
Dynamic Evolution of Humoral and T-Cell Specific Immune Response to COVID-19 mRNA Vaccine in Patients with Multiple Sclerosis Followed until the Booster Dose

 , , , ,
, , , ,  , ,
, ,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Study Population
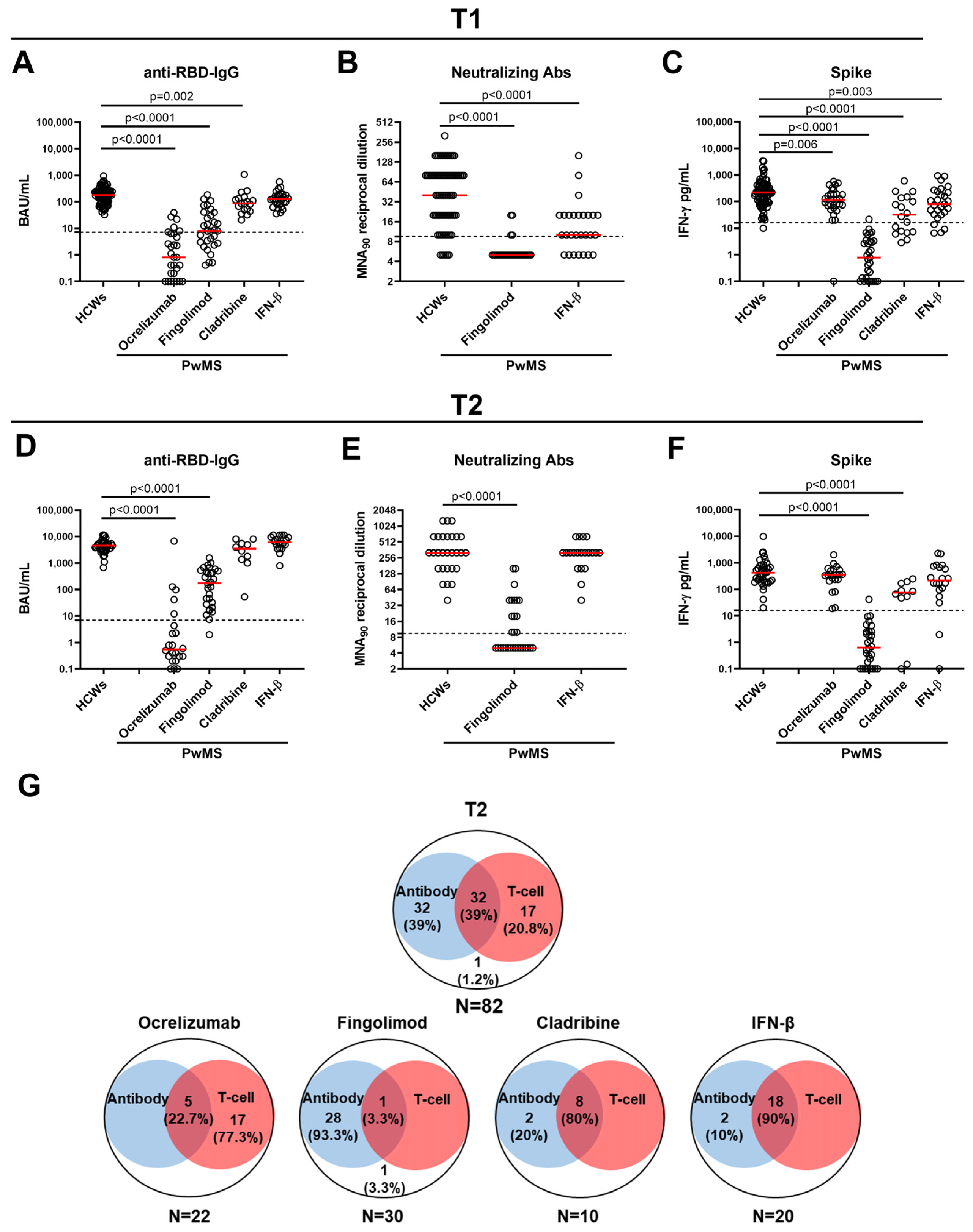
2.2. Antibody and Spike-Specific T-Cell Response in PwMS and HCWs at T1
2.3. Antibody and Spike-Specific T-Cell Response in PwMS and HCWs at T2
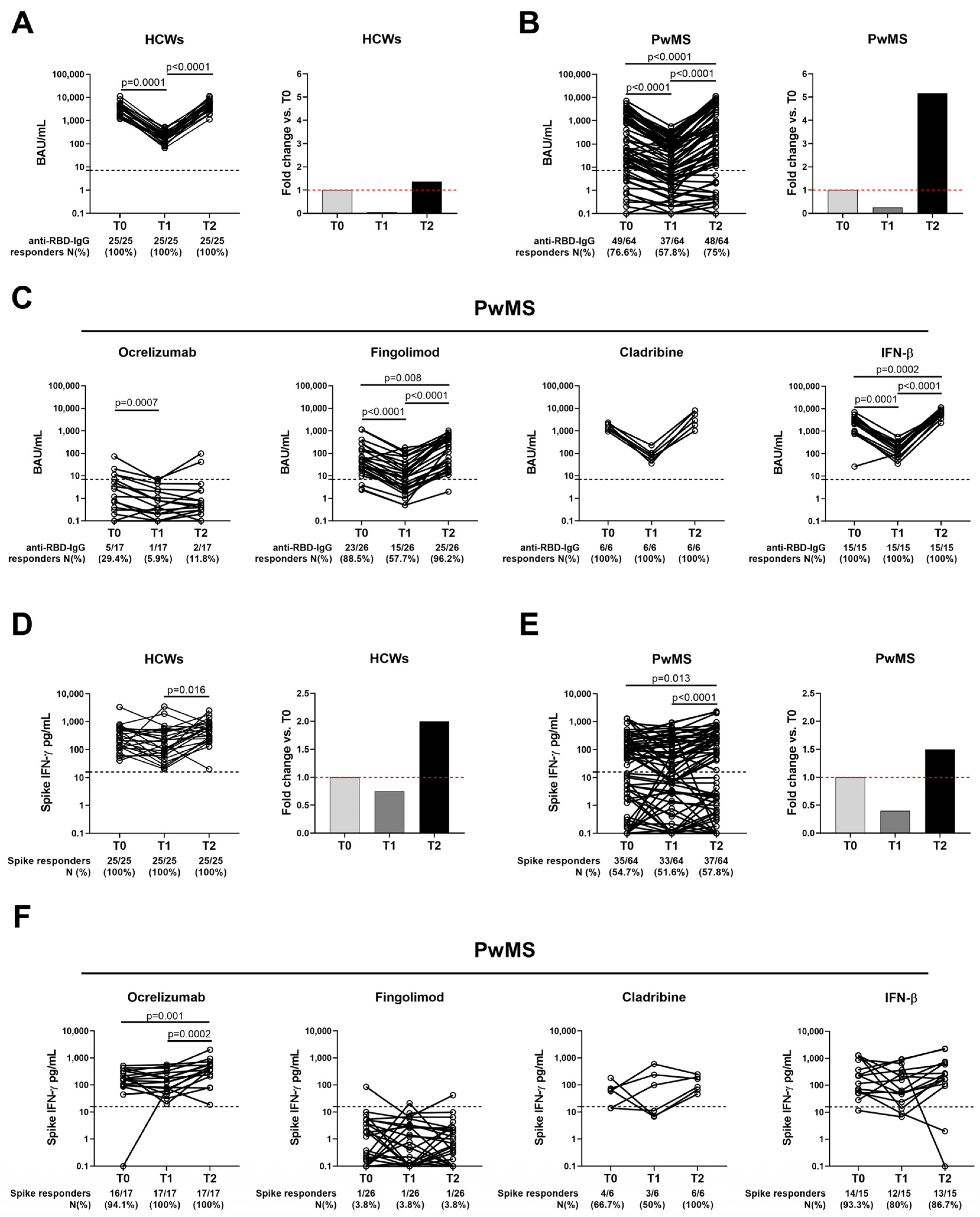
2.4. Temporal Evolution of the Antibody Response to COVID-19 Vaccination in Both HCWs and PwMS
2.5. SARS-CoV-2-Spike-Specific T-Cell Response Persists over Time in Both Cohorts
3. Discussion
3.1. Limitations of the Study
3.2. Conclusions
4. Materials and Methods
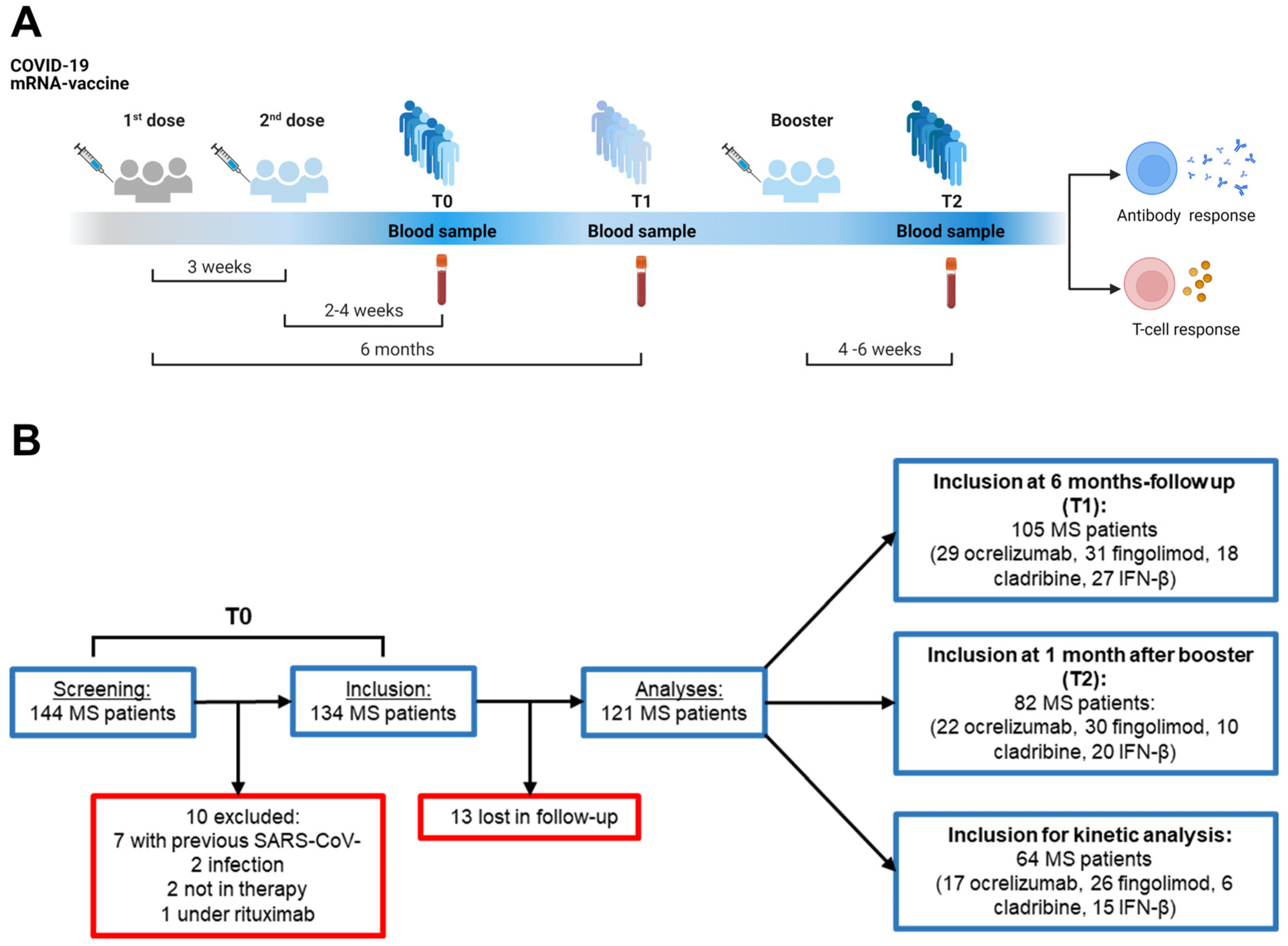
4.1. Study Population
4.2. Study Procedures
4.3. Anti-SARS-CoV-2 Antibody Testing
4.4. IFN-γ Release Assay (IGRA)
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobson, R.; Giovannoni, G. Multiple Sclerosis—A Review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [PubMed]
- McGinley, M.P.; Goldschmidt, C.H.; Rae-Grant, A.D. Diagnosis and Treatment of Multiple Sclerosis: A Review. JAMA 2021, 325, 765–779. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, L.; Tortorella, C.; Haggiag, S.; Ruggieri, S.; Galgani, S.; Gasperini, C. Increased Risk of Death from COVID-19 in Multiple Sclerosis: A Pooled Analysis of Observational Studies. J. Neurol. 2022, 269, 1114–1120. [Google Scholar] [CrossRef]
- Sormani, M.P.; Schiavetti, I.; Landi, D.; Carmisciano, L.; De Rossi, N.; Cordioli, C.; Moiola, L.; Radaelli, M.; Immovilli, P.; Capobianco, M.; et al. SARS-CoV-2 Serology after COVID-19 in Multiple Sclerosis: An International Cohort Study. Mult. Scler. 2022, 28, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Coronavirus Disease (COVID-19)—World Health Organization. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 2 May 2023).
