
Hippocampal Deletion of CB1 Receptor Impairs Social Memory and Leads to Age-Related Changes in the Hippocampus of Adult Mice
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
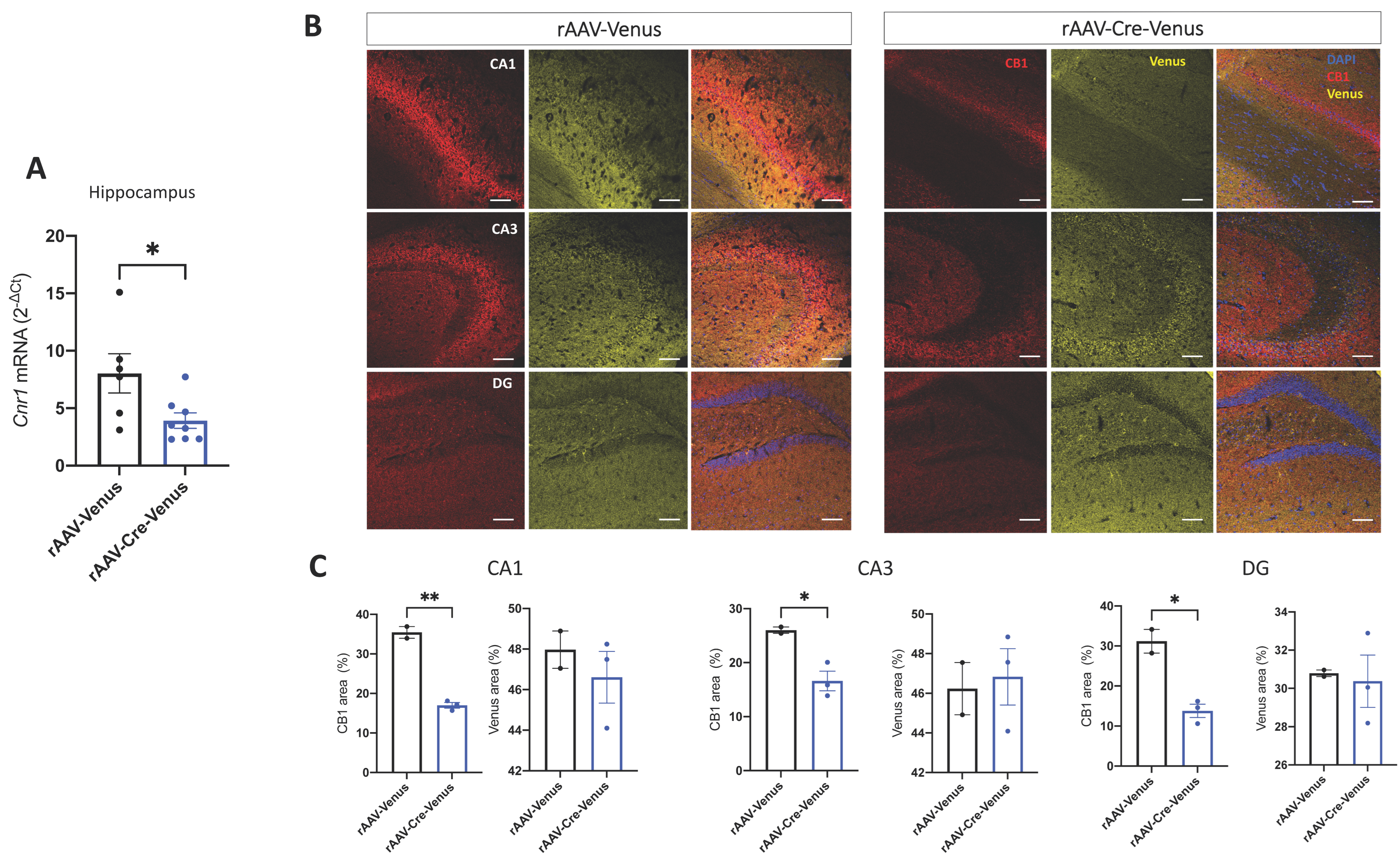
2.1. Validation of Site-Specific Deletion of CB1 in the Hippocampus
2.2. Microglia and Astrocytes Activity in the Hippocampus
2.3. Hippocampal mRNA Expression of Pro-Inflammatory Cytokines
2.4. Cell Proliferation in the Subgranular Zone of DG and in the Subventricular Zone of the Lateral Ventricle
2.5. Neuronal Densities in the Hippocampus
2.6. Evaluation of the Social Memory in the Partner Recognition Test
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Stereotaxic Viral Injections
4.3. Partner Recognition Test
4.4. Real-Time PCR
4.5. Immunohistochemistry
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zia, A.; Pourbagher-Shahri, A.M.; Farkhondeh, T.; Samarghandian, S. Molecular and Cellular Pathways Contributing to Brain Aging. Behav. Brain Funct. 2021, 17, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Glisky, E. Changes in Cognitive Function in Human Aging. In Brain Aging: Models, Methods, and Mechanisms; Riddle, D.R., Ed.; Press/Taylor & Francis: Boca Raton, FL, USA, 2007; pp. 3–20. [Google Scholar]
- Bilkei-Gorzo, A. The Endocannabinoid System in Normal and Pathological Brain Ageing. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3326–3341. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Stella, N.; Zimmer, A. Endocannabinoid Signalling and the Deteriorating Brain. Nat. Rev. Neurosci. 2015, 16, 30–42. [Google Scholar] [CrossRef] [Green Version]
- Piomelli, D. More Surprises Lying Ahead. The Endocannabinoids Keep Us Guessing. Neuropharmacology 2014, 76, 228–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamura, Y.; Fukaya, M.; Maejima, T.; Yoshida, T.; Miura, E.; Watanabe, M.; Ohno-Shosaku, T.; Kano, M. The CB1 Cannabinoid Receptor Is the Major Cannabinoid Receptor at Excitatory Presynaptic Sites in the Hippocampus and Cerebellum. J. Neurosci. 2006, 26, 2991–3001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarrete, M.; Araque, A. Endocannabinoids Mediate Neuron-Astrocyte Communication. Neuron 2008, 57, 883–893. [Google Scholar] [CrossRef] [Green Version]
- Robledo-Menendez, A.; Vella, M.; Grandes, P.; Soria-Gomez, E. Cannabinoid Control of Hippocampal Functions: The Where Matters. FEBS J. 2022, 289, 2162–2175. [Google Scholar] [CrossRef]
- Brunt, T.M.; Bossong, M.G. The Neuropharmacology of Cannabinoid Receptor Ligands in Central Signaling Pathways. Eur. J. Neurosci. 2022, 55, 909–921. [Google Scholar] [CrossRef]
- Rojo-Bustamante, E.; Íñigo-Marco, I.; Abellanas, M.A.; Vinueza-Gavilanes, R.; Baltanás, A.; Luquin, E.; Arrasate, M.; Aymerich, M.S. Cb2 Receptors and Neuron–Glia Interactions Modulate Neurotoxicity Generated by Magl Inhibition. Biomolecules 2020, 10, 1198. [Google Scholar] [CrossRef]
