

Interaction of Bacteria, Immune Cells, and Surface Topography in Periprosthetic Joint Infections
 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Results
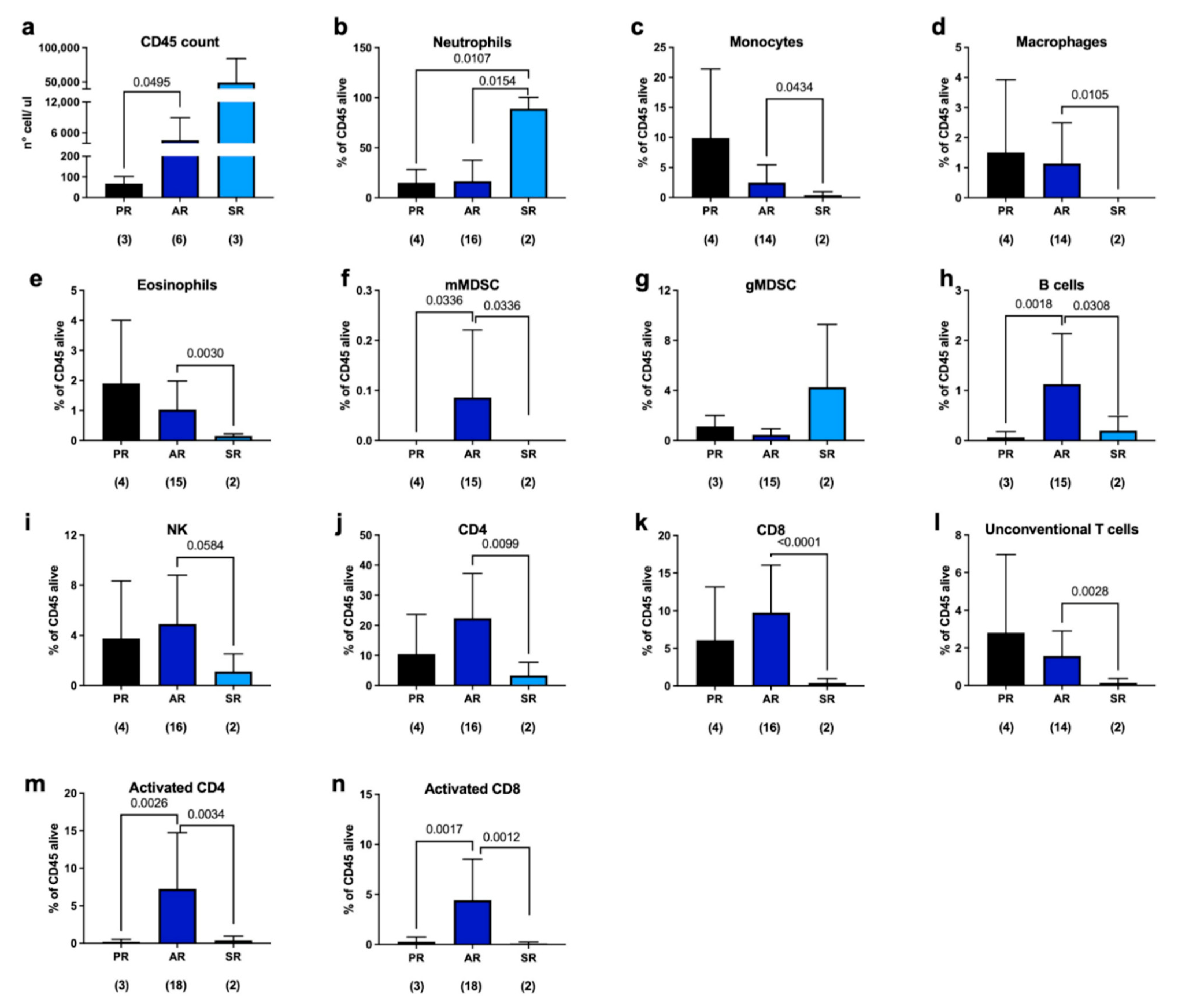
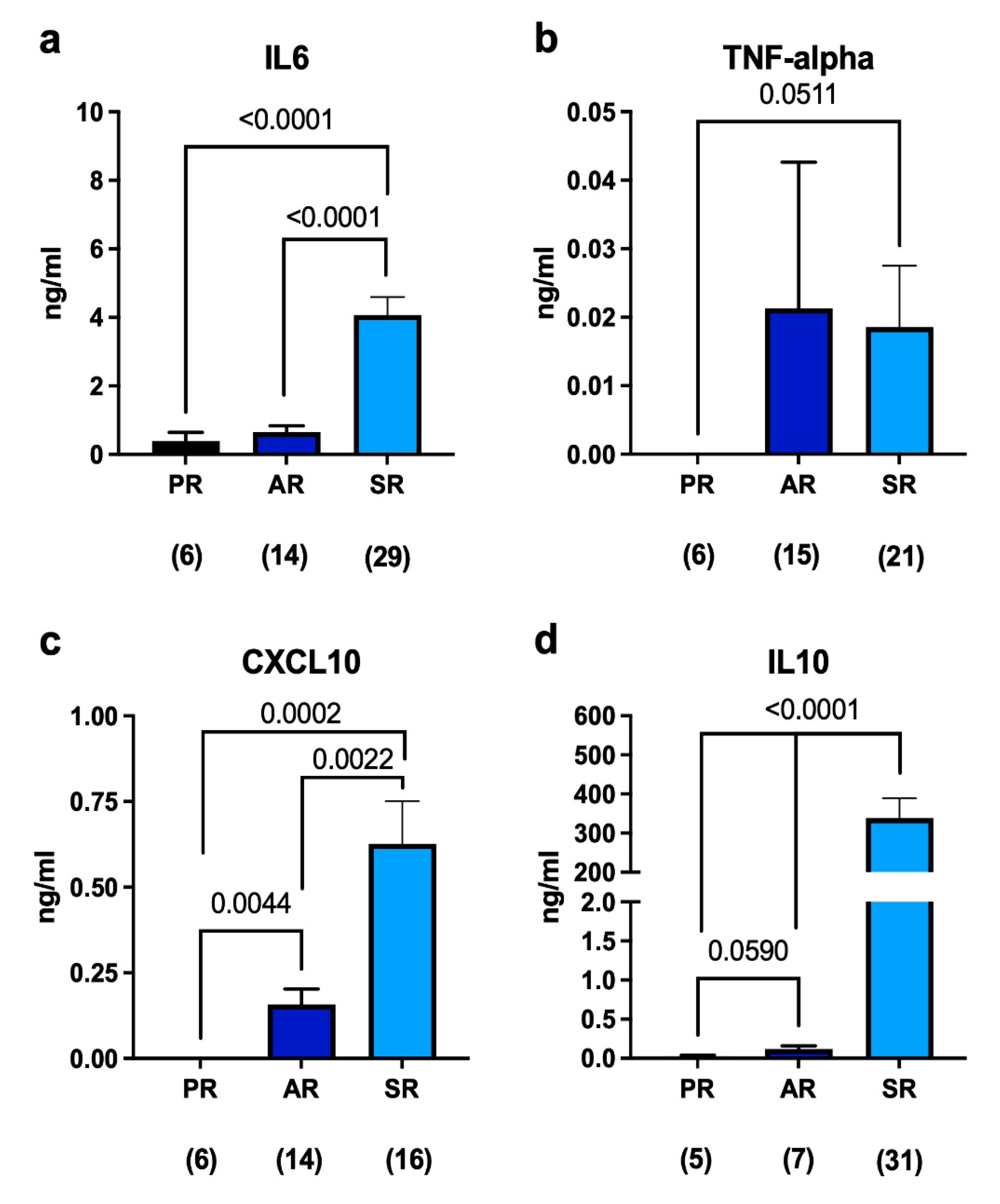
2.1. Immune Response to Joint Replacement
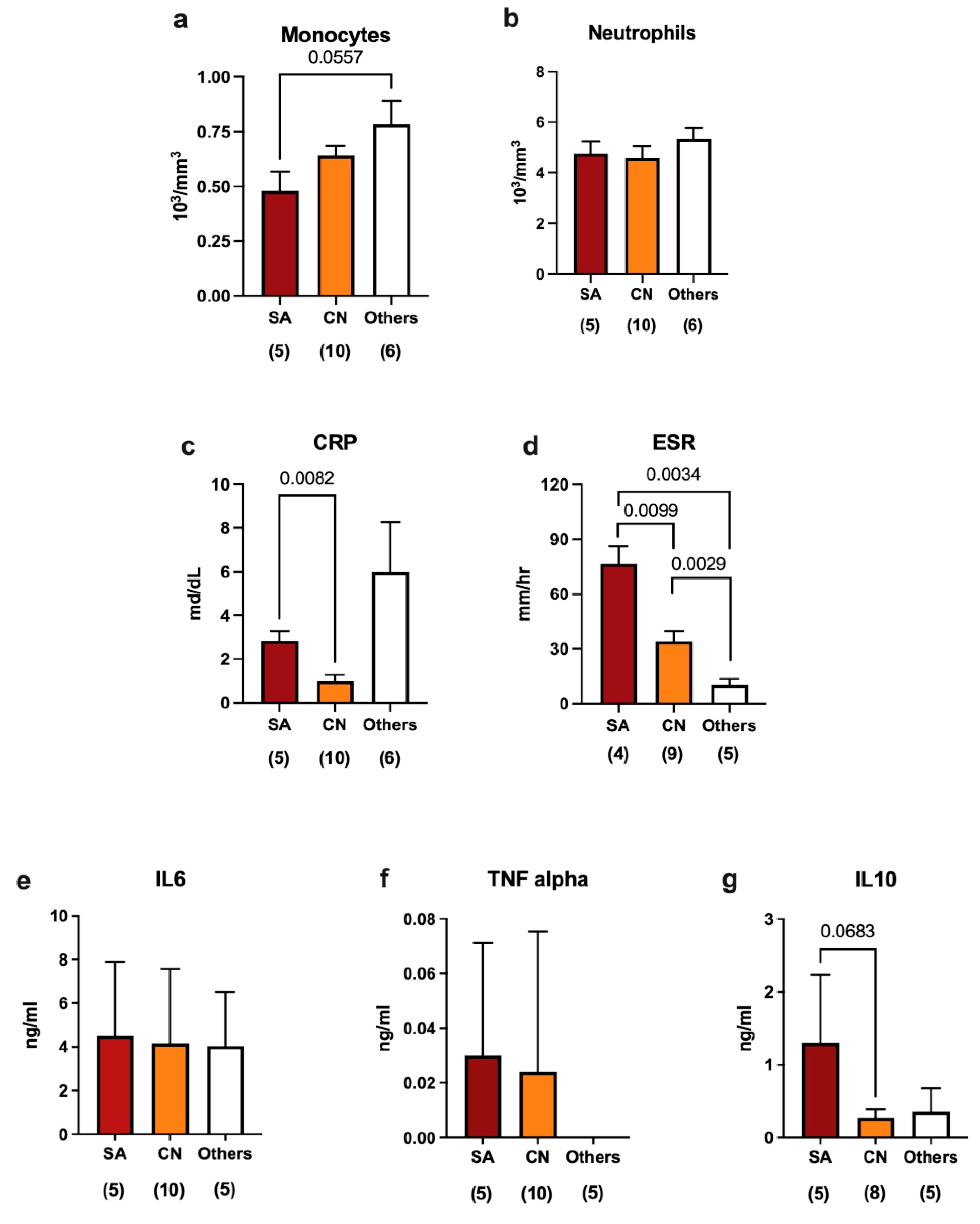
2.2. Immune Response in Septic Revisions
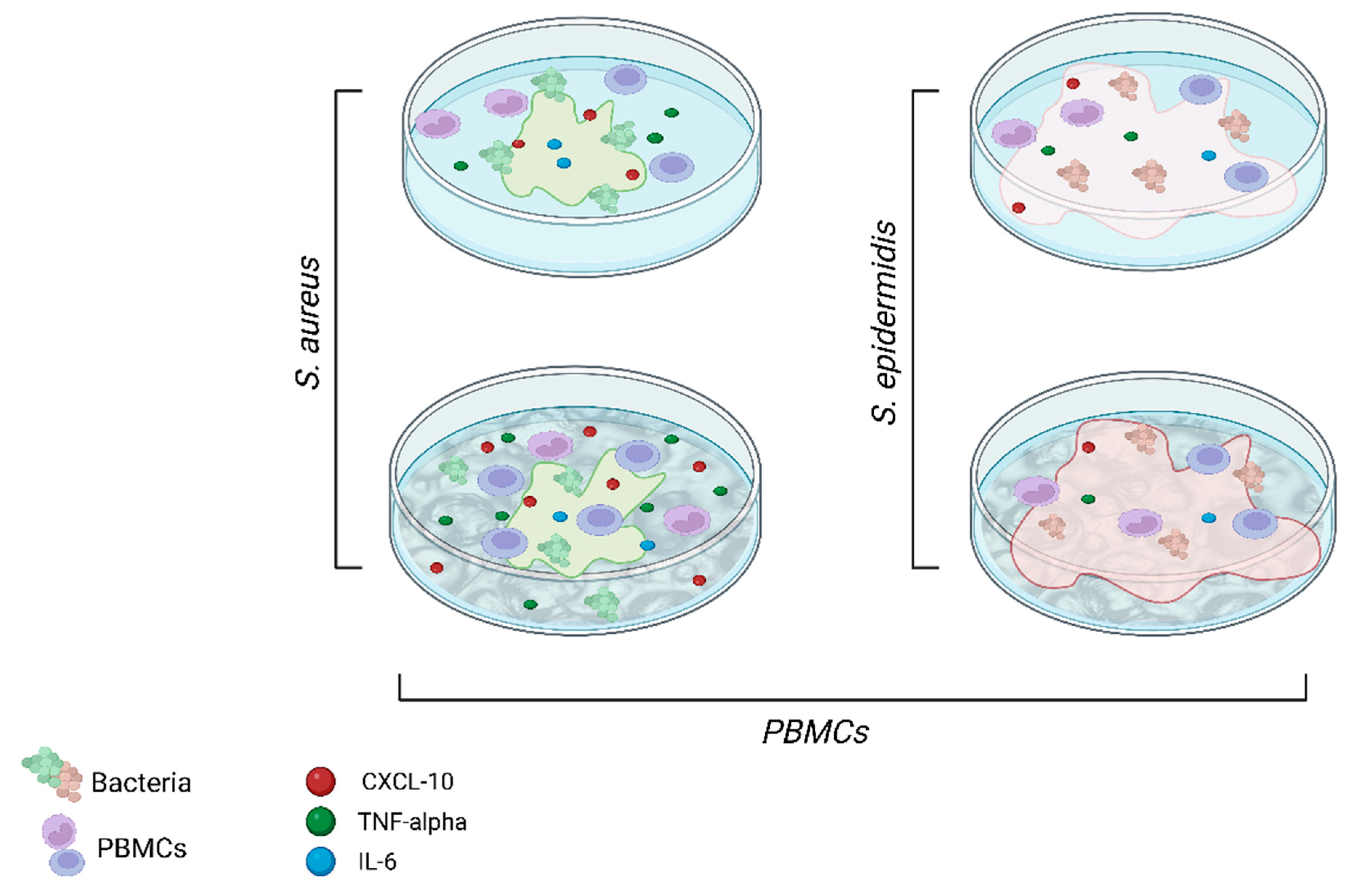
2.3. Pathogenesis Mechanisms in Periprosthetic Joint Infections: An In Vitro Model
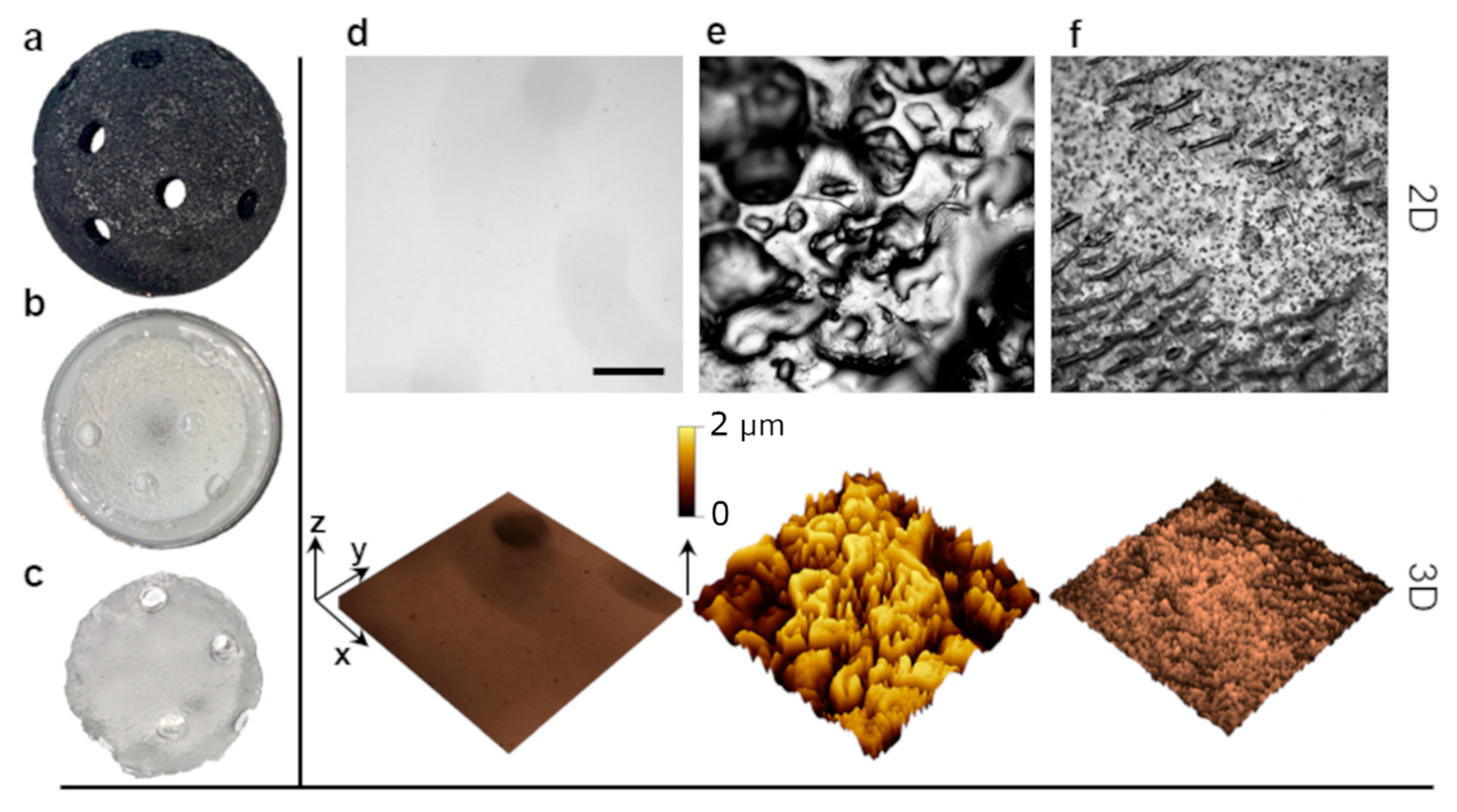
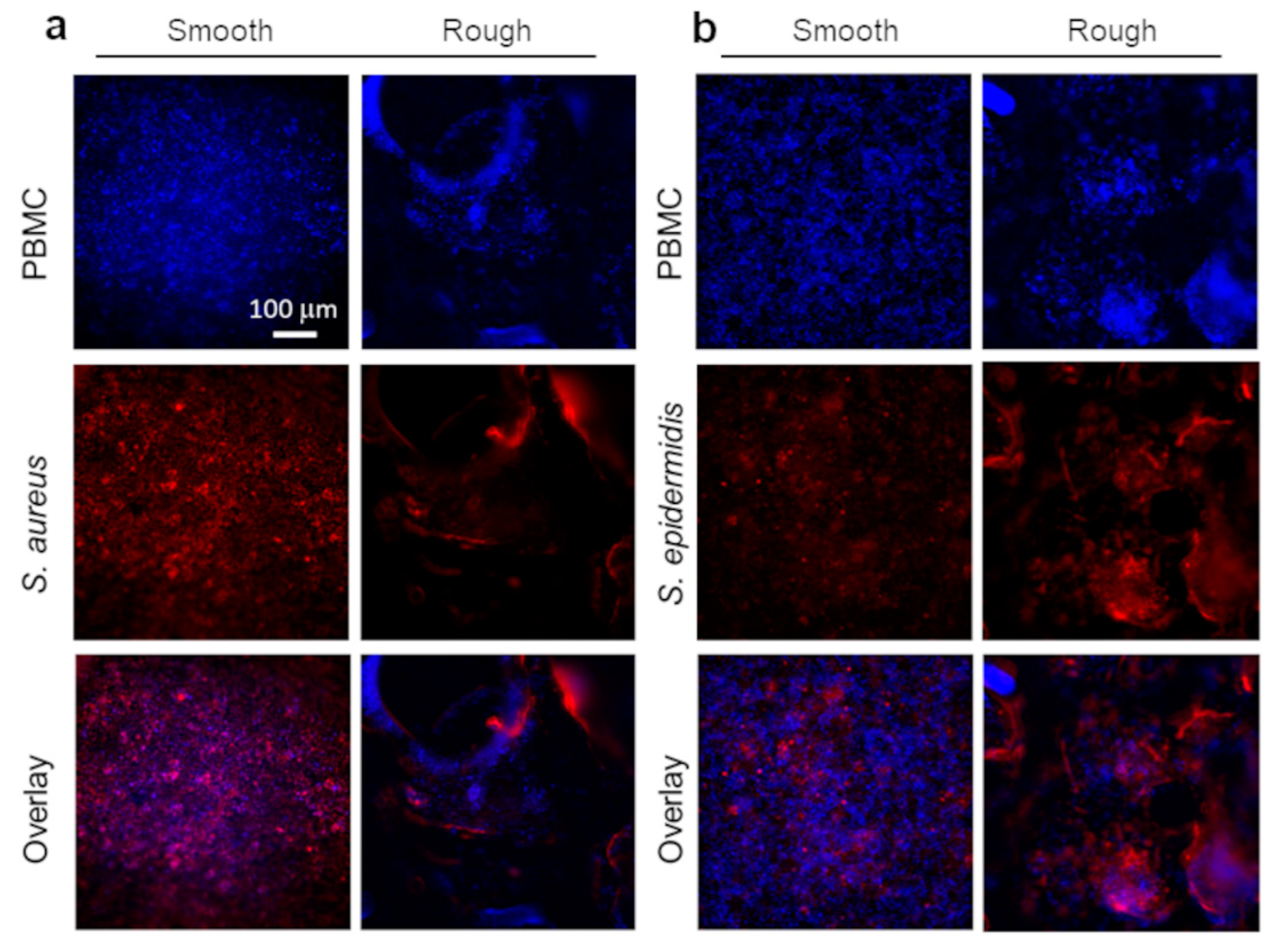
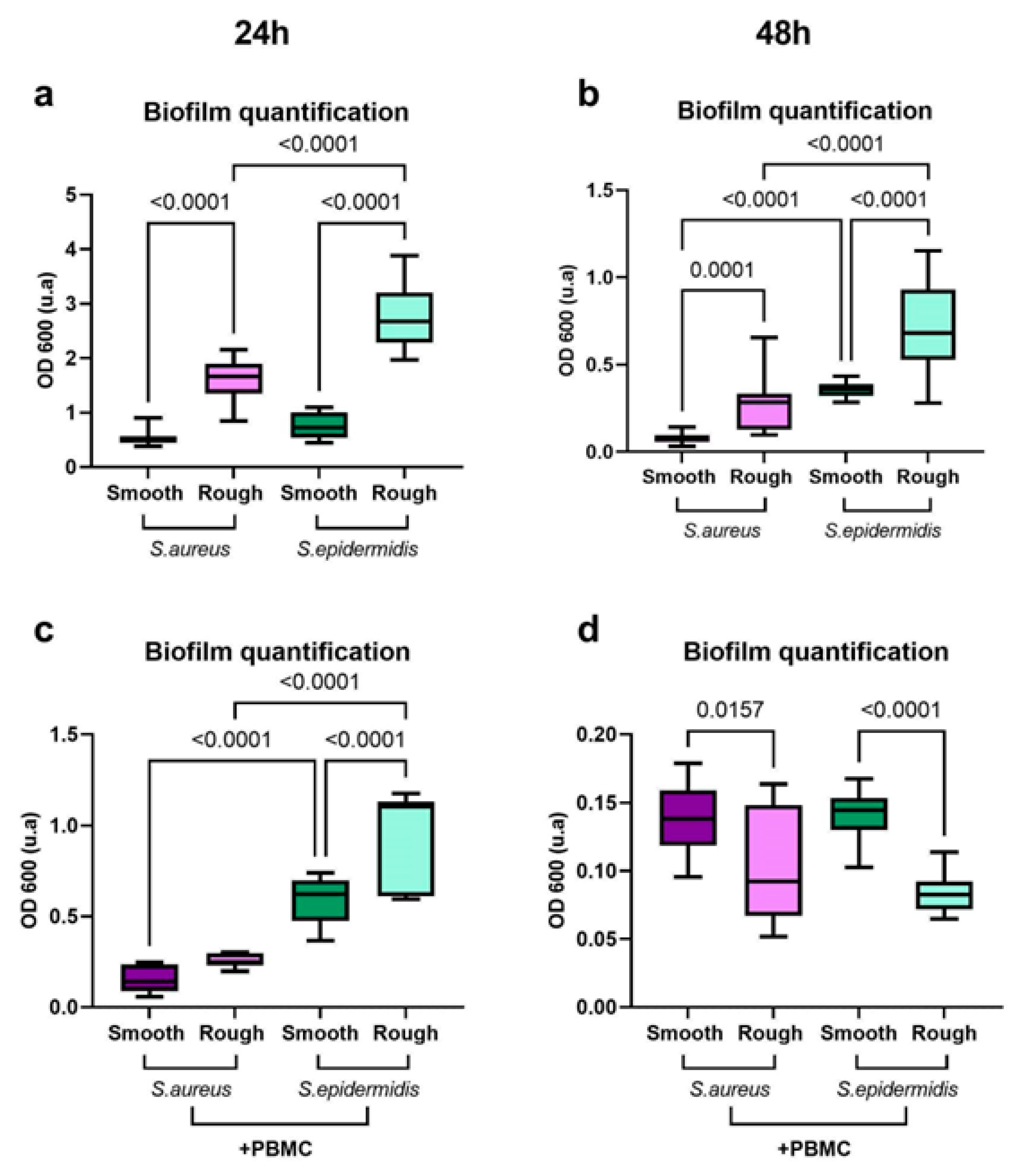
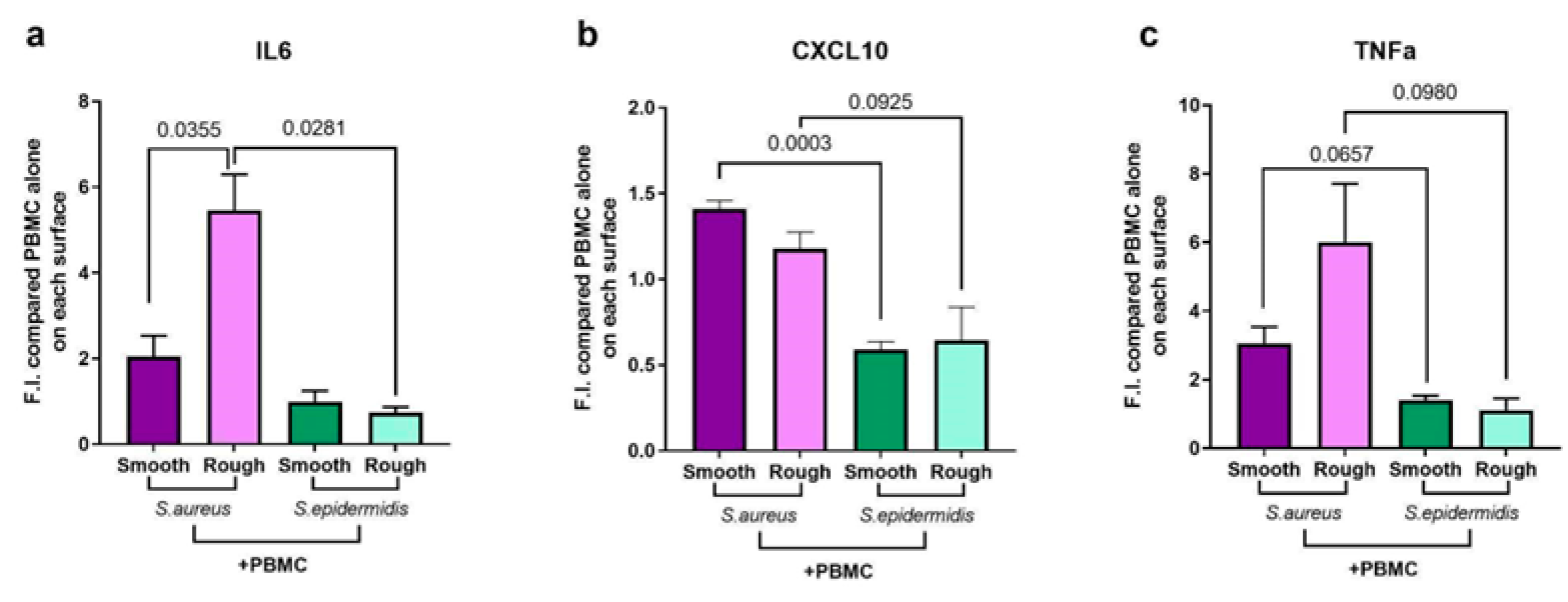
2.4. Surface Topography Modulates Both Bacterial Growth and Immune Response
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Flow Cytometry Experiments
4.3. Model Surface Preparation
4.4. Cell Culture Experiments
4.5. Bacterial Cultures
4.6. In Vitro Model Platform Preparation
4.7. Diagnostic Assay (ELISA)
4.8. Biofilm Growth Assay (Crystal Violet)
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nodzo, S.R.; Boyle, K.K.; Spiro, S.; Nocon, A.A.; Miller, A.O.; Westrich, G.H. Success Rates, Characteristics, and Costs of Articulating Antibiotic Spacers for Total Knee Periprosthetic Joint Infection. Knee 2017, 24, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Patel, R. Periprosthetic Joint Infection. N. Engl. J. Med. 2023, 388, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, W.; Trampuz, A.; Ochsner, P.E. Prosthetic-Joint Infections. N. Engl. J. Med. 2004, 351, 1645–1654. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, W.F.; Silva, P.M.S.; Silva, R.C.S.; Silva, G.M.M.; Machado, G.; Coelho, L.C.B.B.; Correia, M.T.S. Staphylococcus Aureus and Staphylococcus Epidermidis Infections on Implants. J. Hosp. Infect. 2018, 98, 111–117. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant Infections: Adhesion, Biofilm Formation and Immune Evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef]
- Ettinger, M.; Calliess, T.; Kielstein, J.T.; Sibai, J.; Brückner, T.; Lichtinghagen, R.; Windhagen, H.; Lukasz, A. Circulating Biomarkers for Discrimination Between Aseptic Joint Failure, Low-Grade Infection, and High-Grade Septic Failure. Clin. Infect. Dis. 2015, 61, 332–341. [Google Scholar] [CrossRef]
- Gbejuade, H.O.; Lovering, A.M.; Webb, J.C. The Role of Microbial Biofilms in Prosthetic Joint Infections. Acta Orthop. 2014, 86, 147–158. [Google Scholar] [CrossRef]
- Springer, B.D. The Diagnosis of Periprosthetic Joint Infection. J. Arthroplast. 2015, 30, 908–911. [Google Scholar] [CrossRef]
- Izakovicova, P.; Borens, O.; Trampuz, A. Periprosthetic Joint Infection: Current Concepts and Outlook. Efort Open Rev. 2019, 4, 482–494. [Google Scholar] [CrossRef]