- Barouch, D.H. COVID-19 Vaccines—Immunity, Variants, Boosters. N. Engl. J. Med. 2022, 387, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Suthar, A.B.; Wang, J.; Seffren, V.; Wiegand, R.E.; Griffing, S.; Zell, E. Public Health Impact of COVID-19 Vaccines in the US: Observational Study. BMJ 2022, 377, e069317. [Google Scholar] [CrossRef]
- Sunagar, R.; Singh, A.; Kumar, S. SARS-CoV-2: Immunity, Challenges with Current Vaccines, and a Novel Perspective on Mucosal Vaccines. Vaccines 2023, 11, 849. [Google Scholar] [CrossRef]
- Il Sole 24 ORE. Il Vaccino Anti COVID in Italia in Tempo Reale. Available online: https://lab24.ilsole24ore.com/numeri-vaccini-italia-mondo/?refresh_ce=1 (accessed on 2 May 2023).
- Dhawan, M.; Rabaan, A.A.; Fawarah, M.M.A.; Almuthree, S.A.; Alsubki, R.A.; Alfaraj, A.H.; Mashraqi, M.M.; Alshamrani, S.A.; Abduljabbar, W.A.; Alwashmi, A.S.S.; et al. Updated Insights into the T Cell-Mediated Immune Response against SARS-CoV-2: A Step towards Efficient and Reliable Vaccines. Vaccines 2023, 11, 101. [Google Scholar] [CrossRef]
- Gilbert, P.B.; Montefiori, D.C.; McDermott, A.B.; Fong, Y.; Benkeser, D.; Deng, W.; Zhou, H.; Houchens, C.R.; Martins, K.; Jayashankar, L.; et al. Immune Correlates Analysis of the MRNA-1273 COVID-19 Vaccine Efficacy Clinical Trial. Science 2022, 375, 43–50. [Google Scholar] [CrossRef]
- Goletti, D.; Petrone, L.; Manissero, D.; Bertoletti, A.; Rao, S.; Ndunda, N.; Sette, A.; Nikolayevskyy, V. The Potential Clinical Utility of Measuring Severe Acute Respiratory Syndrome Coronavirus 2-Specific T-Cell Responses. Clin. Microbiol. Infect. 2021, 27, 1784–1789. [Google Scholar] [CrossRef]
- Moss, P. The T Cell Immune Response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Shitrit, P.; Zuckerman, N.S.; Mor, O.; Gottesman, B.-S.; Chowers, M. Nosocomial Outbreak Caused by the SARS-CoV-2 Delta Variant in a Highly Vaccinated Population, Israel, July 2021. Eurosurveillance 2021, 26, 2100822. [Google Scholar] [CrossRef]
- Chau, N.V.V.; Ngoc, N.M.; Nguyet, L.A.; Quang, V.M.; Ny, N.T.H.; Khoa, D.B.; Phong, N.T.; Toan, L.M.; Hong, N.T.T.; Tuyen, N.T.K.; et al. An Observational Study of Breakthrough SARS-CoV-2 Delta Variant Infections among Vaccinated Healthcare Workers in Vietnam. EClinicalMedicine 2021, 41, 101143. [Google Scholar] [CrossRef] [PubMed]
- Sansone, E.; Collatuzzo, G.; Renzetti, S.; Ditano, G.; Bonfanti, C.; Sala, E.; Terlenghi, L.; Matteelli, A.; Abedini, M.; Asafo, S.S.; et al. The Effect of the Immunization Schedule and Antibody Levels (Anti-S) on the Risk of SARS-CoV-2 Infection in a Large Cohort of Healthcare Workers in Northern Italy. Vaccines 2023, 11, 746. [Google Scholar] [CrossRef] [PubMed]
- Shrotri, M.; Navaratnam, A.M.D.; Nguyen, V.; Byrne, T.; Geismar, C.; Fragaszy, E.; Beale, S.; Fong, W.L.E.; Patel, P.; Kovar, J.; et al. Spike-Antibody Waning after Second Dose of BNT162b2 or ChAdOx1. Lancet 2021, 398, 385–387. [Google Scholar] [CrossRef]
- Farroni, C.; Picchianti-Diamanti, A.; Aiello, A.; Nicastri, E.; Laganà, B.; Agrati, C.; Castilletti, C.; Meschi, S.; Colavita, F.; Cuzzi, G.; et al. Kinetics of the B- and T-Cell Immune Responses After 6 Months From SARS-CoV-2 MRNA Vaccination in Patients With Rheumatoid Arthritis. Front. Immunol. 2022, 13, 846753. [Google Scholar] [CrossRef]
- Bajwa, H.M.; Novak, F.; Nilsson, A.C.; Nielsen, C.; Holm, D.K.; Østergaard, K.; Witt, A.H.; Byg, K.-E.; Johansen, I.S.; Mittl, K.; et al. Persistently Reduced Humoral and Sustained Cellular Immune Response from First to Third SARS-CoV-2 MRNA Vaccination in Anti-CD20-Treated Multiple Sclerosis Patients. Mult. Scler. Relat. Disord. 2022, 60, 103729. [Google Scholar] [CrossRef]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning Immune Humoral Response to BNT162b2 COVID-19 Vaccine over 6 Months. N. Engl. J. Med. 2021, 385, e84. [Google Scholar] [CrossRef]
- Gillot, C.; Bayart, J.-L.; Closset, M.; Cabo, J.; Maloteau, V.; Dogné, J.-M.; Douxfils, J.; Favresse, J. Peri-Infection Titers of Neutralizing and Binding Antibodies as a Predictor of COVID-19 Breakthrough Infections in Vaccinated Healthcare Professionals: Importance of the Timing. Clin. Chem. Lab. Med. 2023. [Google Scholar] [CrossRef]
- Santoro, A.; Capri, A.; Petrone, D.; Colavita, F.; Meschi, S.; Matusali, G.; Mizzoni, K.; Notari, S.; Agrati, C.; Goletti, D.; et al. SARS-CoV-2 Breakthrough Infections According to the Immune Response Elicited after MRNA Third Dose Vaccination in COVID-19-Naïve Hospital Personnel. Biomedicines 2023, 11, 1247. [Google Scholar] [CrossRef]
- Menegale, F.; Manica, M.; Zardini, A.; Guzzetta, G.; Marziano, V.; d’Andrea, V.; Trentini, F.; Ajelli, M.; Poletti, P.; Merler, S. Evaluation of Waning of SARS-CoV-2 Vaccine-Induced Immunity: A Systematic Review and Meta-Analysis. JAMA Netw. Open. 2023, 6, e2310650. [Google Scholar] [CrossRef]
- EMA Comirnaty and Spikevax: EMA Recommendations on Extra Doses Boosters. Available online: https://www.ema.europa.eu/en/news/comirnaty-spikevax-ema-recommendations-extra-doses-boosters (accessed on 2 January 2023).
