

PLA2G7/PAF-AH as Potential Negative Regulator of the Wnt Signaling Pathway Mediates Protective Effects in BRCA1 Mutant Breast Cancer

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Nuclear PAF-AH Is Highly Expressed in BRCA1 Mutant BC Tissue and Correlates Positively with Membranous β-Catenin
2.2. Only BRCA1 Negative Cell Line HCC1937 Shows Relevant Expression of PLA2G7/PAF-AH
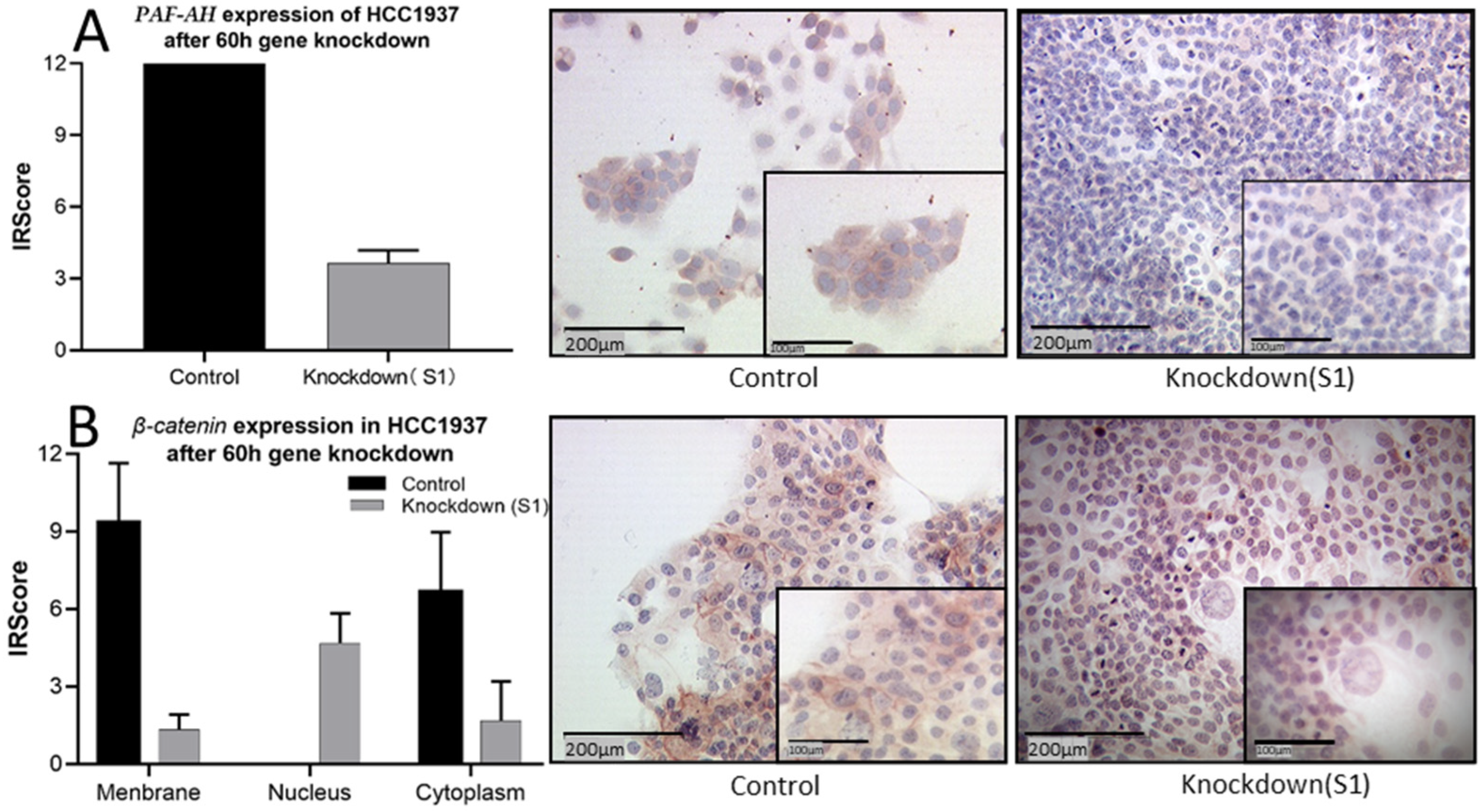
2.3. PLA2G7 Downregulation Enhanced Viability, Proliferation, and Motility of HCC1937 Cells
2.4. The Intracellular Distribution of β-Catenin Changed from High Expression in the Membrane to High Expression in the Nucleus by PLA2G7 Knockdown
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Patient Specimens
4.3. Immunohistochemistry and Immunocytochemistry
4.4. Staining Evaluation
4.5. Statistical Analysis
4.6. Cell Lines
4.7. Real-Time PCR
4.8. siRNA Gene Knockdown
4.9. Western Blot
4.10. Cell Viability Assays
4.11. Proliferation Assay
4.12. Wound Healing Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed]
- Eccles, S.A.; Aboagye, E.O.; Ali, S.; Anderson, A.S.; Armes, J.; Berditchevski, F.; Blaydes, J.P.; Brennan, K.; Brown, N.J.; Bryant, H.E.; et al. Critical research gaps and translational priorities for the successful prevention and treatment of breast cancer. Breast Cancer Res. BCR 2013, 15, R92. [Google Scholar] [CrossRef] [PubMed]
- Armes, J.E.; Egan, A.J.; Southey, M.C.; Dite, G.S.; McCredie, M.R.; Giles, G.G.; Hopper, J.L.; Venter, D.J. The histologic phenotypes of breast carcinoma occurring before age 40 years in women with and without BRCA1 or BRCA2 germline mutations: A population-based study. Cancer 1998, 83, 2335–2345. [Google Scholar] [CrossRef]
- Diab, S.G.; Clark, G.M.; Osborne, C.K.; Libby, A.; Allred, D.C.; Elledge, R.M. Tumor characteristics and clinical outcome of tubular and mucinous breast carcinomas. J. Clin. Oncol. 1999, 17, 1442–1448. [Google Scholar] [CrossRef]
- Livi, L.; Paiar, F.; Meldolesi, E.; Talamonti, C.; Simontacchi, G.; Detti, B.; Salerno, S.; Bianchi, S.; Cardona, G.; Biti, G.P. Tubular carcinoma of the breast: Outcome and loco-regional recurrence in 307 patients. Eur. J. Surg. Oncol. 2005, 31, 9–12. [Google Scholar] [CrossRef]
- Southey, M.C.; Ramus, S.J.; Dowty, J.G.; Smith, L.D.; Tesoriero, A.A.; Wong, E.E.; Dite, G.S.; Jenkins, M.A.; Byrnes, G.B.; Winship, I.; et al. Morphological predictors of BRCA1 germline mutations in young women with breast cancer. Br. J. Cancer 2011, 104, 903–909. [Google Scholar] [CrossRef]
