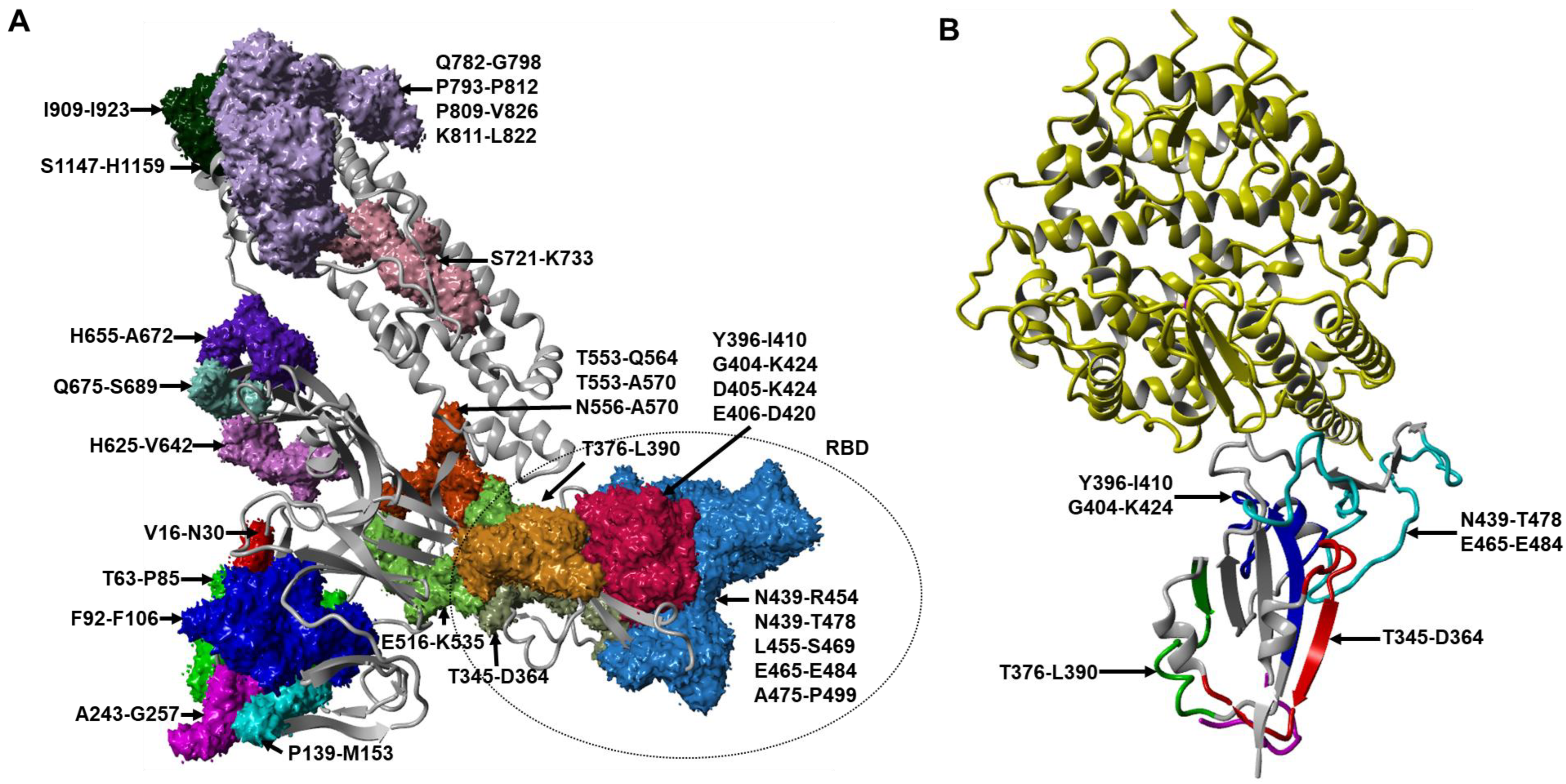
6. Identification of SARS-CoV-2 B-Cell Epitopes within S, M, and N Proteins from the Combination of Bioinformatics and In Vitro Neutralization Assays
7. Monoclonal Antibodies against SARS-CoV-2 RBD Protein
8. Conservancy of Linear B-Cell Epitopes against SARS-CoV-2 Variants
9. Conclusions

{kind=link}
{kind=link}
| mAb | Sources | Target | Efficacy | Protection | Reference |
|---|---|---|---|---|---|
| B38 | Peripheral blood of SARS-CoV-2- infected patients | RBD | LV neutralization: IC50 = 0.177 µg/mL | Protection of mice: Lung viral loads reduced by 32.8% compared with PBS control. | [39] |
| H4 | Peripheral blood of SARS-CoV-2- infected patients | RBD | LV neutralization: IC50 = 0.896 µg/mL | Protection of mice: Lung viral loads reduced by 26% compared with PBS control. | [39] |
| 414-1 | Peripheral blood of SARS-CoV-2- infected patients | RBD | LV neutralization IC50= 1.75 nM | N/A | [37] |
| MD65 | Phage display library constructed using peripheral circulatory lymphocyte of SARS-CoV-2- infected patients | RBD | LV neutralization NT50= 0.22 µg/mL | N/A | [41] |
| COVA1–18 | B cells of convalescent patients | RBD | PsV neutralization: IC50 = 0.008 µg/mL LV neutralization: IC50 = 0.007 µg/mL | N/A | [43] |
| COVA2-15 | B cells of convalescent patients | RBD | PsV neutralization: IC50 = 0.008 µg/mL LV neutralization: IC50 = 0.009 µg/mL | N/A | [43] |
| 2-15 | Peripheral blood of COVID-19 patients | RBD | PsV neutralization: IC50 = 0.7 ng/mL LV neutralization: IC50 = 5 ng/mL | Protection of hamsters: Viral RNA copy numbers and infectious virus titers in lung tissues were reduced by 4 logs or more compared with the PBS control. | [44] |
| S309 | Peripheral blood of SARS-infected patients | RBD | PsV neutralization: IC50 = 120~180 ng/mL | N/A | [42] |
| 3F11 | Humanized phage display library | RBD | PsV neutralization: IC50 = 3.8 ng/mL LV neutralization: IC50 = 436 ng/mL. | N/A | [48] |
| 4A8 | Peripheral blood of COVID-19 convalescent patients | NTD (in S1) | PsV neutralization: EC50 = 49 μg/mL LV neutralization: EC50 = 0.61 μg/mL | N/A | [49] |
| CR3022 | Gene cloning; Protein expression | RBD | LV neutralization: IC50 = ~0.114 μg/mL | N/A | |
