
Microarray Expression Profile of Myricetin-Treated THP-1 Macrophages Exhibits Alterations in Atherosclerosis-Related Regulator Molecules and LXR/RXR Pathway

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Myricetin Exhibited No Significant Change to the Viability and Proliferation of THP-1 Macrophages
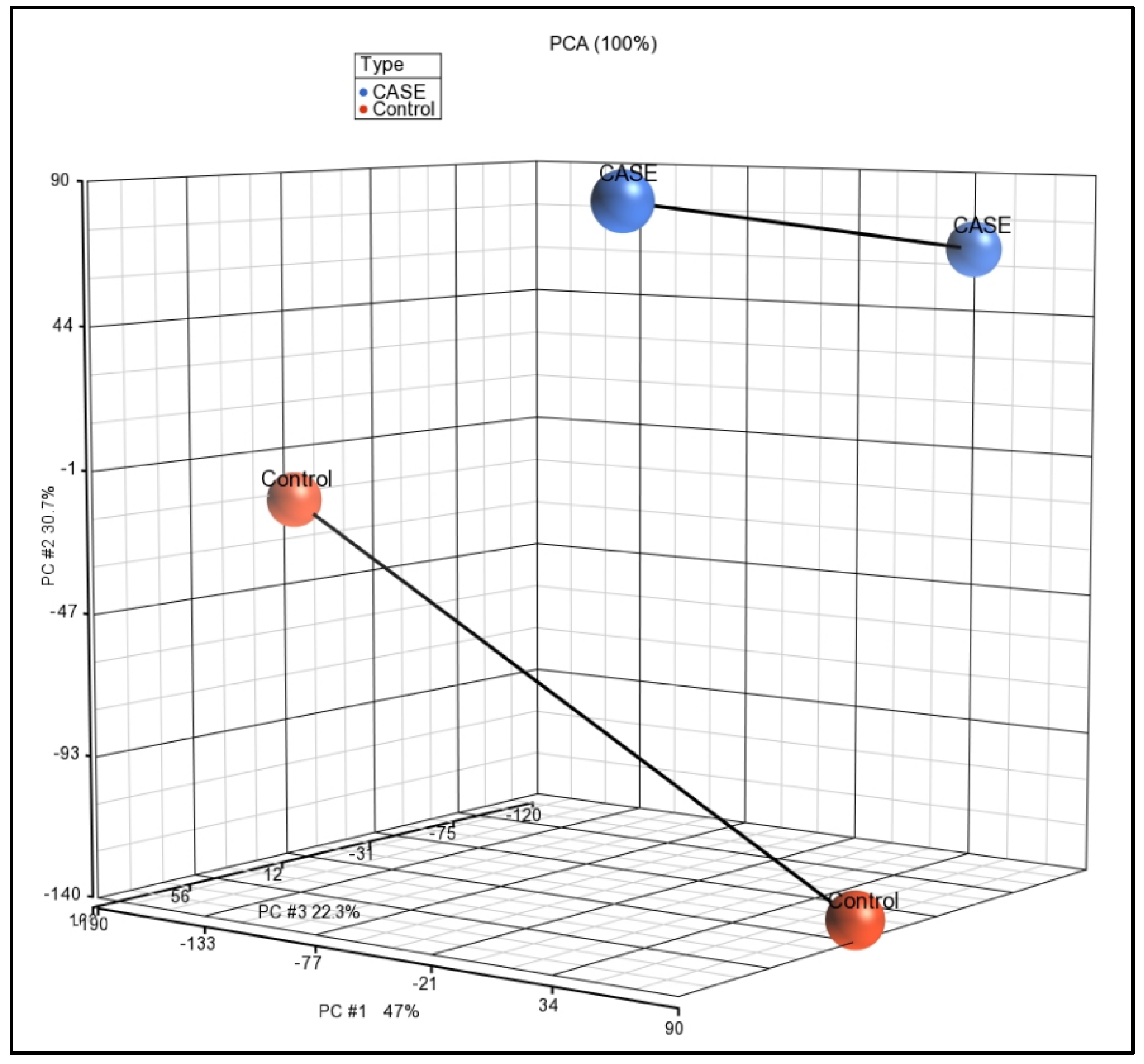
2.2. Gene Expression Profiling of Myricetin-Treated THP-1 Cells
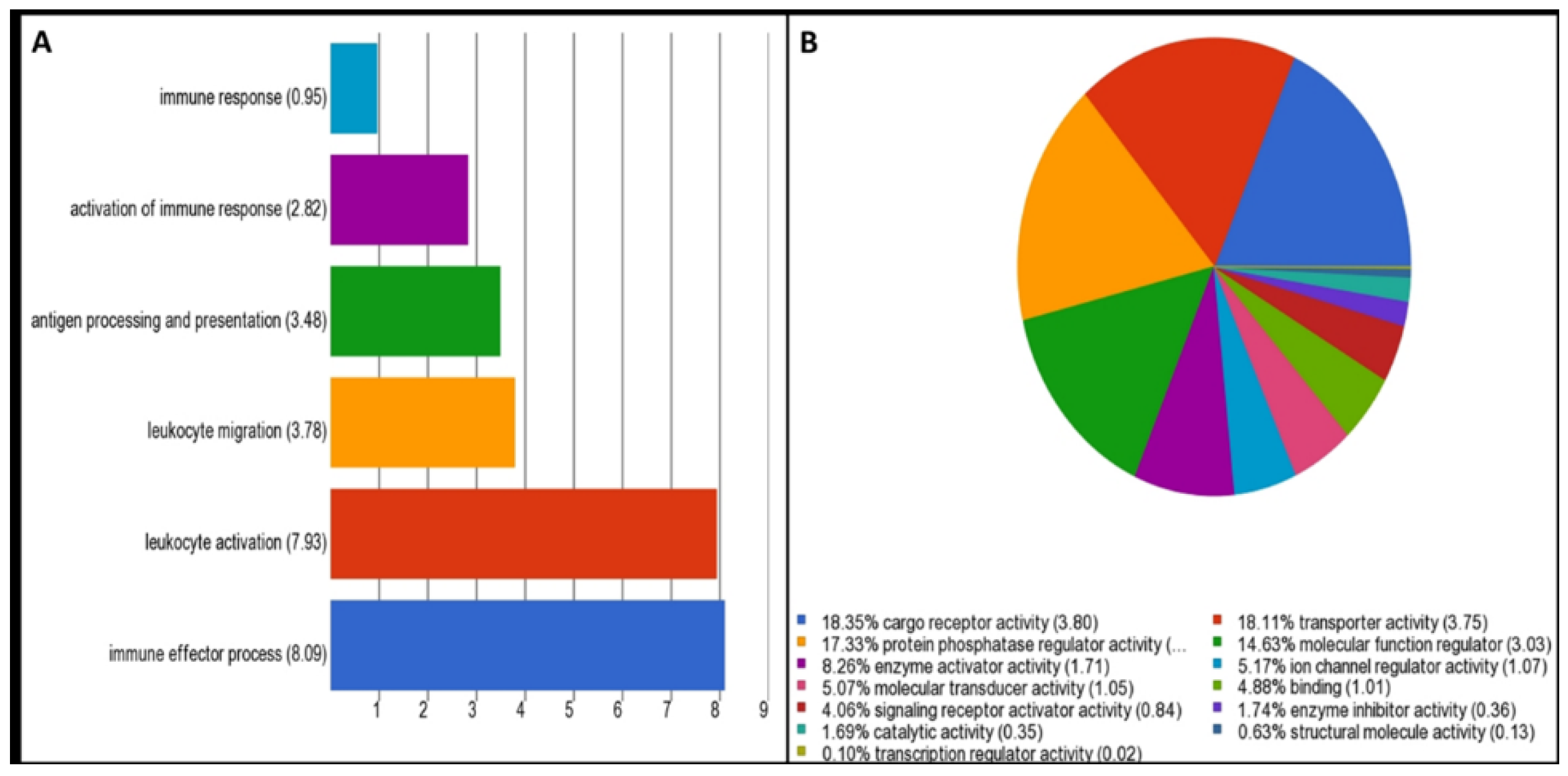
2.3. Gene Ontology, Molecular Pathway, and Network Analysis
2.4. Validation of the DNA Microarray by Using Real-Time Quantitative Polymerase Chain Reaction (q-PCR)
3. Discussion
4. Materials and Methods
4.1. Chemical Materials and Culture Techniques
4.2. Cell Viability and Proliferation Assay
4.3. Migration Study
4.4. Cells Treatment for Microarray Experiment
4.5. Array Processing
4.6. Gene Expression Analysis
4.7. Gene Ontology and Molecular Pathway Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, J.W.; Giannarelli, C.; Rahman, A.; Randolph, G.J.; Kovacic, J.C. Macrophage Biology, Classification, and Phenotype in Cardiovascular Disease: JACC Macrophage in CVD Series (Part 1). J. Am. Coll. Cardiol. 2018, 72, 2166–2180. [Google Scholar] [CrossRef] [PubMed]
- Luque, M.C.A.; Galuppo, M.K.; Capelli-Peixoto, J.; Stolf, B.S. CD100 Effects in Macrophages and Its Roles in Atherosclerosis. Front. Cardiovasc. Med. 2018, 5, 136. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.J. Macrophages in Atherosclerosis Regression. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 20–33. [Google Scholar] [CrossRef]
- Nascimento, C.R.; Rodrigues Fernandes, N.A.; Gonzalez Maldonado, L.A.; Rossa Junior, C. Comparison of monocytic cell lines U937 and THP-1 as macrophage models for in vitro studies. Biochem. Biophys. Rep. 2022, 32, 101383. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Mishra, T.K.; Routray, S. Current perspectives on statins. J. Indian Med. Assoc. 2003, 101, 381–383. [Google Scholar]
- Salvamani, S.; Gunasekaran, B.; Shaharuddin, N.A.; Ahmad, S.A.; Shukor, M.Y. Antiartherosclerotic Effects of Plant Flavonoids. BioMed Res. Int. 2014, 2014, 480258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleemann, R.; Verschuren, L.; Morrison, M.; Zadelaar, S.; van Erk, M.J.; Wielinga, P.Y.; Kooistra, T. Anti-inflammatory, anti-proliferative and anti-atherosclerotic effects of quercetin in human in vitro and in vivo models. Atherosclerosis 2011, 218, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Andarwulan, N.; Batari, R.; Sandrasari, D.A.; Bolling, B.; Wijaya, H. Flavonoid content and antioxidant activity of vegetables from Indonesia. Food Chem. 2010, 121, 1231–1235. [Google Scholar] [CrossRef] [Green Version]