- Bracci, M.; Ciarapica, V.; Zabaleta, M.E.; Tartaglione, M.F.; Pirozzi, S.; Giuliani, L.; Piva, F.; Valentino, M.; Ledda, C.; Rapisarda, V.; et al. BRCA1 and BRCA2 Gene Expression: Diurnal Variability and Influence of Shift Work. Cancers 2019, 11, 1146. [Google Scholar] [CrossRef]
- Laufer-Amorim, R.; Fonseca-Alves, C.E.; Villacis, R.A.R.; Linde, S.A.D.; Carvalho, M.; Larsen, S.J.; Marchi, F.A.; Rogatto, S.R. Comprehensive Genomic Profiling of Androgen-Receptor-Negative Canine Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 1555. [Google Scholar] [CrossRef]
- Murthy, P.; Muggia, F. Women’s cancers: How the discovery of BRCA genes is driving current concepts of cancer biology and therapeutics. Ecancermedicalscience 2019, 13, 904. [Google Scholar] [CrossRef]
- Li, S.; Wei, Y. Association of HMGB1, BRCA1 and P62 expression in ovarian cancer and chemotherapy sensitivity. Oncol. Lett. 2018, 15, 9572–9576. [Google Scholar] [CrossRef]
- Miron, A.; Schildkraut, J.M.; Rimer, B.K.; Winer, E.P.; Sugg Skinner, C.; Futreal, P.A.; Culler, D.; Calingaert, B.; Clark, S.; Kelly Marcom, P.; et al. Testing for hereditary breast and ovarian cancer in the southeastern United States. Ann. Surg. 2000, 231, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Howell, M.E.A.; Sparks-Wallace, A.; Hawkins, C.; Nicksic, C.A.; Kohne, C.; Hall, K.H.; Moorman, J.P.; Yao, Z.Q.; Ning, S. p62-mediated Selective autophagy endows virus-transformed cells with insusceptibility to DNA damage under oxidative stress. PLoS Pathog. 2019, 15, e1007541. [Google Scholar] [CrossRef] [PubMed]
- Yaneva, M.; Li, H.; Marple, T.; Hasty, P. Non-homologous end joining, but not homologous recombination, enables survival for cells exposed to a histone deacetylase inhibitor. Nucleic Acids Res. 2005, 33, 5320–5330. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Hu, Z.; Liu, Y.; Wang, X.; Hopkins, K.M.; Lieberman, H.B.; Hang, H. Mouse Rad1 deletion enhances susceptibility for skin tumor development. Mol. Cancer 2010, 9, 67. [Google Scholar] [CrossRef]
- Lavin, M.F. ATM and the Mre11 complex combine to recognize and signal DNA double-strand breaks. Oncogene 2007, 26, 7749–7758. [Google Scholar] [CrossRef] [PubMed]
- Rupnik, A.; Lowndes, N.F.; Grenon, M. MRN and the race to the break. Chromosoma 2010, 119, 115–135. [Google Scholar] [CrossRef]
- Bochar, D.A.; Wang, L.; Beniya, H.; Kinev, A.; Xue, Y.; Lane, W.S.; Wang, W.; Kashanchi, F.; Shiekhattar, R. BRCA1 is associated with a human SWI/SNF-related complex: Linking chromatin remodeling to breast cancer. Cell 2000, 102, 257–265. [Google Scholar] [CrossRef]