| CB6 | B cells of convalescent patients | RBD | PsV neutralization: ND50 = 0.036 µg/mL LV neutralization: ND50 = 0.036 µg/mL | Protection of rhesus macaques: 50 mg/kg | [40] |
| S2P6 | Memory B cells of SARS-CoV-2 patients | S2 | LV neutralization: IC50 = 1.67 µg/mL PsV D614G: IC50~10 µg/mL PsV P.1: IC50~10 µg/mL PsV B.1.1.7: IC50~100 µg/mL PsV B.1.351: IC50~100 µg/mL PsV 1.1.617: IC50~20 µg/mL | Protection of hamsters: Viral RNA copy numbers in lung tissues were reduced by 2 logs and 1.5 logs against SARS-CoV-2 Wuhan strain and B.1.351 Beta strain. | [45] |
| Protein | MonoclonalAntibody | Peptide ID | Start-End (aa) | Sequences of B-Cell Epitopes | Methods | Host | Inhibition | Conservancy (%) | Reference |
|---|---|---|---|---|---|---|---|---|---|
| S1 S2 | N/A | S14P5 S21P2 | 553–570 809–826 | TESNKKFLPFQQFGRDIA PSKPSKRSFIEDLLFNKV | Overlapping peptide library | COVID-19 sera | >20% of pseudoviruses >20% of pseudoviruses | 84 100 | [28] |
| S1 S2 S2 | N/A | N/A | 655–672 782–798/811–822 1147–1158 | HVNNSYECDIPIGAGICA QIYKTPPIKDFG/KPSKRSFIEDLL SFKEELDKYFKN | Peptide array | COVID-19 plasma | N/A | 93.50 98/100 100 | [27] |
| S1/S2 S2 | N/A | N/A | 675–689 721–733 | QTQTNSPRRARSVAS SVTTEILPVSMTK | Epitope predictions based on 3D protein structure, epitope surface accessibility | COVID-19 sera, BALB/c mice | ~50% of G614 pseudoviruses No inhibition of D614 pseudoviruses | 60.5 98.5 | [30] |
| S1 S1 RBD RBD S1 S2 S2 | N/A | N/A | 16–30 243–257 406–420 475–499 556–570 793–812(N) 909–923 | VNLTTRTQLPPAYTN ALHRSYLTPGDSSSG EVRQIAPGQTGKIAD AGSTPCNGVEGFNCYFPLQSYGFQP NKKFLPFQQFGRDIA PIKDFGGFN(GlcNAc)FSQILPDPSKP IGVTQNVLYENQKLI | 20–40% inhibition of D614 pseudoviruses | 84 85.50 88 53.50 84 99 99.50 | |||
| S1 S1 RBD RBD | N/A | N/A | 92–106 139–153 439–454 455–469 | FASTEKSNIIRGWIF PFLGVYYHKNNKSWM NNLDSKVGGNYNYLYR LFRKSNLKPFERDIS | 40–50% inhibition of D614, G614 pseudoviruses | 89.50 62 76.50 100 | |||
| S1 | N/A | N/A | 63–85 | TWFHAIHVSGTNGTKRFDNPVLP | >80% inhibition of G614 pseudoviruses | 65.50 | |||