- Marsicano, G.; Lutz, B. Expression of the Cannabinoid Receptor CB1 in Distinct Neuronal Subpopulations in the Adult Mouse Forebrain. Eur. J. Neurosci. 1999, 11, 4213–4225. [Google Scholar] [CrossRef]
- Steindel, F.; Lerner, R.; Häring, M.; Ruehle, S.; Marsicano, G.; Lutz, B.; Monory, K. Neuron-Type Specific Cannabinoid-Mediated G Protein Signalling in Mouse Hippocampus. J. Neurochem. 2013, 124, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Bilkei-Gorzo, A.; Albayram, O.; Ativie, F.; Chasan, S.; Zimmer, T.; Bach, K.; Zimmer, A. Cannabinoid 1 Receptor Signaling on GABAergic Neurons Influences Astrocytes in the Ageing Brain. PLoS ONE 2018, 13, e0202566. [Google Scholar] [CrossRef] [PubMed]
- Stella, N. Cannabinoid and Cannabinoid-like Receptors in Microglia, Astrocytes, and Astrocytomas. Glia 2010, 58, 1017–1030. [Google Scholar] [CrossRef] [Green Version]
- Hernangomez, M.; Carrillo-Salinas, F.; Mecha, M.; Correa, F.; Mestre, L.; Loria, F.; Feliu, A.; Docagne, F.; Guaza, C. Brain Innate Immunity in the Regulation of Neuroinflammation: Therapeutic Strategies by Modulating CD200-CD200R Interaction Involve the Cannabinoid System. Curr. Pharm. Des. 2014, 20, 4707–4722. [Google Scholar] [CrossRef] [Green Version]
- Froger, N.; Orellana, J.A.; Cohen-Salmon, M.; Ezan, P.; Amigou, E.; Sáez, J.C.; Giaume, C. Cannabinoids Prevent the Opposite Regulation of Astroglial Connexin43 Hemichannels and Gap Junction Channels Induced by Pro-Inflammatory Treatments. J. Neurochem. 2009, 111, 1383–1397. [Google Scholar] [CrossRef] [PubMed]
- Mecha, M.; Feliú, A.; Carrillo-Salinas, F.J.; Rueda-Zubiaurre, A.; Ortega-Gutiérrez, S.; de Sola, R.G.; Guaza, C. Endocannabinoids Drive the Acquisition of an Alternative Phenotype in Microglia. Brain. Behav. Immun. 2015, 49, 233–245. [Google Scholar] [CrossRef]
- Duffy, S.S.; Hayes, J.P.; Fiore, N.T.; Moalem-Taylor, G. The Cannabinoid System and Microglia in Health and Disease. Neuropharmacology 2021, 190, 108555. [Google Scholar] [CrossRef]
- Young, A.P.; Denovan-Wright, E.M. The Dynamic Role of Microglia and the Endocannabinoid System in Neuroinflammation. Front. Pharmacol. 2022, 12, 1–16. [Google Scholar] [CrossRef]
- Vasincu, A.; Rusu, R.N.; Ababei, D.C.; Larion, M.; Bild, W.; Stanciu, G.D.; Solcan, C.; Bild, V. Endocannabinoid Modulation in Neurodegenerative Diseases: In Pursuit of Certainty. Biology 2022, 11, 440. [Google Scholar] [CrossRef]
- Ward, R.J.; Dexter, D.T.; Crichton, R.R. Ageing, Neuroinflammation and Neurodegeneration. Front. Biosci.-Schol. Ed. 2015, 7S, 189–204. [Google Scholar] [CrossRef]