- Ross, J.A.; Kasum, C.M. Dietary Flavonoids: Bioavailability, Metabolic Effects, and Safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Moss, J.W.E.; Williams, J.O.; Ramji, D.P. Nutraceuticals as therapeutic agents for atherosclerosis. Biochim. Et Biophys. Acta Mol. Basis Dis. 2018, 1864, 1562–1572. [Google Scholar] [CrossRef] [PubMed]
- Semwal, D.K.; Semwal, R.B.; Combrinck, S.; Viljoen, A. Myricetin: A Dietary Molecule with Diverse Biological Activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulvihill, E.E.; Burke, A.C.; Huff, M.W. Citrus Flavonoids as Regulators of Lipoprotein Metabolism and Atherosclerosis. Annu. Rev. Nutr. 2016, 36, 275–299. [Google Scholar] [CrossRef]
- Wang, Z.H.; Ah Kang, K.; Zhang, R.; Piao, M.J.; Jo, S.H.; Kim, J.S.; Kang, S.S.; Lee, J.S.; Park, D.H.; Hyun, J.W. Myricetin suppresses oxidative stress-induced cell damage via both direct and indirect antioxidant action. Environ. Toxicol. Pharmacol. 2010, 29, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, G.; Khanna, A.K.; Sonkar, R.; Mishra, S.K.; Srivastava, S.; Lakshmi, V. Lipid lowering and antioxidant activity of flavones in triton treated hyperlipidemic rats. Med. Chem. Res. 2011, 20, 1622–1626. [Google Scholar] [CrossRef]
- Cho, B.O.; Yin, H.H.; Park, S.H.; Byun, E.B.; Ha, H.Y.; Jang, S.I. Anti-inflammatory activity of myricetin from Diospyros lotus through suppression of NF-κB and STAT1 activation and Nrf2-mediated HO-1 induction in lipopolysaccharide-stimulated RAW264.7 macrophages. Biosci. Biotechnol. Biochem. 2016, 80, 1520–1530. [Google Scholar] [CrossRef] [Green Version]
- Meng, Z.; Wang, M.; Xing, J.; Liu, Y.; Li, H. Myricetin ameliorates atherosclerosis in the low-density-lipoprotein receptor knockout mice by suppression of cholesterol accumulation in macrophage foam cells. Nutr. Metab. 2019, 16, 25. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.; Hu, S.; Su, Z.; Wang, Q.; Meng, G.; Guo, T.; Zhang, J.; Gao, P. Myricetin attenuates LPS-induced inflammation in RAW 264.7 macrophages and mouse models. Future Med. Chem. 2018, 10, 2253–2264. [Google Scholar] [CrossRef]