| RBD RBD RBD | 15G9 12C10 10D2 | R345 R405 R465 | 345–364 405–424 465–484 | TRFASVYAWNRKRISNCVAD DEVRQIAPGQTGKIADYNYK ERDISTEIYQAGSTPCNGVE | Overlapping peptides covering RBD | Swine and mice | 60% of RBD/ACE2 interaction 40% of RBD/ACE2 interaction 20% of RBD/ACE2 interaction | 100 88 57 | [31] |
| RBD RBD RBD | N/A | CoV2_S-10 CoV2_S-11 CoV2_S-13 | 404–424 439–478 516–535 | GDEVRQIAPGQTGKIADYNYK NNLDSKVGGNYNYLYRLFRKSNLKPFERDISTEIYQAGST ELLHAPATVCGPKKSTNLVK | Immunoinformatic prediction (Bepipred-2.0) | Cynomolgus macaques | N/A | 88 74 98.50 | [35] |
| S1 S1 S2 | N/A | S1-93 S1-105 S2-78 | 553–564 625–642 1148–1159 | TESNKKFLPFQQ HADQLTPTWRVYSTGSNV FKEELDKYFKNH | Peptide microarray | COVID-19 sera | 51% of pseudoviruses 35% of pseudoviruses 35% of pseudoviruses | 99 99.5 100 | [34] |
| RBD RBD | Ab 24 & 67 Ab 69 & 90 | N/A | 376–390 396–410 | TFKCYGVSPTKLNDL YADSFVIRGDEVRQI | Overlapping peptides covering S protein | Rabbit sera | N/A | 86.67 53.33 | [38] |
| S2 | S2P6 | N/A | 1148–1156 | KEELDKYF | X-ray crystallography and Cryo-EM | COVID-19 sera | >90% inhibition of live viruses | 100 | [45] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grant, M.C.; Geoghegan, L.; Arbyn, M.; Mohammed, Z.; McGuinness, L.; Clarke, E.L.; Wade, R.G. The prevalence of symptoms in 24,410 adults infected by the novel coronavirus (SARS-CoV-2; COVID-19): A systematic review and meta-analysis of 148 studies from 9 countries. PLoS ONE 2020, 15, e0234765. [Google Scholar] [CrossRef] [PubMed]
- Vallamkondu, J.; John, A.; Wani, W.Y.; Ramadevi, S.P.; Jella, K.K.; Reddy, P.H.; Kandimalla, R. SARS-CoV-2 pathophysiology and assessment of coronaviruses in CNS diseases with a focus on therapeutic targets. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165889. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Velavan, T.P.; Meyer, C.G. The COVID-19 epidemic. Trop. Med. Int. Health 2020, 25, 278–280. [Google Scholar] [CrossRef] [Green Version]
- Jungreis, I.; Sealfon, R.; Kellis, M. SARS-CoV-2 gene content and COVID-19 mutation impact by comparing 44 Sarbecovirus genomes. Nat. Commun. 2021, 12, 2642. [Google Scholar] [CrossRef]
- Alanagreh, L.; Alzoughool, F.; Atoum, M. The human coronavirus disease COVID-19: Its origin, characteristics, and insights into potential drugs and its mechanisms. Pathogens 2020, 9, 331. [Google Scholar] [CrossRef]