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Freedman, L.; Kalkstein, N.; Mizrahi, B.; Alroy-Preis, S.; Ash, N.; Milo, R.; et al. Protection of BNT162b2 Vaccine Booster against COVID-19 in Israel. N. Engl. J. Med. 2021, 385, 1393–1400. [Google Scholar] [CrossRef] [PubMed]
- Farroni, C.; Aiello, A.; Picchianti-Diamanti, A.; Laganà, B.; Petruccioli, E.; Agrati, C.; Garbuglia, A.R.; Meschi, S.; Lapa, D.; Cuzzi, G.; et al. Booster Dose of SARS-CoV-2 MRNA Vaccines Strengthens the Specific Immune Response of Patients with Rheumatoid Arthritis: A Prospective Multicenter Longitudinal Study. Int. J. Infect. Dis. 2022, 125, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Tortorella, C.; Aiello, A.; Gasperini, C.; Agrati, C.; Castilletti, C.; Ruggieri, S.; Meschi, S.; Matusali, G.; Colavita, F.; Farroni, C.; et al. Humoral- and T-Cell-Specific Immune Responses to SARS-CoV-2 MRNA Vaccination in Patients With MS Using Different Disease-Modifying Therapies. Neurology 2022, 98, e541–e554. [Google Scholar] [CrossRef]
- Achiron, A.; Mandel, M.; Dreyer-Alster, S.; Harari, G.; Magalashvili, D.; Sonis, P.; Dolev, M.; Menascu, S.; Flechter, S.; Falb, R.; et al. Humoral Immune Response to COVID-19 MRNA Vaccine in Patients with Multiple Sclerosis Treated with High-Efficacy Disease-Modifying Therapies. Ther. Adv. Neurol. Disord. 2021, 14, 17562864211012835. [Google Scholar] [CrossRef]
- Sabatino, J.J.; Mittl, K.; Rowles, W.M.; McPolin, K.; Rajan, J.V.; Laurie, M.T.; Zamecnik, C.R.; Dandekar, R.; Alvarenga, B.D.; Loudermilk, R.P.; et al. Multiple Sclerosis Therapies Differentially Affect SARS-CoV-2 Vaccine–Induced Antibody and T Cell Immunity and Function. JCI Insight 2022, 7, 156978. [Google Scholar] [CrossRef]
- Corradini, P.; Agrati, C.; Apolone, G.; Mantovani, A.; Giannarelli, D.; Marasco, V.; Bordoni, V.; Sacchi, A.; Matusali, G.; Salvarani, C.; et al. Humoral and T-Cell Immune Response after Three Doses of MRNA SARS-CoV-2 Vaccines in Fragile Patients: The Italian VAX4FRAIL Study. Clin. Infect. Dis. 2022, 76, e426–e438. [Google Scholar] [CrossRef] [PubMed]
- Sainz de la Maza, S.; Walo-Delgado, P.E.; Rodríguez-Domínguez, M.; Monreal, E.; Rodero-Romero, A.; Chico-García, J.L.; Pariente, R.; Rodríguez-Jorge, F.; Ballester-González, R.; Villarrubia, N.; et al. Short- and Long-Term Humoral and Cellular Immune Responses to SARS-CoV-2 Vaccination in Patients with Multiple Sclerosis Treated with Disease-Modifying Therapies. Vaccines 2023, 11, 786. [Google Scholar] [CrossRef]
- König, M.; Torgauten, H.M.; Tran, T.T.; Holmøy, T.; Vaage, J.T.; Lund-Johansen, F.; Nygaard, G.O. Immunogenicity and Safety of a Third SARS-CoV-2 Vaccine Dose in Patients with Multiple Sclerosis and Weak Immune Response after COVID-19 Vaccination. JAMA Neurol. 2022, 79, 307–309. [Google Scholar] [CrossRef]
- Madelon, N.; Heikkilä, N.; Sabater Royo, I.; Fontannaz, P.; Breville, G.; Lauper, K.; Goldstein, R.; Grifoni, A.; Sette, A.; Siegrist, C.-A.; et al. Omicron-Specific Cytotoxic T-Cell Responses After a Third Dose of MRNA COVID-19 Vaccine Among Patients With Multiple Sclerosis Treated With Ocrelizumab. JAMA Neurol. 2022, 79, 399–404. [Google Scholar] [CrossRef]
- Achiron, A.; Mandel, M.; Dreyer-Alster, S.; Magalashvili, D.; Menascu, S.; Warszawer, Y.; Dolev, M.; Didikin, M.; Harari, G.; Sonis, P.; et al. In-Depth Characterization of Long-Term Humoral and Cellular Immune Responses to COVID-19m-RNA Vaccination in Multiple Sclerosis Patients Treated with Teriflunomide or Alemtuzumab. Mult. Scler. Relat. Disord. 2023, 72, 104616. [Google Scholar] [CrossRef] [PubMed]
- Katz Sand, I.; Gnjatic, S.; Krammer, F.; Tuballes, K.; Carreño, J.M.; Satyanarayan, S.; Filomena, S.; Staker, E.; Tcheou, J.; Miller, A.; et al. Evaluation of Immunological Responses to Third COVID-19 Vaccine among People Treated with Sphingosine Receptor-1 Modulators and Anti-CD20 Therapy. Mult. Scler. Relat. Disord. 2023, 70, 104486. [Google Scholar] [CrossRef]
- Dreyer-Alster, S.; Menascu, S.; Mandel, M.; Shirbint, E.; Magalashvili, D.; Dolev, M.; Flechter, S.; Givon, U.; Guber, D.; Stern, Y.; et al. COVID-19 Vaccination in Patients with Multiple Sclerosis: Safety and Humoral Efficacy of the Third Booster Dose. J. Neurol. Sci. 2022, 434, 120155. [Google Scholar] [CrossRef] [PubMed]
- Maglione, A.; Morra, M.; Meroni, R.; Matta, M.; Clerico, M.; Rolla, S. Humoral Response after the Booster Dose of Anti-SARS-CoV-2 Vaccine in Multiple Sclerosis Patients Treated with High-Efficacy Therapies. Mult. Scler. Relat. Disord. 2022, 61, 103776. [Google Scholar] [CrossRef] [PubMed]