- Shoji, M.; Chen, A. Biofilms in Periprosthetic Joint Infections: A Review of Diagnostic Modalities, Current Treatments, and Future Directions. J. Knee Surg. 2020, 33, 119–131. [Google Scholar] [CrossRef]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial Biofilms: Development, Dispersal, and Therapeutic Strategies in the Dawn of the Postantibiotic Era. CSH Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef]
- Secchi, E.; Savorana, G.; Vitale, A.; Eberl, L.; Stocker, R.; Rusconi, R. The Structural Role of Bacterial EDNA in the Formation of Biofilm Streamers. Proc. Natl. Acad. Sci. USA 2022, 119, e2113723119. [Google Scholar] [CrossRef] [PubMed]
- Rusconi, R.; Lecuyer, S.; Autrusson, N.; Guglielmini, L.; Stone, H.A. Secondary Flow as a Mechanism for the Formation of Biofilm Streamers. Biophys. J. 2011, 100, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Ashok, N.; Bauer, C.E. Evidence of Defined Temporal Expression Patterns That Lead a Gram-Negative Cell out of Dormancy. PLoS Genet. 2020, 16, e1008660. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.; Patel, S.K.S.; Kharga, K.; Kumar, R.; Kumar, P.; Pandohee, J.; Kulshresha, S.; Harjai, K.; Chhibber, S. Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria. Molecules 2022, 27, 7584. [Google Scholar] [CrossRef]
- Khajanchi, B.K.; Kirtley, M.L.; Brackman, S.M.; Chopra, A.K. Immunomodulatory and Protective Roles of Quorum-Sensing Signaling Molecules N-Acyl Homoserine Lactones during Infection of Mice with Aeromonas Hydrophila. Infect. Immun. 2011, 79, 2646–2657. [Google Scholar] [CrossRef]