- Rastogi, M.; Pandey, N.; Shukla, A.; Singh, S.K. SARS coronavirus 2: From genome to infectome. Respir. Res. 2020, 21, 318. [Google Scholar] [CrossRef]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Zhang, S.; Wang, Q.; Anang, S.; Wang, J.; Ding, H.; Kappes, J.C.; Sodroski, J. Spike glycoprotein and host cell determinants of SARS-CoV-2 entry and cytopathic effects. J. Virol. 2020, 95, e02304-20. [Google Scholar] [CrossRef]
- Ahmed, S.; Khan, M.S.; Gayathri, S.; Singh, R.; Kumar, S.; Patel, U.R.; Malladi, S.K.; Rajmani, R.S.; van Vuren, P.J.; Riddell, S.; et al. A stabilized, monomeric, receptor binding domain elicits high-titer neutralizing antibodies against all SARS-CoV-2 variants of concern. Front Immunol. 2021, 12, 765211. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccoli, L.; Park, Y.-J.; Tortorici, M.A.; Czudnochowski, N.; Walls, A.C.; Beltramello, M.; Silacci-Fregni, C.; Pinto, D.; Rosen, L.E.; Bowen, J.E.; et al. Mapping neutralizing and immunodominant sites on the SARS-CoV-2 spike receptor-binding domain by structure-guided high-resolution serology. Cell 2020, 183, 1024–1042.e21. [Google Scholar] [CrossRef] [PubMed]
- Collie, S.; Champion, J.; Moultrie, H.; Bekker, L.-G.; Gray, G. Effectiveness of BNT162b2 vaccine against Omicron variant in South Africa. N. Engl. J. Med. 2021, 386, 494–496. [Google Scholar] [CrossRef] [PubMed]
- Cele, S.; Jackson, L.; Khan, K.; Khoury, D.; Moyo-Gwete, T.; Tegally, H.; Scheepers, C.; Amoako, D.; Karim, F.; Bernstein, M.; et al. SARS-CoV-2 Omicron has extensive but incomplete escape of Pfizer BNT162b2 elicited neutralization and requires ACE2 for infection. medRxiv 2021. [Google Scholar] [CrossRef]
- Rössler, A.; Riepler, L.; Bante, D.; von Laer, D.; Kimpel, J. SARS-CoV-2 omicron variant neutralization in serum from vaccinated and convalescent persons. N. Engl. J. Med. 2022, 386, 698–700. [Google Scholar] [CrossRef]
- Pfizer and BioNTech Initiate Study to Evaluate Omicron-Based COVID-19 Vaccine in Adults 18 to 55 Years of Age. Available online: https://www.pfizer.com/news/press-release/press-release-detail/pfizer-and-biontech-initiate-study-evaluate-omicron-based (accessed on 15 February 2022).