- Tallantyre, E.C.; Scurr, M.J.; Vickaryous, N.; Richards, A.; Anderson, V.; Baker, D.; Chance, R.; Evangelou, N.; George, K.; Giovannoni, G.; et al. Response to COVID-19 Booster Vaccinations in Seronegative People with Multiple Sclerosis. Mult. Scler. Relat. Disord. 2022, 64, 103937. [Google Scholar] [CrossRef] [PubMed]
- Palomares Cabeza, V.; Kummer, L.Y.L.; Wieske, L.; Hagen, R.R.; Duurland, M.; Konijn, V.A.L.; van Dam, K.P.J.; Stalman, E.W.; van de Sandt, C.E.; Boekel, L.; et al. Longitudinal T-Cell Responses After a Third SARS-CoV-2 Vaccination in Patients With Multiple Sclerosis on Ocrelizumab or Fingolimod. Neurol. Neuroimmunol. Neuroinflamm. 2022, 9, e1178. [Google Scholar] [CrossRef]
- Torres, P.; Sancho-Saldaña, A.; Gil Sánchez, A.; Peralta, S.; Solana, M.J.; Bakkioui, S.; González-Mingot, C.; Quibus, L.; Ruiz-Fernández, E.; San Pedro-Murillo, E.; et al. A Prospective Study of Cellular Immune Response to Booster COVID-19 Vaccination in Multiple Sclerosis Patients Treated with a Broad Spectrum of Disease-Modifying Therapies. J. Neurol. 2023, 270, 2380–2391. [Google Scholar] [CrossRef]
- Lambrianides, A.; Deeba, E.; Hadjiagapiou, M.; Pantzaris, M.; Krashias, G.; Christodoulou, C. SARS-CoV-2-Specific Antibody Responses Following BNT162b2 Vaccination in Individuals with Multiple Sclerosis Receiving Different Disease-Modifying Treatments. Front. Neurol. 2023, 14, 1092999. [Google Scholar] [CrossRef]
- Harbo, H.F.; Gold, R.; Tintoré, M. Sex and Gender Issues in Multiple Sclerosis. Ther. Adv. Neurol. Disord. 2013, 6, 237–248. [Google Scholar] [CrossRef]
- Capuano, R.; Altieri, M.; Conte, M.; Bisecco, A.; d’Ambrosio, A.; Donnarumma, G.; Grimaldi, E.; Coppola, N.; Medici, N.; Galdiero, M.; et al. Humoral Response and Safety of the Third Booster Dose of BNT162b2 MRNA COVID-19 Vaccine in Patients with Multiple Sclerosis Treated with Ocrelizumab or Fingolimod. J. Neurol. 2022, 269, 6185–6192. [Google Scholar] [CrossRef]
- Qi, H.; Liu, B.; Wang, X.; Zhang, L. The Humoral Response and Antibodies against SARS-CoV-2 Infection. Nat. Immunol. 2022, 23, 1008–1020. [Google Scholar] [CrossRef] [PubMed]
- Wieske, L.; van Dam, K.P.J.; Steenhuis, M.; Stalman, E.W.; Kummer, L.Y.L.; van Kempen, Z.L.E.; Killestein, J.; Volkers, A.G.; Tas, S.W.; Boekel, L.; et al. Humoral Responses after Second and Third SARS-CoV-2 Vaccination in Patients with Immune-Mediated Inflammatory Disorders on Immunosuppressants: A Cohort Study. Lancet Rheumatol. 2022, 4, e338–e350. [Google Scholar] [CrossRef] [PubMed]
- Apostolidis, S.A.; Kakara, M.; Painter, M.M.; Goel, R.R.; Mathew, D.; Lenzi, K.; Rezk, A.; Patterson, K.R.; Espinoza, D.A.; Kadri, J.C.; et al. Cellular and Humoral Immune Responses Following SARS-CoV-2 MRNA Vaccination in Patients with Multiple Sclerosis on Anti-CD20 Therapy. Nat. Med. 2021, 27, 1990–2001. [Google Scholar] [CrossRef] [PubMed]
- Milo, R.; Staun-Ram, E.; Karussis, D.; Karni, A.; Hellmann, M.A.; Bar-Haim, E.; Miller, A. Israeli Neuroimmunology Study Group on COVID-19 Vaccination in Multiple Sclerosis Humoral and Cellular Immune Responses to SARS-CoV-2 MRNA Vaccination in Patients with Multiple Sclerosis: An Israeli Multi-Center Experience Following 3 Vaccine Doses. Front. Immunol. 2022, 13, 868915. [Google Scholar] [CrossRef] [PubMed]
- Habek, M.; Željko, C.; Savić Mlakar, A.; Bendelja, K.; Rogić, D.; Adamec, I.; Barun, B.; Gabelić, T.; Krbot Skorić, M. Humoral and Cellular Immunity in Convalescent and Vaccinated COVID-19 People with Multiple Sclerosis: Effects of Disease Modifying Therapies. Mult. Scler. Relat. Disord. 2022, 59, 103682. [Google Scholar] [CrossRef]
- Han, S.; Zhang, X.; Wang, G.; Guan, H.; Garcia, G.; Li, P.; Feng, L.; Zheng, B. FTY720 Suppresses Humoral Immunity by Inhibiting Germinal Center Reaction. Blood 2004, 104, 4129–4133. [Google Scholar] [CrossRef]
- Capasso, N.; Palladino, R.; Cerbone, V.; Spiezia, A.L.; Covelli, B.; Fiore, A.; Lanzillo, R.; Carotenuto, A.; Petracca, M.; Stanziola, L.; et al. Ocrelizumab Effect on Humoral and Cellular Immunity in Multiple Sclerosis and Its Clinical Correlates: A 3-Year Observational Study. J. Neurol. 2022, 270, 272–282. [Google Scholar] [CrossRef]
- Schiavetti, I.; Cordioli, C.; Stromillo, M.L.; Teresa Ferrò, M.; Laroni, A.; Cocco, E.; Cola, G.; Pasquali, L.; Rilla, M.T.; Signoriello, E.; et al. Breakthrough SARS-CoV-2 Infections in MS Patients on Disease-Modifying Therapies. Mult. Scler. 2022, 28, 2106–2111. [Google Scholar] [CrossRef]
- Okamoto, A.; Fujigaki, H.; Iriyama, C.; Goto, N.; Yamamoto, H.; Mihara, K.; Inaguma, Y.; Miura, Y.; Furukawa, K.; Yamamoto, Y.; et al. CD19-Positive Lymphocyte Count Is Critical for Acquisition of Anti-SARS-CoV-2 IgG after Vaccination in B-Cell Lymphoma. Blood Adv. 2022, 6, 3230–3233. [Google Scholar] [CrossRef]