- Scherr, T.D.; Heim, C.E.; Morrison, J.M.; Kielian, T. Hiding in Plain Sight: Interplay between Staphylococcal Biofilms and Host Immunity. Front. Immunol. 2014, 5, 37. [Google Scholar] [CrossRef]
- Thurlow, L.R.; Hanke, M.L.; Fritz, T.; Angle, A.; Aldrich, A.; Williams, S.H.; Engebretsen, I.L.; Bayles, K.W.; Horswill, A.R.; Kielian, T. Staphylococcus Aureus Biofilms Prevent Macrophage Phagocytosis and Attenuate Inflammation In Vivo. J. Immunol. 2011, 186, 6585–6596. [Google Scholar] [CrossRef]
- Caldara, M.; Belgiovine, C.; Secchi, E.; Rusconi, R. Environmental, Microbiological, and Immunological Features of Bacterial Biofilms Associated with Implanted Medical Devices. Clin. Microbiol. Rev. 2022, 35, e00221-20. [Google Scholar] [CrossRef]
- Kharga, K.; Kumar, L.; Patel, S.K.S. Recent Advances in Monoclonal Antibody-Based Approaches in the Management of Bacterial Sepsis. Biomedicines 2023, 11, 765. [Google Scholar] [CrossRef]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign Body Reaction to Biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef]
- Sheikh, Z.; Brooks, P.J.; Barzilay, O.; Fine, N.; Glogauer, M. Macrophages, Foreign Body Giant Cells and Their Response to Implantable Biomaterials. Materials 2015, 8, 5671–5701. [Google Scholar] [CrossRef] [PubMed]