- Chevalier-Larsen, E.; Holzbaur, E.L.F. Axonal Transport and Neurodegenerative Disease. Biochim. Biophys. Acta-Mol. Basis Dis. 2006, 1762, 1094–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zhang, J.; Chen, C. Endocannabinoid 2-Arachidonoylglycerol Protects Neuronsagainst β-Amyloid Insults. Neuroscience 2011, 178, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aso, E.; Sánchez-Pla, A.; Vegas-Lozano, E.; Maldonado, R.; Ferrer, I. Cannabis-Based Medicine Reduces Multiple Pathological Processes in AβPP/PS1 Mice. J. Alzheimer’s Dis. 2015, 43, 977–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aso, E.; Andrés-Benito, P.; Carmona, M.; Maldonado, R.; Ferrer, I. Cannabinoid Receptor 2 Participates in Amyloid-β Processing in a Mouse Model of Alzheimer’s Disease but Plays a Minor Role in the Therapeutic Properties of a Cannabis-Based Medicine. J. Alzheimer’s Dis. 2016, 51, 489–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodward, M.R.; Harper, D.G.; Stolyar, A.; Forester, B.P.; Ellison, J.M. Dronabinol for the Treatment of Agitation and Aggressive Behavior in Acutely Hospitalized Severely Demented Patients with Noncognitive Behavioral Symptoms. Am. J. Geriatr. Psychiatry 2014, 22, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, F.; García-Gutiérrez, M.S.; Aracil-Fernández, A.; Lanciego, J.L.; Manzanares, J. Cannabinoid Cb1 and Cb2 Receptors, and Monoacylglycerol Lipase Gene Expression Alterations in the Basal Ganglia of Patients with Parkinson’s Disease. Neurotherapeutics 2018, 15, 459–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peball, M.; Krismer, F.; Knaus, H.G.; Djamshidian, A.; Werkmann, M.; Carbone, F.; Ellmerer, P.; Heim, B.; Marini, K.; Valent, D.; et al. Non-Motor Symptoms in Parkinson’s Disease Are Reduced by Nabilone. Ann. Neurol. 2020, 88, 712–722. [Google Scholar] [CrossRef]
- Chagas, M.H.N.; Zuardi, A.W.; Tumas, V.; Pena-Pereira, M.A.; Sobreira, E.T.; Bergamaschi, M.M.; Dos Santos, A.C.; Teixeira, A.L.; Hallak, J.E.C.; Crippa, J.A.S. Effects of Cannabidiol in the Treatment of Patients with Parkinson’s Disease: An Exploratory Double-Blind Trial. J. Psychopharmacol. 2014, 28, 1088–1092. [Google Scholar] [CrossRef]
- Benito, C.; Romero, J.P.; Tolón, R.M.; Clemente, D.; Docagne, F.; Hillard, C.J.; Guaza, C.; Romero, J. Cannabinoid CB1 and CB2 Receptors and Fatty Acid Amide Hydrolase Are Specific Markers of Plaque Cell Subtypes in Human Multiple Sclerosis. J. Neurosci. 2007, 27, 2396–2402. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, S.; Germanos, R.; Weier, M.; Pollard, J.; Degenhardt, L.; Hall, W.; Buckley, N.; Farrell, M. The Use of Cannabis and Cannabinoids in Treating Symptoms of Multiple Sclerosis: A Systematic Review of Reviews. Curr. Neurol. Neurosci. Rep. 2018, 18. [Google Scholar] [CrossRef]