- Hannon, D.B.; Thompson, J.T.; Khoo, C.; Juturu, V.; Vanden Heuvel, J.P. Effects of cranberry extracts on gene expression in THP-1 cells. Food Sci. Nutr. 2016, 5, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Agraharam, G.; Girigoswami, A.; Girigoswami, K. Myricetin: A Multifunctional Flavonol in Biomedicine. Curr. Pharmacol. Rep. 2022, 8, 48–61. [Google Scholar] [CrossRef]
- Kim, J.D.; Liu, L.; Guo, W.; Meydani, M. Chemical structure of flavonols in relation to modulation of angiogenesis and immune-endothelial cell adhesion. J. Nutr. Biochem. 2006, 17, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Ignatova, I.D.; Angdisen, J.; Moran, E.; Schulman, I.G. Differential Regulation of Gene Expression by LXRs in Response to Macrophage Cholesterol Loading. Mol. Endocrinol. 2013, 27, 1036–1047. [Google Scholar] [CrossRef] [Green Version]
- Remmerie, A.; Scott, C.L. Macrophages and lipid metabolism. Cell. Immunol. 2018, 330, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Saenz, J.; Santa-María, C.; Reyes-Quiroz, M.E.; Geniz, I.; Jiménez, J.; Sobrino, F.; Alba, G. Grapefruit Flavonoid Naringenin Regulates the Expression of LXRα in THP-1 Macrophages by Modulating AMP-Activated Protein Kinase. Mol. Pharm. 2018, 15, 1735–1745. [Google Scholar] [CrossRef]
- Cuthbert, G.A.; Shaik, F.; Harrison, M.A.; Ponnambalam, S.; Homer-Vanniasinkam, S. Scavenger Receptors as Biomarkers and Therapeutic Targets in Cardiovascular Disease. Cells 2020, 9, 2453. [Google Scholar] [CrossRef]
- Park, Y.M. CD36, a scavenger receptor implicated in atherosclerosis. Exp. Mol. Med. 2014, 46, e99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, M.W. Scavenger receptors in atherosclerosis. Curr. Opin. Hematol. 1997, 4, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, B.; Volpert, O.V.; Crawford, S.E.; Febbraio, M.; Silverstein, R.L.; Bouck, N. Signals leading to apoptosis-dependent inhibition of neovascularization by thrombospondin-1. Nat. Med. 2000, 6, 41–48. [Google Scholar] [CrossRef]
- Pennathur, S.; Pasichnyk, K.; Bahrami, N.M.; Zeng, L.; Febbraio, M.; Yamaguchi, I.; Okamura, D.M. The macrophage phagocytic receptor CD36 promotes fibrogenic pathways on removal of apoptotic cells during chronic kidney injury. Am. J. Pathol. 2015, 185, 2232–2245. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Varghese, Z.; Moorhead, J.F.; Chen, Y.; Ruan, X.Z. CD36 and lipid metabolism in the evolution of atherosclerosis. Br. Med. Bull. 2018, 126, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Lopez, R.M.; Llorente-Cortes, V.; López-Carmona, D.; Mayas, D.M.; Gomez-Huelgas, R.; Tinahones, F.J.; Badimon, L. Modulation of human monocyte CD36 by type 2 diabetes mellitus and other atherosclerotic risk factors. Eur. J. Clin. Investig. 