- Frazão, L.P.; Castro, J.V.d.; Neves, N.M. Biomimicked Biomaterials, Advances in Tissue Engineering and Regenerative Medicine. Adv. Exp. Med. Biol. 2020, 1250, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Doloff, J.C.; Veiseh, O.; Mezerville, R.d.; Sforza, M.; Perry, T.A.; Haupt, J.; Jamiel, M.; Chambers, C.; Nash, A.; Aghlara-Fotovat, S.; et al. The Surface Topography of Silicone Breast Implants Mediates the Foreign Body Response in Mice, Rabbits and Humans. Nat. Biomed. Eng. 2021, 5, 1115–1130. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Feng, G.; Moraru, C.I. Micro- and Nanotopography Sensitive Bacterial Attachment Mechanisms: A Review. Front. Microbiol. 2019, 10, 191. [Google Scholar] [CrossRef]
- Secchi, E.; Vitale, A.; Miño, G.L.; Kantsler, V.; Eberl, L.; Rusconi, R.; Stocker, R. The Effect of Flow on Swimming Bacteria Controls the Initial Colonization of Curved Surfaces. Nat. Commun. 2020, 11, 2851. [Google Scholar] [CrossRef]
- Murray, D.W. Cemented Femoral Fixation. Bone Jt. J. 2013, 95-B, 51–52. [Google Scholar] [CrossRef]
- Gandhi, R.; Tsvetkov, D.; Davey, J.R.; Mahomed, N.N. Survival and Clinical Function of Cemented and Uncemented Prostheses in Total Knee Replacement. Bone Jt. J. 2009, 91-B, 889–895. [Google Scholar] [CrossRef]
- Maggs, J.; Wilson, M. The Relative Merits of Cemented and Uncemented Prostheses in Total Hip Arthroplasty. Indian J. Orthop. 2017, 51, 377–385. [Google Scholar] [CrossRef]
- Levine, B. A New Era in Porous Metals: Applications in Orthopaedics. Adv. Eng. Mater. 2008, 10, 788–792. [Google Scholar] [CrossRef]