- Scotter, E.L.; Abood, M.E.; Glass, M. The Endocannabinoid System as a Target for the Treatment of Neurodegenerative Disease. Br. J. Pharmacol. 2010, 160, 480–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacher, P.; Kunos, G. Modulating the Endocannabinoid System in Human Health and Disease—Successes and Failures. FEBS J. 2013, 280, 1918–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skaper, S.D.; Di Marzo, V. Endocannabinoids in Nervous System Health and Disease: The Big Picture in a Nutshell. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3193–3200. [Google Scholar] [CrossRef] [PubMed]
- Nidadavolu, P.; Bilkei-Gorzo, A.; Effah, F.; Leidmaa, E.; Schürmann, B.; Berger, M.; Bindila, L.; Schmid, M.; Lutz, B.; Zimmer, A.; et al. Dynamic Changes in the Endocannabinoid System during the Aging Process: Focus on the Middle-Age Crisis. Int. J. Mol. Sci. 2022, 23, 10254. [Google Scholar] [CrossRef]
- Canas, P.M.; Duarte, J.M.N.; Rodrigues, R.J.; Köfalvi, A.; Cunha, R.A. Modification upon Aging of the Density of Presynaptic Modulation Systems in the Hippocampus. Neurobiol. Aging 2009, 30, 1877–1884. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, J.; Harvey-White, J.; Zimmer, A.; Kunos, G. Endocannabinoid Signaling via Cannabinoid Receptor 1 Is Involved in Ethanol Preference and Its Age-Dependent Decline in Mice. Proc. Natl. Acad. Sci. USA 2003, 100, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Feliszek, M.; Bindila, L.; Lutz, B.; Zimmer, A.; Bilkei-Gorzo, A.; Schlicker, E. Lack of Hippocampal CB1 Receptor Desensitization by Δ9-Tetrahydrocannabinol in Aged Mice and by Low Doses of JZL 184. Naunyn. Schmiedebergs. Arch. Pharmacol. 2016, 389, 603–612. [Google Scholar] [CrossRef]
- Piyanova, A.; Lomazzo, E.; Bindila, L.; Lerner, R.; Albayram, O.; Ruhl, T.; Lutz, B.; Zimmer, A.; Bilkei-Gorzo, A. Age-Related Changes in the Endocannabinoid System in the Mouse Hippocampus. Mech. Ageing Dev. 2015, 150, 55–64. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Compagnucci, C.; Di Siena, S.; Bustamante, M.B.; Di Giacomo, D.; Di Tommaso, M.; Maccarrone, M.; Grimaldi, P.; Sette, C. Type-1 (CB1) Cannabinoid Receptor Promotes Neuronal Differentiation and Maturation of Neural Stem Cells. PLoS ONE 2013, 8, e54271. [Google Scholar] [CrossRef]
- Borro, M.; Frati, P.; Luca, N.; Simmaco, M.; Ferracuti, S. The Cannabinoids Switch of Oxidative Stress: A Double-Edged Sword. Mini. Rev. Org. Chem. 2013, 10, 343–348. [Google Scholar] [CrossRef]
- Idris, A.I.; Sophocleous, A.; Landao-Bassonga, E.; Canals, M.; Milligan, G.; Baker, D.; van’t Hof, R.J.; Ralston, S.H. Cannabinoid Receptor Type 1 Protects against Age- Related Osteoporosis by Regulating Osteoblast and Adipocyte Differentiation in Marrow Stromal Cells. Cell Metab. 2009, 10, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilkei-Gorzo, A.; Racz, I.; Valverde, O.; Otto, M.; Michel, K.; Sarstre, M.; Zimmer, A. Early Age-Related Cognitive Impairment in Mice Lacking Cannabinoid CB1 Receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 15670–15675. [Google Scholar] [CrossRef] [Green Version]