- Brill, L.; Rechtman, A.; Shifrin, A.; Rozenberg, A.; Afanasiev, S.; Zveik, O.; Haham, N.; Levin, N.; Vaknin-Dembinsky, A. Longitudinal Humoral Response in MS Patients Treated with Cladribine Tablets after Receiving the Second and Third Doses of SARS-CoV-2 MRNA Vaccine. Mult. Scler. Relat. Disord. 2022, 63, 103863. [Google Scholar] [CrossRef]
- Conway, S.; Saxena, S.; Baecher-Allan, C.; Krishnan, R.; Houtchens, M.; Glanz, B.; Saraceno, T.J.; Polgar-Turcsanyi, M.; Bose, G.; Bakshi, R.; et al. Preserved T Cell but Attenuated Antibody Response in MS Patients on Fingolimod and Ocrelizumab Following 2nd and 3rd SARS-CoV-2 MRNA Vaccine. Mult. Scler. J. Exp. Transl. Clin. 2023, 9, 20552173231165196. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Coelho, C.H.; Zhang, Z.; Dan, J.M.; Yu, E.D.; Methot, N.; Bloom, N.I.; Goodwin, B.; Phillips, E.; Mallal, S.; et al. SARS-CoV-2 Vaccination Induces Immunological T Cell Memory Able to Cross-Recognize Variants from Alpha to Omicron. Cell 2022, 185, 847–859.e11. [Google Scholar] [CrossRef] [PubMed]
- Petrone, L.; Tortorella, C.; Aiello, A.; Farroni, C.; Ruggieri, S.; Castilletti, C.; Meschi, S.; Cuzzi, G.; Vanini, V.; Palmieri, F.; et al. Humoral and Cellular Response to Spike of Delta SARS-CoV-2 Variant in Vaccinated Patients With Multiple Sclerosis. Front. Neurol. 2022, 13, 881988. [Google Scholar] [CrossRef] [PubMed]
- Baba, C.; Ozcelik, S.; Kaya, E.; Samedzada, U.; Ozdogar, A.T.; Cevik, S.; Dogan, Y.; Ozakbas, S. Three Doses of COVID-19 Vaccines in Multiple Sclerosis Patients Treated with Disease-Modifying Therapies. Mult. Scler. Relat. Disord. 2022, 68, 104119. [Google Scholar] [CrossRef] [PubMed]
- Kshirsagar, M.; Nasir, M.; Mukherjee, S.; Becker, N.; Dodhia, R.; Weeks, W.B.; Ferres, J.L.; Richardson, B. The Risk of Hospitalization and Mortality After Breakthrough SARS-CoV-2 Infection by Vaccine Type: Observational Study of Medical Claims Data. JMIR Public Health Surveill. 2022, 8, e38898. [Google Scholar] [CrossRef] [PubMed]
- Sormani, M.P.; Schiavetti, I.; Inglese, M.; Carmisciano, L.; Laroni, A.; Lapucci, C.; Visconti, V.; Serrati, C.; Gandoglia, I.; Tassinari, T.; et al. Breakthrough SARS-CoV-2 Infections after COVID-19 MRNA Vaccination in MS Patients on Disease Modifying Therapies during the Delta and the Omicron Waves in Italy. EBioMedicine 2022, 80, 104042. [Google Scholar] [CrossRef]
- Aiello, A.; Coppola, A.; Ruggieri, S.; Farroni, C.; Altera, A.M.G.; Salmi, A.; Vanini, V.; Cuzzi, G.; Petrone, L.; Meschi, S.; et al. Longitudinal Characterisation of B and T-Cell Immune Responses after the Booster Dose of COVID-19 MRNA-Vaccine in People with Multiple Sclerosis Using Different Disease-Modifying Therapies. J. Neurol. Neurosurg. Psychiatry 2022, 94, 290–299. [Google Scholar] [CrossRef]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. MRNA Vaccines Induce Durable Immune Memory to SARS-CoV-2 and Variants of Concern. Science 2021, 374, abm0829. [Google Scholar] [CrossRef]
- Bruel, T.; Stéfic, K.; Nguyen, Y.; Toniutti, D.; Staropoli, I.; Porrot, F.; Guivel-Benhassine, F.; Bolland, W.-H.; Planas, D.; Hadjadj, J.; et al. Longitudinal Analysis of Serum Neutralization of SARS-CoV-2 Omicron BA.2, BA.4, and BA.5 in Patients Receiving Monoclonal Antibodies. Cell Rep. Med. 2022, 3, 100850. [Google Scholar] [CrossRef]
- NIH. Information on COVID-19 Treatment, Prevention and Research. Available online: https://www.covid19treatmentguidelines.nih.gov/ (accessed on 5 May 2023).
- Levin, M.J.; Ustianowski, A.; De Wit, S.; Launay, O.; Avila, M.; Templeton, A.; Yuan, Y.; Seegobin, S.; Ellery, A.; Levinson, D.J.; et al. Intramuscular AZD7442 (Tixagevimab-Cilgavimab) for Prevention of COVID-19. N. Engl. J. Med. 2022, 386, 2188–2200. [Google Scholar] [CrossRef]
- Vita, S.; Rosati, S.; Ascoli Bartoli, T.; Beccacece, A.; D’Abramo, A.; Mariano, A.; Scorzolini, L.; Goletti, D.; Nicastri, E. Monoclonal Antibodies for Pre- and Postexposure Prophylaxis of COVID-19: Review of the Literature. Pathogens 2022, 11, 882. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of Multiple Sclerosis: 2017 Revisions of the McDonald Criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.; Najafi Fard, S.; Petruccioli, E.; Petrone, L.; Vanini, V.; Farroni, C.; Cuzzi, G.; Navarra, A.; Gualano, G.; Mosti, S.; et al. Spike Is the Most Recognized Antigen in the Whole-Blood Platform in Both Acute and Convalescent COVID-19 Patients. Int. J. Infect. Dis. 2021, 106, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.; Coppola, A.; Vanini, V.; Petrone, L.; Cuzzi, G.; Salmi, A.; Altera, A.M.G.; Tortorella, C.; Gualano, G.; Gasperini, C.; et al. Accuracy of QuantiFERON SARS-CoV-2 RUO Assay and Characterization of the CD4+ and CD8+ T-Cell-SARS-CoV-2 Response: Comparison with a Homemade IFN-γ Release Assay. Int. J. Infect. Dis. 2022, 122, 841–849. [Google Scholar] [CrossRef]