- Cantor, S.B.; Bell, D.W.; Ganesan, S.; Kass, E.M.; Drapkin, R.; Grossman, S.; Wahrer, D.C.; Sgroi, D.C.; Lane, W.S.; Haber, D.A.; et al. BACH1, a novel helicase-like protein, interacts directly with BRCA1 and contributes to its DNA repair function. Cell 2001, 105, 149–160. [Google Scholar] [CrossRef]
- Kuchenbaecker, K.B.; Hopper, J.L.; Barnes, D.R.; Phillips, K.A.; Mooij, T.M.; Roos-Blom, M.J.; Jervis, S.; van Leeuwen, F.E.; Milne, R.L.; Andrieu, N.; et al. Risks of Breast, Ovarian, and Contralateral Breast Cancer for BRCA1 and BRCA2 Mutation Carriers. JAMA 2017, 317, 2402–2416. [Google Scholar] [CrossRef]
- Castro Faria Neto, H.C.; Stafforini, D.M.; Prescott, S.M.; Zimmerman, G.A. Regulating inflammation through the anti-inflammatory enzyme platelet-activating factor-acetylhydrolase. Mem. Inst. Oswaldo Cruz 2005, 100 (Suppl. S1), 83–91. [Google Scholar] [CrossRef]
- Karasawa, K. Clinical aspects of plasma platelet-activating factor-acetylhydrolase. Biochim. Biophys. Acta 2006, 1761, 1359–1372. [Google Scholar] [CrossRef] [PubMed]
- Stafforini, D.M. Biology of platelet-activating factor acetylhydrolase (PAF-AH, lipoprotein associated phospholipase A2). Cardiovasc. Drugs Ther. 2009, 23, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.M.; Zimmerman, G.A.; Stafforini, D.M.; McIntyre, T.M. Platelet-activating factor and related lipid mediators. Annu. Rev. Biochem. 2000, 69, 419–445. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.A.; Barcelos, L.S.; Teixeira, M.M.; Bakhle, Y.S.; Andrade, S.P. Tumor growth, angiogenesis and inflammation in mice lacking receptors for platelet activating factor (PAF). Life Sci. 2007, 81, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.C.; Russo, R.C.; Guabiraba, R.; Fagundes, C.T.; Polidoro, R.B.; Tavares, L.P.; Salgado, A.P.; Cassali, G.D.; Sousa, L.P.; Machado, A.V.; et al. Platelet-activating factor receptor plays a role in lung injury and death caused by Influenza A in mice. PLoS Pathog. 2010, 6, e1001171. [Google Scholar] [CrossRef]
- Kume, K.; Shimizu, T. Platelet-activating factor (PAF) induces growth stimulation, inhibition, and suppression of oncogenic transformation in NRK cells overexpressing the PAF receptor. J. Biol. Chem. 1997, 272, 22898–22904. [Google Scholar] [CrossRef]
- Melnikova, V.O.; Balasubramanian, K.; Villares, G.J.; Dobroff, A.S.; Zigler, M.; Wang, H.; Petersson, F.; Price, J.E.; Schroit, A.; Prieto, V.G.; et al. Crosstalk between protease-activated receptor 1 and platelet-activating factor receptor regulates melanoma cell adhesion molecule (MCAM/MUC18) expression and melanoma metastasis. J. Biol. Chem. 2009, 284, 28845–28855. [Google Scholar] [CrossRef]