- Matharu, G.S.; Judge, A.; Murray, D.W.; Pandit, H.G. Do Trabecular Metal Acetabular Components Reduce the Risk of Rerevision After Revision THA Performed for Periprosthetic Joint Infection? A Study Using the NJR Data Set. Clin. Orthop. Relat. Res. 2018, 477, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-Related Inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Eberl, G.; Colonna, M.; Santo, J.P.D.; McKenzie, A.N.J. Innate Lymphoid Cells. Innate Lymphoid Cells: A New Paradigm in Immunology. Science 2015, 348, aaa6566. [Google Scholar] [CrossRef] [PubMed]
- Loppini, M.; Pisano, A.; Maio, M.D.; Camera, F.L.; Casana, M.; Grappiolo, G. Outcomes of Patients with Unexpected Diagnosis of Infection at Total Hip or Total Knee Arthroplasty Revisions. Int. Orthop. 2021, 45, 2791–2796. [Google Scholar] [CrossRef]
- Pellegrino, L.; Kriem, L.S.; Robles, E.S.J.; Cabral, J.T. Microbial Response to Micrometer-Scale Multiaxial Wrinkled Surfaces. ACS Appl. Mater. Interfaces 2022, 14, 31463–31473. [Google Scholar] [CrossRef]
- Bongaerts, J.H.H.; Fourtouni, K.; Stokes, J.R. Soft-Tribology: Lubrication in a Compliant PDMS–PDMS Contact. Tribol. Int. 2007, 40, 1531–1542. [Google Scholar] [CrossRef]
- Schildhauer, T.A.; Peter, E.; Muhr, G.; Köller, M. Activation of Human Leukocytes on Tantalum Trabecular Metal in Comparison to Commonly Used Orthopedic Metal Implant Materials. J. Biomed. Mater. Res. A 2009, 88, 332–341. [Google Scholar] [CrossRef]