- New Studies Look to Assess Immunogenicity of Omicron-Based Vaccines. Available online: https://www.europeanpharmaceuticalreview.com/news/168012/studies-assess-immunogenicity-omicron-based-vaccines/ (accessed on 15 February 2022).
- Garcia-Beltran, W.F.; St Denis, K.J.; Hoelzemer, A.; Lam, E.C.; Nitido, A.D.; Sheehan, M.L.; Berrios, C.; Ofoman, O.; Chang, C.C.; Hauser, B.M.; et al. mRNA-based COVID-19 vaccine boosters induce neutralizing immunity against SARS-CoV-2 Omicron variant. Cell 2022, 185, 457–466.e4. [Google Scholar] [CrossRef]
- Pajon, R.; Doria-Rose, N.A.; Shen, X.; Schmidt, S.D.; O’Dell, S.; McDanal, C.; Feng, W.; Tong, J.; Eaton, A.; Maglinao, M.; et al. SARS-CoV-2 omicron variant neutralization after mRNA-1273 booster vaccination. N. Engl. J. Med. 2022, 386, 1088–1091. [Google Scholar] [CrossRef]
- Sarma, V.R.; Olotu, F.A.; Soliman, M.E.S. Integrative immunoinformatics paradigm for predicting potential B-cell and T-cell epitopes as viable candidates for subunit vaccine design against COVID-19 virulence. Biomed. J. 2021, 44, 447–460. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Mahajan, S.; Peters, B.; Nielsen, M.; Marcatili, P. Antibody Specific B-Cell Epitope Predictions: Leveraging Information From Antibody-Antigen Protein Complexes. Front. Immunol. 2019, 10, 298. [Google Scholar] [CrossRef]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef] [Green Version]
- Emini, E.A.; Hughes, J.V.; Perlow, D.S.; Boger, J. Induction of hepatitis A virus-neutralizing antibody by a virus-specific synthetic peptide. J. Virol. 1985, 55, 836–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kar, T.; Narsaria, U.; Basak, S.; Deb, D.; Castiglione, F.; Mueller, D.M.; Srivastava, A.P. A candidate multi-epitope vaccine against SARS-CoV-2. Sci. Rep. 2020, 10, 10895. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Chu, Z.; Yu, F.; Zha, Y. Contriving multi-epitope subunit of vaccine for COVID-19: Immunoinformatics approaches. Front. Immunol. 2020, 11, 1784. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Trincado, J.L.; Gomez-Perosanz, M.; Reche, P.A. Fundamentals and methods for T- and B-cell epitope prediction. J. Immunol. Res. 2017, 2017, 2680160. [Google Scholar] [CrossRef] [Green Version]
- Farrera-Soler, L.; Daguer, J.-P.; Barluenga, S.; Vadas, O.; Cohen, P.; Pagano, S.; Yerly, S.; Kaiser, L.; Vuilleumier, N.; Winssinger, N. Identification of immunodominant linear epitopes from SARS-CoV-2 patient plasma. PLoS ONE 2020, 15, e0238089. [Google Scholar] [CrossRef] [PubMed]
- Poh, C.M.; Carissimo, G.; Wang, B.; Amrun, S.N.; Lee, C.Y.; Chee, R.S.; Fong, S.W.; Yeo, N.K.; Lee, W.H.; Torres-Ruesta, A.; et al. Two linear epitopes on the SARS-CoV-2 spike protein that elicit neutralising antibodies in COVID-19 patients. Nat. Commun. 2020, 11, 2806. [Google Scholar] [CrossRef]