- Lehtinen, L.; Vainio, P.; Wikman, H.; Huhtala, H.; Mueller, V.; Kallioniemi, A.; Pantel, K.; Kronqvist, P.; Kallioniemi, O.; Carpèn, O.; et al. PLA2G7 associates with hormone receptor negativity in clinical breast cancer samples and regulates epithelial-mesenchymal transition in cultured breast cancer cells. J. Pathol. Clin. Res. 2017, 3, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Vainio, P.; Lehtinen, L.; Mirtti, T.; Hilvo, M.; Seppänen-Laakso, T.; Virtanen, J.; Sankila, A.; Nordling, S.; Lundin, J.; Rannikko, A.; et al. Phospholipase PLA2G7, associated with aggressive prostate cancer, promotes prostate cancer cell migration and invasion and is inhibited by statins. Oncotarget 2011, 2, 1176–1190. [Google Scholar] [CrossRef]
- Biancone, L.; Cantaluppi, V.; Del Sorbo, L.; Russo, S.; Tjoelker, L.W.; Camussi, G. Platelet-activating factor inactivation by local expression of platelet-activating factor acetyl-hydrolase modifies tumor vascularization and growth. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2003, 9, 4214–4220. [Google Scholar]
- Liao, Y.; Badmann, S.; Kaltofen, T.; Mayr, D.; Schmoeckel, E.; Deuster, E.; Mannewitz, M.; Landgrebe, S.; Kolben, T.; Hester, A.; et al. Platelet-Activating Factor Acetylhydrolase Expression in BRCA1 Mutant Ovarian Cancer as a Protective Factor and Potential Negative Regulator of the Wnt Signaling Pathway. Biomedicines 2021, 9, 706. [Google Scholar] [CrossRef] [PubMed]
- Khramtsov, A.I.; Khramtsova, G.F.; Tretiakova, M.; Huo, D.; Olopade, O.I.; Goss, K.H. Wnt/beta-catenin pathway activation is enriched in basal-like breast cancers and predicts poor outcome. Am. J. Pathol. 2010, 176, 2911–2920. [Google Scholar] [CrossRef] [PubMed]
- López-Knowles, E.; Zardawi, S.J.; McNeil, C.M.; Millar, E.K.; Crea, P.; Musgrove, E.A.; Sutherland, R.L.; O’Toole, S.A. Cytoplasmic localization of beta-catenin is a marker of poor outcome in breast cancer patients. Cancer Epidemiol. Biomark. Prev. 2010, 19, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Rask, K.; Nilsson, A.; Brännström, M.; Carlsson, P.; Hellberg, P.; Janson, P.O.; Hedin, L.; Sundfeldt, K. Wnt-signalling pathway in ovarian epithelial tumours: Increased expression of beta-catenin and GSK3beta. Br. J. Cancer 2003, 89, 1298–1304. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Duchartre, Y.; Kim, Y.M.; Kahn, M. The Wnt signaling pathway in cancer. Crit Rev. Oncol. Hematol. 2016, 99, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, C.; Sharma, M.; Henderson, B.R. Targeting the β-catenin nuclear transport pathway in cancer. Semin. Cancer Biol. 2014, 27, 20–29. [Google Scholar] [CrossRef]