- Albayram, O.; Bilkei-Gorzo, A.; Zimmer, A. Loss of CB1 Receptors Leads to Differential Age-Related Changes in Reward-Driven Learning and Memory. Front. Aging Neurosci. 2012, 4, 34. [Google Scholar] [CrossRef] [Green Version]
- Piyanova, A.; Albayram, O.; Rossi, C.A.; Farwanah, H.; Michel, K.; Nicotera, P.; Sandhoff, K.; Bilkei-Gorzo, A. Loss of CB1 Receptors Leads to Decreased Cathepsin D Levels and Accelerated Lipofuscin Accumulation in the Hippocampus. Mech. Ageing Dev. 2013, 134, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Albayram, O.; Alferink, J.; Pitsch, J.; Piyanova, A.; Neitzert, K.; Poppensieker, K.; Mauer, D.; Michel, K.; Legler, A.; Becker, A.; et al. Role of CB1 Cannabinoid Receptors on GABAergic Neurons in Brain Aging. Proc. Natl. Acad. Sci. USA 2011, 108, 11256–11261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, W.S.; Hu, S.; Min, X.; Cabral, G.A.; Lokensgard, J.R.; Peterson, P.K. Synthetic Cannabinoid WIN55,212-2 Inhibits Generation of Inflammatory Mediators by IL-1β-Stimulated Human Astrocytes. Glia 2005, 49, 211–219. [Google Scholar] [CrossRef]
- Michaud, M.; Balardy, L.; Moulis, G.; Gaudin, C.; Peyrot, C.; Vellas, B.; Cesari, M.; Nourhashemi, F. Proinflammatory Cytokines, Aging, and Age-Related Diseases. J. Am. Med. Dir. Assoc. 2013, 14, 877–882. [Google Scholar] [CrossRef]
- Bilkei-Gorzo, A.; Albayram, O.; Draffehn, A.; Michel, K.; Piyanova, A.; Oppenheimer, H.; Dvir-Ginzberg, M.; Rácz, I.; Ulas, T.; Imbeault, S.; et al. A Chronic Low Dose of Δ9-Tetrahydrocannabinol (THC) Restores Cognitive Function in Old Mice. Nat. Med. 2017, 23, 782–787. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Gómez-Ruiz, M.; García, C.; Hernández, M.; Ramos, J.A. Modeling Neurodegenerative Disorders for Developing Cannabinoid-Based Neuroprotective Therapies. Methods Enzymol. 2017, 593, 175–198. [Google Scholar] [CrossRef]
- Zamberletti, E.; Rubino, T. Dos(e)Age: Role of Dose and Age in the Long-Term Effect of Cannabinoids on Cognition. Molecules 2022, 27, 1411. [Google Scholar] [CrossRef] [PubMed]
- Bilkei-Gorzo, A.; Drews, E.; Albayram, Ö.; Piyanova, A.; Gaffal, E.; Tueting, T.; Michel, K.; Mauer, D.; Maier, W.; Zimmer, A. Early Onset of Aging-like Changes Is Restricted to Cognitive Abilities and Skin Structure in Cnr1-/- Mice. Neurobiol. Aging 2012, 33, 200.e11–200.e22. [Google Scholar] [CrossRef] [PubMed]