- Tian, X.; Li, C.; Huang, A.; Xia, S.; Lu, S.; Shi, Z.; Lu, L.; Jiang, S.; Yang, Z.; Wu, Y.; et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg. Microbes Infect. 2020, 9, 382–385. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Xie, X.X.; Zhao, L.; Wang, B.; Zhu, J.; Yang, T.R.; Yang, G.W.; Ji, M.; Lv, C.P.; Xue, J.; et al. The immunodominant and neutralization linear epitopes for SARS-CoV-2. Cell Rep. 2021, 34, 108666. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, G.; Liu, H.; Ding, P.; Liu, Y.; Tian, Y.; Wang, Y.; Wang, A. Epitope profiling reveals the critical antigenic determinants in SARS-CoV-2 RBD-based antigen. Front. Immunol. 2021, 12, 707977. [Google Scholar] [CrossRef]
- Li, L.; Zhao, Z.; Yang, X.; Li, W.; Chen, S.; Sun, T.; Wang, L.; He, Y.; Liu, G.; Han, X.; et al. Identification of four linear B-cell epitopes on the SARS-CoV-2 spike protein able to elicit neutralizing antibodies. bioRxiv 2020. [Google Scholar] [CrossRef]
- Yi, Z.; Ling, Y.; Zhang, X.; Chen, J.; Hu, K.; Wang, Y.; Song, W.; Ying, T.; Zhang, R.; Lu, H.; et al. Functional mapping of B-cell linear epitopes of SARS-CoV-2 in COVID-19 convalescent population. Emerg. Microbes Infect. 2020, 9, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lai, D.Y.; Zhang, H.N.; Jiang, H.W.; Tian, X.; Ma, M.L.; Qi, H.; Meng, Q.F.; Guo, S.J.; Wu, Y.; et al. Linear epitopes of SARS-CoV-2 spike protein elicit neutralizing antibodies in COVID-19 patients. Cell Mol. Immunol. 2020, 17, 1095–1097. [Google Scholar] [CrossRef] [PubMed]
- Polyiam, K.; Phoolcharoen, W.; Butkhot, N.; Srisaowakarn, C.; Thitithanyanont, A.; Auewarakul, P.; Hoonsuwan, T.; Ruengjitchatchawalya, M.; Mekvichitsaeng, P.; Roshorm, Y.M. Immunodominant linear B cell epitopes in the spike and membrane proteins of SARS-CoV-2 identified by immunoinformatics prediction and immunoassay. Sci. Rep. 2021, 11, 20383. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Wang, A.; Liu, M.; Wang, Q.; Chen, J.; Xia, S.; Ling, Y.; Zhang, Y.; Xun, J.; Lu, L.; et al. Neutralizing antibody responses to SARS-CoV-2 in a COVID-19 recovered patient cohort and their implications. medRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Xing, S.; Ding, L.; Wang, Y.; Gu, C.; Wu, Y.; Rong, B.; Li, C.; Wang, S.; Chen, K.; et al. Human-IgG-neutralizing monoclonal antibodies block the SARS-CoV-2 infection. Cell Rep. 2020, 32, 107918. [Google Scholar] [CrossRef]
- Makdasi, E.; Levy, Y.; Alcalay, R.; Noy-Porat, T.; Zahavy, E.; Mechaly, A.; Epstein, E.; Peretz, E.; Cohen, H.; Bar-On, L.; et al. Neutralizing monoclonal anti-SARS-CoV-2 antibodies isolated from immunized rabbits define novel vulnerable spike-protein epitope. Viruses 2021, 13, 566. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Shen, C.; Peng, W.; Li, D.; Zhao, C.; Li, Z.; Li, S.; Bi, Y.; Yang, Y.; et al. A noncompeting pair of human neutralizing antibodies block COVID-19 virus binding to its receptor ACE2. Science 2020, 368, 1274–1278. [Google Scholar] [CrossRef]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef]
- Noy-Porat, T.; Makdasi, E.; Alcalay, R.; Mechaly, A.; Levy, Y.; Bercovich-Kinori, A.; Zauberman, A.; Tamir, H.; Yahalom-Ronen, Y.; Israeli, M.A.; et al. A panel of human neutralizing mAbs targeting SARS-CoV-2 spike at multiple epitopes. Nat. Commun. 2020, 11, 4303. [Google Scholar] [CrossRef]
- Pinto, D.; Park, Y.J.; Beltramello, M.; Walls, A.C.; Tortorici, M.A.; Bianchi, S.; Jaconi, S.; Culap, K.; Zatta, F.; De Marco, A.; et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 2020, 583, 290–295. [Google Scholar] [CrossRef]