- Matusali, G.; Colavita, F.; Lapa, D.; Meschi, S.; Bordi, L.; Piselli, P.; Gagliardini, R.; Corpolongo, A.; Nicastri, E.; Antinori, A.; et al. SARS-CoV-2 Serum Neutralization Assay: A Traditional Tool for a Brand-New Virus. Viruses 2021, 13, 655. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | PwMS | HCWs | p Value | |
|---|---|---|---|---|
| N (%) | 121 (55) | 99 (45) | ||
| Age median (IQR) | 47 (39–55) | 43 (31–52) | 0.001 * | |
| Female N (%) | 85 (70.2) | 72 (72.7) | 0.685 § | |
| Origin N (%) | West Europe | 118 (97.5) | 97 (98) | 0.531 § |
| East Europe | 2 (1.7) | 1 (1.0) | ||
| Asia | 0 (0) | 1 (1.0) | ||
| South America | 1 (0.8) | 0 (0) | ||
| BMI (kg/mq), median (IQR) | 23.7 (21.2–26.4) | - | ||
| MS duration, median (IQR) | 14 (8–22) | - | ||
| MS course N (%) | Relapsing-remitting | 110 (90) | - | |
| Primary-progressive | 11 (10) | - | ||
| EDSS score, median (IQR) | 2 (1–4) | - | ||
| Multiple sclerosis treatment N (%) | Ocrelizumab | 34 (28.1) | - | |
| Fingolimod | 35 (28.9) | - | ||
| Cladribine | 20 (16.5) | - | ||
| IFN-β | 32 (26.5) | - | ||
| Lymphocytes count N (%) | Available | 105 (86.8) | 0 (0) | |
| Lymphocytes count N (%) Median × 103/µL (IQR) | Ocrelizumab | 34 (32.4) 1.62 (1.28–1.89) | - | <0.0001 ** |
| Fingolimod | 35 (33.3) 0.65 (0.57–0.91) | - | ||
| Cladribine | 19 (18.1) 1.10 (0.87–1.45) | - | ||
| IFN-β | 17 (16.2) 1.65 (1.53–2.06) | - | ||
| Time between withdrawals, median days (IQR) | T0-T1 | 161 (159–163) | 139 (135–141) | |
| T1-T2 | 63 (52–77) | 168 (159–178) | ||
| T0-T2 | 218 (210–235) | 305 (298–316) | ||
| Characteristics | p Value | ||||||
|---|---|---|---|---|---|---|---|
| PwMS | HCWs | Within MS Cohort | PwMS vs. HCWs | ||||
| N (%) | 105 (54.1) | 89 (45.9) | |||||
| Antibody response | Qualitative response | anti-RBD abs responders N (%) | 69 (65.7) | 89 (100) | <0.0001 § | ||
| anti-RBD abs responders within the subgroups N (%) | Ocrelizumab | 6/29 (20.7) | - | <0.0001 § | <0.0001 § | ||
| Fingolimod | 18/31 (58.1) | - | <0.0001 § | ||||
| Cladribine | 18/18 (100) | - | - | ||||
| IFN-β | 27/27 (100) | - | - | ||||
| Quantitative response | anti-RBD titers BAU/mL median (IQR) | 32 (2.4–104) | 177 (95.5–262.6) | <0.0001 * | |||
| Ocrelizumab | 0.80 (0.15–6.25) | - | <0.0001 # | <0.0001 * | |||
| Fingolimod | 7.9 (2.7–39.7) | - | <0.0001 * | ||||
| Cladribine | 85.9 (48–137) | - | 0.002 * | ||||
| IFN-β | 126.4 (80–184) | - | 0.047 * | ||||
| Spike-specific IFN-γ T-cell response | Qualitative response | spike responders N (%) | 63 (60) | 88 (98.9) | <0.0001 § | ||
| spike responders within the subgroups N (%) | Ocrelizumab | 28/29 (96.5) | - | <0.0001 § | 0.399 § | ||
| Fingolimod | 1/31 (3.2) | - | <0.0001 § | ||||
| Cladribine | 11/18 (61.1) | - | <0.0001 § | ||||
| IFN-β | 23/27 (85.2) | - | 0.002 § | ||||
| Quantitative response | spike IFN-γ levels pg/mL median (IQR) | 39.9 (3.4–142.5) | 218 (93–378) | <0.0001 * | |||
| Ocrelizumab | 114 (64–193) | - | <0.0001 # | 0.006 * | |||
| Fingolimod | 0.8 (0.1–3.3) | - | <0.0001 * | ||||
| Cladribine | 32 (7–139) | - | <0.0001 * | ||||
| IFN-β | 79 (33–237) | - | 0.003 * | ||||
| PwMS vs. HCWs | ||||||
|---|---|---|---|---|---|---|
| RBD | T1 | T2 | ||||
| IRR | 95%CI | p | IRR | 95%CI | p | |
| HCWs | 1.00 | 1.00 | ||||
| IFN-β | 1.96 | 0.78–4.91 | 0.149 | 2.92 | 1.32–6.44 | 0.008 |
| Cladribine | 1.19 | 0.48–2.92 | 0.708 | 1.67 | 0.59–4.77 | 0.334 |
| Fingolimod | 0.32 | 0.14–0.75 | 0.009 | 0.14 | 0.07–0.28 | <0.001 |
| Ocrelizumab | 0.05 | 0.02–0.12 | <0.001 | 0.09 | 0.05–0.18 | <0.001 |
| Weeks elapsed from the vaccination | 0.82 | 0.68–0.97 | <0.001 | 0.72 | 0.61–0.86 | <0.001 |
| Within PwMS cohort | ||||||
| RBD | T1 | T2 | ||||
| IRR | 95%CI | p | IRR | 95%CI | p | |
| IFN-β | 1.00 | 1.00 | ||||
| Cladribine | 0.78 | 0.29–2.11 | 0.624 | 0.50 | 0.08–3.20 | 0.465 |
| Fingolimod | 0.23 | 0.10–0.53 | 0.001 | 0.04 | 0.01–0.16 | <0.001 |
| Ocrelizumab | 0.04 | 0.02–0.12 | <0.001 | 0.03 | 0.01–0.22 | <0.001 |
| Treatment duration | 1.02 | 0.95–1.09 | 0.585 | 0.98 | 0.88–1.10 | 0.776 |
| EDSS | 0.96 | 0.84–1.09 | 0.488 | 1.08 | 0.84–1.39 | 0.546 |
| PwMS vs. HCWs | ||||||
| Spike | T1 | T2 | ||||
| IRR | 95%CI | p | IRR | 95%CI | p | |
| HCWs | 1.00 | 1.00 | ||||
| IFN-β | 0.78 | 0.17–3.45 | 0.743 | 0.55 | 0.27–1.12 | 0.098 |
| Cladribine | 0.33 | 0.07–1.54 | 0.163 | 0.12 | 0.05–0.30 | <0.001 |
| Fingolimod | 0.01 | 0.002–0.04 | <0.001 | 0.003 | 0.002–0.01 | <0.001 |
| Ocrelizumab | 0.73 | 0.18–2.96 | 0.662 | 0.48 | 0.24–0.96 | 0.037 |
| Weeks elapsed from the vaccination | 0.91 | 0.67–1.25 | 0.586 | |||
| Age | 0.97 | 0.96–0.99 | 0.009 | |||
| Within PwMS cohort | ||||||
| Spike | T1 | T2 | ||||