2011, 41, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Carmona, M.D.; Plaza-Seron, M.C.; Vargas-Candela, A.; Tinahones, F.J.; Gomez-Huelgas, R.; Bernal-Lopez, M.R. CD36 overexpression: A possible etiopathogenic mechanism of atherosclerosis in patients with prediabetes and diabetes. Diabetol. Metab. Syndr. 2017, 9, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, H.Y.; Pearce, S.F.; Yesner, L.M.; Schindler, J.L.; Silverstein, R.L. Regulated expression of CD36 during monocyte-to-macrophage differentiation: Potential role of CD36 in foam cell formation. Blood 1996, 87, 2020–2028. [Google Scholar] [CrossRef] [Green Version]
- Seimon, T.A.; Obstfeld, A.; Moore, K.J.; Golenbock, D.T.; Tabas, I. Combinatorial pattern recognition receptor signaling alters the balance of life and death in macrophages. Proc. Natl. Acad. Sci. USA 2006, 103, 19794–19799. [Google Scholar] [CrossRef] [Green Version]
- Seimon, T.; Tabas, I. Mechanisms and consequences of macrophage apoptosis in atherosclerosis. J. Lipid Res. 2009, 50, S382–S387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, T.W.; Wang, L.; Lo, Y.H.; Huang, I.J.; Wu, M.J. Fisetin, morin and myricetin attenuate CD36 expression and oxLDL uptake in U937-derived macrophages. Biochim. Et Biophys. Acta 2008, 1781, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.D. Myricetin Inhibits Angiogenesis by Inducing Apoptosis and Suppressing PI3K/Akt/mTOR Signaling in Endothelial Cells. J. Cancer Prev. 2017, 22, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Yao, H.; Sun, Z.; Wang, Y.; Zhao, Y.; Wang, Z.; Li, L. Role of NFAT in the Progression of Diabetic Atherosclerosis. Front. Cardiovasc. Med. 2021, 8, 635172. [Google Scholar] [CrossRef]
- Molkentin, J.D. Calcineurin–NFAT signaling regulates the cardiac hypertrophic response in coordination with the MAPKs. Cardiovasc. Res. 2004, 63, 467–475. [Google Scholar] [CrossRef]
- Fortelny, N.; Overall, C.M.; Pavlidis, P.; Freue, G.V.C. Can we predict protein from mRNA levels? Nature 2017, 547, E19–E20. [Google Scholar] [CrossRef]