- Brouwer, P.J.M.; Caniels, T.G.; van der Straten, K.; Snitselaar, J.L.; Aldon, Y.; Bangaru, S.; Torres, J.L.; Okba, N.M.A.; Claireaux, M.; Kerster, G.; et al. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science 2020, 369, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.; Sahi, V.; Figueroa, A.; et al. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Sauer, M.M.; Czudnochowski, N.; Low, J.S.; Tortorici, M.A.; Housley, M.P.; Noack, J.; Walls, A.C.; Bowen, J.E.; Guarino, B.; et al. Broad betacoronavirus neutralization by a stem helix-specific human antibody. Science 2021, 373, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Bui, H.H.; Sidney, J.; Li, W.; Fusseder, N.; Sette, A. Development of an epitope conservancy analysis tool to facilitate the design of epitope-based diagnostics and vaccines. BMC Bioinform. 2007, 8, 361. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Wang, J.; Jian, F.; Xiao, T.; Song, W.; Yisimayi, A.; Huang, W.; Li, Q.; Wang, P.; An, R.; et al. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 2021, 602, 657–663. [Google Scholar] [CrossRef]
- Chi, X.; Liu, X.; Wang, C.; Zhang, X.; Li, X.; Hou, J.; Ren, L.; Jin, Q.; Wang, J.; Yang, W. Humanized single domain antibodies neutralize SARS-CoV-2 by targeting the spike receptor binding domain. Nat. Commun. 2020, 11, 4528. [Google Scholar] [CrossRef]
- Chi, X.; Yan, R.; Zhang, J.; Zhang, G.; Zhang, Y.; Hao, M.; Zhang, Z.; Fan, P.; Dong, Y.; Yang, Y.; et al. A neutralizing human antibody binds to the N-terminal domain of the spike protein of SARS-CoV-2. Science 2020, 369, 650–655. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.X.; Masomian, M.; Khalid, K.; Kumar, A.U.; MacAry, P.A.; Poh, C.L. Identification of B-Cell Epitopes for Eliciting Neutralizing Antibodies against the SARS-CoV-2 Spike Protein through Bioinformatics and Monoclonal Antibody Targeting. Int. J. Mol. Sci. 2022, 23, 4341. https://doi.org/10.3390/ijms23084341
Lim HX, Masomian M, Khalid K, Kumar AU, MacAry PA, Poh CL. Identification of B-Cell Epitopes for Eliciting Neutralizing Antibodies against the SARS-CoV-2 Spike Protein through Bioinformatics and Monoclonal Antibody Targeting. International Journal of Molecular Sciences. 2022; 23(8):4341. https://doi.org/10.3390/ijms23084341
Chicago/Turabian StyleLim, Hui Xuan, Malihe Masomian, Kanwal Khalid, Asqwin Uthaya Kumar, Paul A. MacAry, and Chit Laa Poh. 2022. "Identification of B-Cell Epitopes for Eliciting Neutralizing Antibodies against the SARS-CoV-2 Spike Protein through Bioinformatics and Monoclonal Antibody Targeting" International Journal of Molecular Sciences 23, no. 8: 4341. https://doi.org/10.3390/ijms23084341
APA StyleLim, H. X., Masomian, M., Khalid, K., Kumar, A. U., MacAry, P. A., & Poh, C. L. (2022). Identification of B-Cell Epitopes for Eliciting Neutralizing Antibodies against the SARS-CoV-2 Spike Protein through Bioinformatics and Monoclonal Antibody Targeting. International Journal of Molecular Sciences, 23(8), 4341. https://doi.org/10.3390/ijms23084341