| IRR | 95%CI | p | IRR | 95%CI | p | |
| IFN-β | 1.00 | 1.00 | ||||
| Cladribine | 0.68 | 0.26–1.75 | 0.421 | 0.23 | 0.08–0.72 | 0.012 |
| Fingolimod | 0.02 | 0.01–0.04 | <0.001 | 0.01 | 0.001–0.01 | <0.001 |
| Ocrelizumab | 1.13 | 0.49–2.57 | 0.778 | 0.90 | 0.38–2.15 | 0.814 |
| Lymphocytes | 1.01 | 0.74–1.38 | 0.939 | 0.92 | 0.61–1.40 | 0.709 |
| Characteristics | p Value | ||||||
|---|---|---|---|---|---|---|---|
| PwMS | HCWs | Within MS Cohort | PwMS vs. HCWs | ||||
| N (%) | 82 (68.3) | 38 (31.7) | |||||
| Antibody response | Qualitative response | anti-RBD abs responders N (%) | 64 (78) | 38 (100) | 0.0017 § | ||
| anti-RBD abs responders within the subgroups N (%) | Ocrelizumab | 5/22 (22.7) | - | <0.0001 § | <0.0001 § | ||
| Fingolimod | 29/30 (96.7) | - | 0.257 § | ||||
| Cladribine | 10/10 (100) | - | - | ||||
| IFN-β | 20/20 (100) | - | - | ||||
| Quantitative response | anti-RBD titers BAU/mL median (IQR) | 408 (11.6–4235) | 4516 (3098–5477) | <0.0001 * | |||
| Ocrelizumab | 0.55 (0.27–6.27) | - | <0.0001 # | <0.0001 * | |||
| Fingolimod | 174 (26–514) | - | <0.0001 * | ||||
| Cladribine | 3455 (1300–6060) | - | 0.295 * | ||||
| IFN-β | 6060 (3906–9414) | - | 0.023 * | ||||
| Spike-specific IFN-γ T-cell response | Qualitative response | spike responders N (%) | 49 (59.7) | 38 (100) | <0.0001 § | ||
| spike responders within the subgroups N (%) | Ocrelizumab | 22/22 (100) | - | <0.0001 § | - | ||
| Fingolimod | 1/30 (3.3) | - | <0.0001 § | ||||
| Cladribine | 8/10 (80) | - | 0.005 § | ||||
| IFN-β | 18/20 (90) | - | 0.047 § | ||||
| Quantitative response | spike IFN-γ levels pg/mL median (IQR) | 71.9 (1.2–334.5) | 426 (196–792) | <0.0001 * | |||
| Ocrelizumab | 352 (230–544) | - | <0.0001 # | 0.364 * | |||
| Fingolimod | 0.6 (0.1–2.6) | - | <0.0001 * | ||||
| Cladribine | 75 (35–176) | - | <0.0001 * | ||||
| IFN-β | 215 (99–728) | - | 0.084 * | ||||
| T0 | T1 | T2 | p Value | |||
|---|---|---|---|---|---|---|
| HCWs | N (%) | 25 (100%) | 25 (100%) | 25 (100%) | ||
| Qualitative response | Antibody responders N (%) | 25/25 (100%) | 25/25 (100%) | 25/25 (100%) | - | |
| T-cell responders N (%) | 25/25 (100%) | 25/25 (100%) | 25/25 (100%) | - | ||
| Quantitative response | anti-RBD titers BAU/mL median (IQR) | 3377 (1647–4839) | 178 (132–271) | 4608 (3040–6408) | <0.0001 # | |
| spike IFN-γ levels pg/mL median (IQR) | 262 (118–531) | 196 (66–490) | 525 (234–812) | 0.012 # | ||
| PwMS | N (%) | 64 (100%) | 64 (100%) | 64 (100%) | ||
| Qualitative response | Antibody responders N (%) | 49/64 (76.6%) | 37/64 (57.8%) | 48/64 (75%) | <0.0001 * | |
| T-cell responders N (%) | 35/64 (54.7%) | 33/64 (51.6%) | 37/64 (57.8%) | 0.368 * | ||
| Quantitative response | anti-RBD titers BAU/mL median (IQR) | 52.8 (7.8–1165) | 13.2 (1.6–98.6) | 272.5 (6.0–4558) | <0.0001 # | |
| spike IFN-γ levels pg/mL median (IQR) | 52.4 (1.7–178) | 20.8 (1.7–171) | 78.6 (1.0–351) | 0.031 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruggieri, S.; Aiello, A.; Tortorella, C.; Navarra, A.; Vanini, V.; Meschi, S.; Lapa, D.; Haggiag, S.; Prosperini, L.; Cuzzi, G.; et al. Dynamic Evolution of Humoral and T-Cell Specific Immune Response to COVID-19 mRNA Vaccine in Patients with Multiple Sclerosis Followed until the Booster Dose. Int. J. Mol. Sci. 2023, 24, 8525. https://doi.org/10.3390/ijms24108525
Ruggieri S, Aiello A, Tortorella C, Navarra A, Vanini V, Meschi S, Lapa D, Haggiag S, Prosperini L, Cuzzi G, et al. Dynamic Evolution of Humoral and T-Cell Specific Immune Response to COVID-19 mRNA Vaccine in Patients with Multiple Sclerosis Followed until the Booster Dose. International Journal of Molecular Sciences. 2023; 24(10):8525. https://doi.org/10.3390/ijms24108525
Chicago/Turabian StyleRuggieri, Serena, Alessandra Aiello, Carla Tortorella, Assunta Navarra, Valentina Vanini, Silvia Meschi, Daniele Lapa, Shalom Haggiag, Luca Prosperini, Gilda Cuzzi, and et al. 2023. "Dynamic Evolution of Humoral and T-Cell Specific Immune Response to COVID-19 mRNA Vaccine in Patients with Multiple Sclerosis Followed until the Booster Dose" International Journal of Molecular Sciences 24, no. 10: 8525. https://doi.org/10.3390/ijms24108525
APA StyleRuggieri, S., Aiello, A., Tortorella, C., Navarra, A., Vanini, V., Meschi, S., Lapa, D., Haggiag, S., Prosperini, L., Cuzzi, G., Salmi, A., Quartuccio, M. E., Altera, A. M. G., Garbuglia, A. R., Ascoli Bartoli, T., Galgani, S., Notari, S., Agrati, C., Puro, V., ... Goletti, D. (2023). Dynamic Evolution of Humoral and T-Cell Specific Immune Response to COVID-19 mRNA Vaccine in Patients with Multiple Sclerosis Followed until the Booster Dose. International Journal of Molecular Sciences, 24(10), 8525. https://doi.org/10.3390/ijms24108525