- Crémet, L.; Corvec, S.; Batard, E.; Auger, M.; Lopez, I.; Pagniez, F.; Dauvergne, S.; Caroff, N. Comparison of Three Methods to Study Biofilm Formation by Clinical Strains of Escherichia Coli. Diagn. Microbiol. Infect. Dis. 2013, 75, 252–255. [Google Scholar] [CrossRef]
- Hamdi, M. Association Between Breast Implant-Associated Anaplastic Large Cell Lymphoma (BIA-ALCL) Risk and Polyurethane Breast Implants: Clinical Evidence and European Perspective. Aesthet. Surg. J. 2019, 39, S49–S54. [Google Scholar] [CrossRef]
- Kellogg, B.C.; Hiro, M.E.; Payne, W.G. Implant-Associated Anaplastic Large Cell Lymphoma: Beyond Breast Prostheses. Ann. Plast. Surg. 2013, 73, 461–464. [Google Scholar] [CrossRef]
- Vivacqua, A.; Kerwin, K.J.; Tubbs, R.; Roselli, E.E. Lymphoma of Prosthetic Aortic Graft Presenting as Recurrent Embolization. Ann. Thorac. Surg. 2015, 99, 306–307. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.; Vasconcelos, D.P.; Sousa, D.M.; Sousa, B.; Conceição, F.; Neto, E.; Lamghari, M.; Alves, C.J. The Mechanisms Underlying the Biological Response to Wear Debris in Periprosthetic Inflammation. Front. Mater. 2020, 7, 274. [Google Scholar] [CrossRef]