- Bànkfalvi, A.; Terpe, H.J.; Breukelmann, D.; Bier, B.; Rempe, D.; Pschadka, G.; Krech, R.; Lellè, R.J.; Boecker, W. Immunophenotypic and prognostic analysis of E-cadherin and beta-catenin expression during breast carcinogenesis and tumour progression: A comparative study with CD44. Histopathology 1999, 34, 25–34. [Google Scholar] [CrossRef]
- Lin, S.Y.; Xia, W.; Wang, J.C.; Kwong, K.Y.; Spohn, B.; Wen, Y.; Pestell, R.G.; Hung, M.C. Beta-catenin, a novel prognostic marker for breast cancer: Its roles in cyclin D1 expression and cancer progression. Proc. Natl. Acad. Sci. USA 2000, 97, 4262–4266. [Google Scholar] [CrossRef]
- Wong, S.C.; Lo, S.F.; Lee, K.C.; Yam, J.W.; Chan, J.K.; Wendy Hsiao, W.L. Expression of frizzled-related protein and Wnt-signalling molecules in invasive human breast tumours. J. Pathol. 2002, 196, 145–153. [Google Scholar] [CrossRef]
- Jung, H.Y.; Jun, S.; Lee, M.; Kim, H.C.; Wang, X.; Ji, H.; McCrea, P.D.; Park, J.I. PAF and EZH2 induce Wnt/β-catenin signaling hyperactivation. Mol. Cell 2013, 52, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jung, Y.S.; Jun, S.; Lee, S.; Wang, W.; Schneider, A.; Sun Oh, Y.; Lin, S.H.; Park, B.J.; Chen, J.; et al. PAF-Wnt signaling-induced cell plasticity is required for maintenance of breast cancer cell stemness. Nat. Commun. 2016, 7, 10633. [Google Scholar] [CrossRef] [PubMed]
- Livnat, I.; Finkelshtein, D.; Ghosh, I.; Arai, H.; Reiner, O. PAF-AH Catalytic Subunits Modulate the Wnt Pathway in Developing GABAergic Neurons. Front. Cell. Neurosci. 2010, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Arai, H. Platelet-activating factor acetylhydrolase. Prostaglandins Other Lipid Mediat. 2002, 68–69, 83–94. [Google Scholar] [CrossRef]
- Karasawa, K.; Harada, A.; Satoh, N.; Inoue, K.; Setaka, M. Plasma platelet activating factor-acetylhydrolase (PAF-AH). Prog. Lipid Res. 2003, 42, 93–114. [Google Scholar] [CrossRef] [PubMed]
- Hong, R.; Zhang, W.; Xia, X.; Zhang, K.; Wang, Y.; Wu, M.; Fan, J.; Li, J.; Xia, W.; Xu, F.; et al. Preventing BRCA1/ZBRK1 repressor complex binding to the GOT2 promoter results in accelerated aspartate biosynthesis and promotion of cell proliferation. Mol. Oncol. 2019, 13, 959–977. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Baniasadi, P.S.; Harris, I.S.; Silvester, J.; Inoue, S.; Snow, B.; Joshi, P.A.; Wakeham, A.; Molyneux, S.D.; Martin, B.; et al. BRCA1 interacts with Nrf2 to regulate antioxidant signaling and cell survival. J. Exp. Med. 2013, 210, 1529–1544. [Google Scholar] [CrossRef]
- Dodson, M.; de la Vega, M.R.; Cholanians, A.B.; Schmidlin, C.J.; Chapman, E.; Zhang, D.D. Modulating NRF2 in Disease: Timing Is Everything. Annu. Rev. Pharm. Toxicol. 2019, 59, 555–575. [Google Scholar] [CrossRef]