- Reibaud, M.; Obinu, M.C.; Ledent, C.; Parmentier, M.; Böhme, G.A.; Imperato, A. Enhancement of Memory in Cannabinoid CB1 Receptor Knock-out Mice. Eur. J. Pharmacol. 1999, 379, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Jeon, O.H.; Kim, C.; Laberge, R.M.; Demaria, M.; Rathod, S.; Vasserot, A.P.; Chung, J.W.; Kim, D.H.; Poon, Y.; David, N.; et al. Local Clearance of Senescent Cells Attenuates the Development of Post-Traumatic Osteoarthritis and Creates a pro-Regenerative Environment. Nat. Med. 2017, 23, 775–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentijn, F.A.; Falke, L.L.; Nguyen, T.Q.; Goldschmeding, R. Cellular Senescence in the Aging and Diseased Kidney. J. Cell Commun. Signal. 2018, 12, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.W.; Wang, W.-T.; Chou, L.C.; Liao, C.-D.; Liou, T.H.; Lin, H.W. Osteoarthritis Increases the Risk of Dementia: A Nationwide Cohort Study in Taiwan. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Deckers, K.; Camerino, I.; van Boxtel, M.P.J.; Verhey, F.R.J.; Irving, K.; Brayne, C.; Kivipelto, M.; Starr, J.M.; Yaffe, K.; de Leeuw, P.W.; et al. Dementia Risk in Renal Dysfunction. Neurology 2017, 88, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Villeda, S.A.; Luo, J.; Mosher, K.I.; Zou, B.; Britschgi, M.; Bieri, G.; Stan, T.M.; Fainberg, N.; Ding, Z.; Eggel, A.; et al. The Ageing Systemic Milieu Negatively Regulates Neurogenesis and Cognitive Function. Nature 2011, 477, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Villeda, S.A.; Plambeck, K.E.; Middeldorp, J.; Castellano, J.M.; Mosher, K.I.; Luo, J.; Smith, L.K.; Bieri, G.; Lin, K.; Berdnik, D.; et al. Young Blood Reverses Age-Related Impairments in Cognitive Function and Synaptic Plasticity in Mice. Nat. Med. 2014, 20, 659–663. [Google Scholar] [CrossRef] [Green Version]
- Trezza, V.; Damsteegt, R.; Manduca, A.; Petrosino, S.; van Kerkhof, L.W.M.; Jeroen Pasterkamp, R.; Zhou, Y.; Campolongo, P.; Cuomo, V.; Di Marzo, V.; et al. Endocannabinoids in Amygdala and Nucleus Accumbens Mediate Social Play Reward in Adolescent Rats. J. Neurosci. 2012, 32, 14899–14908. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Lee, D.Y.; Li, D.; Daglian, J.; Jung, K.M.; Piomelli, D. A Role for the Endocannabinoid 2-Arachidonoyl-Sn-Glycerol for Social and High-Fat Food Reward in Male Mice. Psychopharmacology 2016, 233, 1911–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loureiro, M.; Kramar, C.; Renard, J.; Rosen, L.G.; Laviolette, S.R. Cannabinoid Transmission in the Hippocampus Activates Nucleus Accumbens Neurons and Modulates Reward and Aversion-Related Emotional Salience. Biol. Psychiatry 2016, 80, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Pocock, J.M.; Kettenmann, H. Neurotransmitter Receptors on Microglia. Trends Neurosci. 2007, 30, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Alain, B.; Béchade, C.; Bernard, D.; Roumier, A. Microglial Control of Neuronal Death and Synaptic Properties. Glia 2007, 55, 233–238. [Google Scholar] [CrossRef]