- Guo, Y.; Xiao, P.; Lei, S.; Deng, F.; Xiao, G.G.; Liu, Y.; Chen, X.; Li, L.; Wu, S.; Chen, Y.; et al. How is mRNA expression predictive for protein expression? A correlation study on human circulating monocytes. Acta Biochim. Et Biophys. Sin. 2008, 40, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Perl, K.; Ushakov, K.; Pozniak, Y.; Yizhar-Barnea, O.; Bhonker, Y.; Shivatzki, S.; Geiger, T.; Avraham, K.B.; Shamir, R. Reduced changes in protein compared to mRNA levels across non-proliferating tissues. BMC Genom. 2017, 18, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almassabi, R.F.; Huwait, E.A.; Almowallad, S.J.; Saddeek, S.Y.; Gauthaman, K. In Vitro: The Modulating Effect of Myricetin on the Atherosclerosis Related Processes in THP1 Macrophages. J. Pharm. Res. Int. 2021, 32, 62–73. [Google Scholar] [CrossRef]
- Moorthy, A.; Venugopal, D.C.; Shyamsundar, V.; Madhavan, Y.; Ravindran, S.; Kuppuloganathan, M.; Krishnamurthy, A.; Sankarapandian, S.; Ganapathy, V.; Ramshankar, V. Identification of EGFR as a Biomarker in Saliva and Buccal Cells from Oral Submucous Fibrosis Patients-A Baseline Study. Diagnostics 2022, 12, 1935. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene_Assignment | Gene_Symbol | p-Value | Fold-Change |
|---|---|---|---|
| NM_002445//macrophage scavenger receptor 1//8p22 | MSR1 | 0.044 | 8.80 |
| NM_000072//CD36 molecule (thrombospondin receptor)//7q11.2 | CD36 | 0.003 | 8.30 |
| NM_001293298//cell migration inducing protein, hyaluronan binding//15q24 | CEMIP | 0.036 | 7.01 |
| NM_001130101//nuclear receptor subfamily 1, group H, member 3//11p11.2 // | NR1H3 | 0.024 | 6.67 |
| NM_001001924//microtubule associated tumor suppressor 1//8p22 | MTUS1 | 0.001 | 6.35 |
| NM_145244//DNA-damage-inducible transcript 4-like//4q24 | DDIT4L | 0.005 | 4.78 |
| NM_001130455//dysferlin//2p13.3 | DYSF | 0.049 | 4.31 |
| NM_001306129//fibronectin 1//2q34 | FN1 | 0.016 | 3.98 |
| NM_001304441//matrix metallopeptidase 8//11q22.3 | MMP8 | 0.021 | 3.97 |
| NM_001444//fatty acid binding protein 5//8q21.13 | FABP5 | 0.027 | 3.91 |
| NM_001122//perilipin 2//9p22.1 | PLIN2 | 0.019 | 3.79 |
| NM_000582//secreted phosphoprotein 1//4q22.1 | SPP1 | 0.042 | 3.71 |
| BT007449//fatty acid binding protein 5 (psoriasis-associated)//8q21.13 // | FABP5 | 0.030 | 3.46 |
| NM_006207//platelet-derived growth factor receptor-like//8p22-p21.3 | PDGFRL | 0.007 | 3.46 |
| NM_001286449//TEX9//testis expressed 9//15q21.3 | TEX9 | 0.004 | 3.44 |
| NR_002324//small nucleolar RNA, H/ACA box 62//3p22.1 | SNORA62 | 0.031 | −3.27 |
| NM_001136219//Fc fragment of IgG, low affinity IIa, receptor (CD32)//1q23 | FCGR2A | 0.037 | −3.18 |
| NM_014278//heat shock 70kDa protein 4-like//4q28 | HSPA4L | 0.011 | −2.58 |