- Paek, J.; Lo, J.Y.; Narasimhan, S.D.; Nguyen, T.N.; Glover-Cutter, K.; Robida-Stubbs, S.; Suzuki, T.; Yamamoto, M.; Blackwell, T.K.; Curran, S.P. Mitochondrial SKN-1/Nrf mediates a conserved starvation response. Cell Metab. 2012, 16, 526–537. [Google Scholar] [CrossRef]
- Wu, Z.Q.; Li, X.Y.; Hu, C.Y.; Ford, M.; Kleer, C.G.; Weiss, S.J. Canonical Wnt signaling regulates Slug activity and links epithelial-mesenchymal transition with epigenetic Breast Cancer 1, Early Onset (BRCA1) repression. Proc. Natl. Acad. Sci. USA 2012, 109, 16654–16659. [Google Scholar] [CrossRef]
- Li, H.; Sekine, M.; Tung, N.; Avraham, H.K. Wild-type BRCA1, but not mutated BRCA1, regulates the expression of the nuclear form of beta-catenin. Mol. Cancer Res. 2010, 8, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, D.; Jiang, W.; Edwards, D.; Qiu, W.; Barroilhet, L.M.; Rho, J.H.; Jin, L.; Seethappan, V.; Vitonis, A.; et al. Activated networking of platelet activating factor receptor and FAK/STAT1 induces malignant potential in BRCA1-mutant at-risk ovarian epithelium. Reprod. Biol. Endocrinol. 2010, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Scholz, C.; Heublein, S.; Lenhard, M.; Friese, K.; Mayr, D.; Jeschke, U. Glycodelin A is a prognostic marker to predict poor outcome in advanced stage ovarian cancer patients. BMC Res. Notes 2012, 5, 551. [Google Scholar] [CrossRef] [PubMed]
- Remmele, W.; Stegner, H.E. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Pathologe 1987, 8, 138–140. [Google Scholar] [PubMed]
- Whitney, J. Testing for differences with the nonparametric Mann-Whitney U test. J. Wound Ostomy Cont. Nurs. 1997, 24, 12. [Google Scholar] [CrossRef]
- Spearman’s rank correlation coefficient. BMJ 2018, 362, k4131. [CrossRef]
- Thomsen, R.; Sølvsten, C.A.; Linnet, T.E.; Blechingberg, J.; Nielsen, A.L. Analysis of qPCR data by converting exponentially related Ct values into linearly related X0 values. J. Bioinform. Comput. Biol. 2010, 8, 885–900. [Google Scholar] [CrossRef]
- Tremmel, E.; Hofmann, S.; Kuhn, C.; Heidegger, H.; Heublein, S.; Hermelink, K.; Wuerstlein, R.; Harbeck, N.; Mayr, D.; Mahner, S.; et al. Thyronamine regulation of TAAR1 expression in breast cancer cells and investigation of its influence on viability and migration. Breast Cancer 2019, 11, 87–97. [Google Scholar] [CrossRef]
- Campion, E.M.; Loughran, S.T.; Walls, D. Protein Quantitation and Analysis of Purity. Methods Mol. Biol. 2017, 1485, 225–255. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAF-AH Nucleus | PAF-AH Cytoplasm | |
|---|---|---|
| β-catenin nucleus | ||
| Cc | −0.062 | 0.018 |
| P | >0.05 | >0.05 |