- Llorente-Ovejero, A.; Bengoetxea De Tena, I.; Martínez-Gardeazabal, J.; Moreno-Rodríguez, M.; Lombardero, L.; Manuel, I.; Rodríguez-Puertas, R. Cannabinoid Receptors and Glial Response Following a Basal Forebrain Cholinergic Lesion. ACS Pharmacol. Transl. Sci. 2022, 5, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Lou, Z.Y.; Cheng, J.; Wang, X.R.; Zhao, Y.F.; Gan, J.; Zhou, G.Y.; Liu, Z.G.; Xiao, B.G. The Inhibition of CB1 Receptor Accelerates the Onset and Development of EAE Possibly by Regulating Microglia/Macrophages Polarization. J. Neuroimmunol. 2018, 317, 37–44. [Google Scholar] [CrossRef]
- Wolf, S.; Tauber, S.; Ullrich, O. CNS Immune Surveillance and Neuroinflammation: Endocannabinoids Keep Control. Curr. Pharm. Des. 2008, 14, 2266–2278. [Google Scholar] [CrossRef]
- Egaña-Huguet, J.; Soria-Gómez, E.; Grandes, P. The Endocannabinoid System in Glial Cells and Their Profitable Interactions to Treat Epilepsy: Evidence from Animal Models. Int. J. Mol. Sci. 2021, 22, 13231. [Google Scholar] [CrossRef]
- Cutando, L.; Busquets-Garcia, A.; Puighermanal, E.; Gomis-González, M.; Delgado-García, J.M.; Gruart, A.; Maldonado, R.; Ozaita, A. Microglial Activation Underlies Cerebellar Deficits Produced by Repeated Cannabis Exposure. J. Clin. Invest. 2013, 123, 2816–2831. [Google Scholar] [CrossRef] [Green Version]
- De Meij, J.; Alfanek, Z.; Morel, L.; Decoeur, F.; Leyrolle, Q.; Picard, K.; Carrier, M.; Aubert, M.; Séré, A.; Lucas, C.; et al. Microglial Cannabinoid Type 1 Receptor Regulates Brain Inflammation in a Sex-Specific Manner. Cannabis Cannabinoid Res. 2021, 6, 488–507. [Google Scholar] [CrossRef]
- Ativie, F.; Komorowska, J.A.; Beins, E.; Albayram, Ö.; Zimmer, T.; Zimmer, A.; Tejera, D.; Heneka, M.; Bilkei-Gorzo, A. Cannabinoid 1 Receptor Signaling on Hippocampal GABAergic Neurons Influences Microglial Activity. Front. Mol. Neurosci. 2018, 11, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasatkina, L.A.; Rittchen, S.; Sturm, E.M. Neuroprotective and Immunomodulatory Action of the Endocannabinoid System under Neuroinflammation. Int. J. Mol. Sci. 2021, 22, 5431. [Google Scholar] [CrossRef] [PubMed]
- Milano, W.; Capasso, A. Neuroprotection by Cannabinoids in Neurodegenerative Diseases. Alzheimer’s Dement. Cogn. Neurol. 2018, 2. [Google Scholar] [CrossRef] [Green Version]
- Stella, N.; Piomelli, D. Receptor-Dependent Formation of Endogenous Cannabinoids in Cortical Neurons. Eur. J. Pharmacol. 2001, 425, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Umpierre, A.D.; Wu, L.J. How Microglia Sense and Regulate Neuronal Activity. Glia 2021, 69, 1637–1653. [Google Scholar] [CrossRef]
- Franklin, A.; Parmentier-Batteur, S.; Walter, L.; Greenberg, D.A.; Stella, N. Palmitoylethanolamide Increases after Focal Cerebral Ischemia and Potentiates Microglial Cell Motility. J. Neurosci. 2003, 23, 7767–7775. [Google Scholar] [CrossRef] [Green Version]