| NM_001271594//SESN3//sestrin 3//11q21 | SESN3 | 0.026 | −2.27 |
| NR_002590//SNORA41//small nucleolar RNA, H/ACA box 41//2q33 | SNORA41 | 0.001 | −2.20 |
| Ingenuity Canonical Pathways | −log(p-Value) | Z-Score | Genes |
|---|---|---|---|
| LXR/RXR Activation | 4.62 | 2.3 | CD36, MSR1, NR1H3 |
| VDR/RXR Activation | 4.46 | 2.13 | IGFBP3, PRKCH, SPP1, TGFB2 |
| Inhibition of Angiogenesis by TSP1 | 3.71 | 2.21 | CD36, SDC2 |
| Phagosome Formation | 3.69 | 2.121 | C5AR2, CD209, CD36, FCGR2A, FN1, ITGA3, MSR1, PRKCH |
| Tumor Microenvironment Pathway | 3.08 | 2.03 | FN1, MMP8, SPP1, TGFB2 |
| Cardiac Hypertrophy Signaling (Enhanced) | 3.05 | 2.236 | ADCY8, ITGA3, PRKCH, RCAN1, TGFB2 |
| Hepatic Cholestasis | 2.97 | 2.107 | ADCY8, NR1H3, PRKCH, TGFB2 |
| RAR Activation | 2.87 | 2.06 | ADCY8, IGFBP3, PRKCH, TGFB2 |
| Role of NFAT in Cardiac Hypertrophy | 2.73 | 2.04 | ADCY8, PRKCH, RCAN1, TGFB2 |
| Wound Healing Signaling Pathway | 2.53 | 2.008 | FN1, ITGA3, MMP8, TGFB2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huwait, E.; Almassabi, R.; Almowallad, S.; Saddeek, S.; Karim, S.; Kalamegam, G.; Mirza, Z. Microarray Expression Profile of Myricetin-Treated THP-1 Macrophages Exhibits Alterations in Atherosclerosis-Related Regulator Molecules and LXR/RXR Pathway. Int. J. Mol. Sci. 2023, 24, 278. https://doi.org/10.3390/ijms24010278
Huwait E, Almassabi R, Almowallad S, Saddeek S, Karim S, Kalamegam G, Mirza Z. Microarray Expression Profile of Myricetin-Treated THP-1 Macrophages Exhibits Alterations in Atherosclerosis-Related Regulator Molecules and LXR/RXR Pathway. International Journal of Molecular Sciences. 2023; 24(1):278. https://doi.org/10.3390/ijms24010278
Chicago/Turabian StyleHuwait, Etimad, Rehab Almassabi, Sanaa Almowallad, Salma Saddeek, Sajjad Karim, Gauthaman Kalamegam, and Zeenat Mirza. 2023. "Microarray Expression Profile of Myricetin-Treated THP-1 Macrophages Exhibits Alterations in Atherosclerosis-Related Regulator Molecules and LXR/RXR Pathway" International Journal of Molecular Sciences 24, no. 1: 278. https://doi.org/10.3390/ijms24010278
APA StyleHuwait, E., Almassabi, R., Almowallad, S., Saddeek, S., Karim, S., Kalamegam, G., & Mirza, Z. (2023). Microarray Expression Profile of Myricetin-Treated THP-1 Macrophages Exhibits Alterations in Atherosclerosis-Related Regulator Molecules and LXR/RXR Pathway. International Journal of Molecular Sciences, 24(1), 278. https://doi.org/10.3390/ijms24010278