| N | 44 | 44 |
| β-catenin membrane | ||
| Cc | 0.766 | 0.057 |
| P | <0.001 * | >0.05 |
| N | 44 | 44 |
| Clinicopathological Characteristics of Patients | |||
|---|---|---|---|
| n (121) | Percentage [%] | ||
| Histology | Invasive ductal | 80 | 66.1 |
| Invasive lobular | 13 | 10.7 | |
| Invasive medullary | 12 | 9.9 | |
| Invasively mucinous | 1 | 0.8 | |
| Unknown | 15 | 11.8 | |
| Grading | 1 | 4 | 3.3 |
| 2 | 45 | 37.1 | |
| 3 | 60 | 49.6 | |
| Unknown | 12 | 9.9 | |
| Age | ≥60 | 29 | 24.0 |
| <60 | 92 | 76.0 | |
| Primary tumor expansion | Tis | 5 | 4.1 |
| T1a | 3 | 2.5 | |
| T1b | 13 | 10.7 | |
| T1c | 22 | 21.5 | |
| T2 | 43 | 36.2 | |
| T3 | 12 | 9.9 | |
| T4a | 2 | 1.6 | |
| T4b | 2 | 1.6 | |
| T4c | 1 | 0.8 | |
| T4d | 6 | 5.0 | |
| Tx | 12 | 9.9 | |
| Nodal status | N0 | 38 | 31.4 |
| N1 | 34 | 28.1 | |
| N2 | 9 | 7.4 | |
| N3 | 7 | 5.8 | |
| Nx | 33 | 27.3 | |
| Distant Metastasis | M0 | 46 | 38.0 |
| M1 | 28 | 23.1 | |
| Mx | 47 | 53.5 | |
| BRCA1 mutation status | No BRCA1 mutation | 28 | 23.1 |
| BRCA1 mutation | 22 | 18.2 | |
| Unknown | 71 | 58.7 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Y.; Badmann, S.; Kraus, F.; Topalov, N.E.; Mayr, D.; Kolben, T.; Hester, A.; Beyer, S.; Mahner, S.; Jeschke, U.; et al. PLA2G7/PAF-AH as Potential Negative Regulator of the Wnt Signaling Pathway Mediates Protective Effects in BRCA1 Mutant Breast Cancer. Int. J. Mol. Sci. 2023, 24, 882. https://doi.org/10.3390/ijms24010882
Liao Y, Badmann S, Kraus F, Topalov NE, Mayr D, Kolben T, Hester A, Beyer S, Mahner S, Jeschke U, et al. PLA2G7/PAF-AH as Potential Negative Regulator of the Wnt Signaling Pathway Mediates Protective Effects in BRCA1 Mutant Breast Cancer. International Journal of Molecular Sciences. 2023; 24(1):882. https://doi.org/10.3390/ijms24010882
Chicago/Turabian StyleLiao, Yue, Susann Badmann, Fabian Kraus, Nicole Elisabeth Topalov, Doris Mayr, Thomas Kolben, Anna Hester, Susanne Beyer, Sven Mahner, Udo Jeschke, and et al. 2023. "PLA2G7/PAF-AH as Potential Negative Regulator of the Wnt Signaling Pathway Mediates Protective Effects in BRCA1 Mutant Breast Cancer" International Journal of Molecular Sciences 24, no. 1: 882. https://doi.org/10.3390/ijms24010882
APA StyleLiao, Y., Badmann, S., Kraus, F., Topalov, N. E., Mayr, D., Kolben, T., Hester, A., Beyer, S., Mahner, S., Jeschke, U., Trillsch, F., Czogalla, B., & Burges, A. (2023). PLA2G7/PAF-AH as Potential Negative Regulator of the Wnt Signaling Pathway Mediates Protective Effects in BRCA1 Mutant Breast Cancer. International Journal of Molecular Sciences, 24(1), 882. https://doi.org/10.3390/ijms24010882