- Prenderville, J.A.; Kelly, Á.M.; Downer, E.J. The Role of Cannabinoids in Adult Neurogenesis. Br. J. Pharmacol. 2015, 172, 3950–3963. [Google Scholar] [CrossRef] [Green Version]
- Aguado, T.; Monory, K.; Palazuelos, J.; Stella, N.; Cravatt, B.; Lutz, B.; Marsicano, G.; Kokaia, Z.; Guzmán, M.; Galve-roperh, I. The Endocannabinoid System Drives Neural Progenitor Proliferation. FASEB J. 2005, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, T.; Maroso, M.; Beer, A.; Baddenhausen, S.; Ludewig, S.; Fan, W.; Vennin, C.; Loch, S.; Berninger, B.; Hofmann, C.; et al. Neural Stem Cell Lineage-Specific Cannabinoid Type-1 Receptor Regulates Neurogenesis and Plasticity in the Adult Mouse Hippocampus. Cereb. Cortex 2018, 28, 4454–4471. [Google Scholar] [CrossRef]
- Jenniches, I.; Ternes, S.; Albayram, O.; Otte, D.M.; Bach, K.; Bindila, L.; Michel, K.; Lutz, B.; Bilkei-Gorzo, A.; Zimmer, A. Anxiety, Stress, and Fear Response in Mice with Reduced Endocannabinoid Levels. Biol. Psychiatry 2016, 79, 858–868. [Google Scholar] [CrossRef]
- Schuele, L.L.; Schuermann, B.; Bilkei-Gorzo, A.; Gorgzadeh, S.; Zimmer, A.; Leidmaa, E. Regulation of Adult Neurogenesis by the Endocannabinoid-Producing Enzyme Diacylglycerol Lipase Alpha (DAGLa). Sci. Rep. 2022, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Thiemann, G.; Watt, C.A.; Ledent, C.; Molleman, A.; Hasenöhrl, R.U. Modulation of Anxiety by Acute Blockade and Genetic Deletion of the CB1 Cannabinoid Receptor in Mice Together with Biogenic Amine Changes in the Forebrain. Behav. Brain Res. 2009, 200, 60–67. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmisano, M.; Gargano, A.; Olabiyi, B.F.; Lutz, B.; Bilkei-Gorzo, A. Hippocampal Deletion of CB1 Receptor Impairs Social Memory and Leads to Age-Related Changes in the Hippocampus of Adult Mice. Int. J. Mol. Sci. 2023, 24, 26. https://doi.org/10.3390/ijms24010026
Palmisano M, Gargano A, Olabiyi BF, Lutz B, Bilkei-Gorzo A. Hippocampal Deletion of CB1 Receptor Impairs Social Memory and Leads to Age-Related Changes in the Hippocampus of Adult Mice. International Journal of Molecular Sciences. 2023; 24(1):26. https://doi.org/10.3390/ijms24010026
Chicago/Turabian StylePalmisano, Michela, Alessandra Gargano, Bolanle Fatimat Olabiyi, Beat Lutz, and Andras Bilkei-Gorzo. 2023. "Hippocampal Deletion of CB1 Receptor Impairs Social Memory and Leads to Age-Related Changes in the Hippocampus of Adult Mice" International Journal of Molecular Sciences 24, no. 1: 26. https://doi.org/10.3390/ijms24010026
APA StylePalmisano, M., Gargano, A., Olabiyi, B. F., Lutz, B., & Bilkei-Gorzo, A. (2023). Hippocampal Deletion of CB1 Receptor Impairs Social Memory and Leads to Age-Related Changes in the Hippocampus of Adult Mice. International Journal of Molecular Sciences, 24(1), 26. https://doi.org/10.3390/ijms24010026