- Haak, V.M.; Huang, S.; Panigrahy, D. Debris-Stimulated Tumor Growth: A Pandora’s Box? Cancer Metastasis Rev. 2021, 40, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.; Zhang, J.-F.; Fang, F.-Z. In Vitro Evaluation of Artificial Joints: A Comprehensive Review. Adv. Manuf. 2019, 7, 1–14. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Speziale, P.; Montanaro, L.; Costerton, J.W. Biofilm Formation in Staphylococcus Implant Infections. A Review of Molecular Mechanisms and Implications for Biofilm-Resistant Materials. Biomaterials 2012, 33, 5967–5982. [Google Scholar] [CrossRef] [PubMed]
- Cossarizza, A.; Chang, H.; Radbruch, A.; Abrignani, S.; Addo, R.; Akdis, M.; Andrä, I.; Andreata, F.; Annunziato, F.; Arranz, E.; et al. Guidelines for the Use of Flow Cytometry and Cell Sorting in Immunological Studies (Third Edition). Eur. J. Immunol. 2021, 51, 2708–3145. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of patients | 74 |
| Septic revision | 50 |
| Aseptic revision | 18 |
| Primary replacement | 6 |
| Age median (IQR) | 63 (33.5–90.3) |
| Septic revision | 71 (33.5–90.3) |
| Aseptic revision | 64 (53–86) |
| Primary replacement | 64.5 (57–84) |
| Sex (female) | 29 (39%) |
| Septic revision | 18 |
| Aseptic revision | 10 |
| Primary replacement | 1 |
| Hip | 61 (82%) |
| Septic revision | 31 |
| Aseptic revision | 15 |
| Primary replacement | 5 |
| Knee | 13 (18%) |
| Septic revision | 10 |
| Aseptic revision | 2 |
| Primary replacement | 1 |
| Etiology | |
| Staphylococcus aureus | 6 (14%) 1 |
| Coagulase Negative | 11 (26%) 1 |
| Staphylococcus epidermidis | 7 (17%) 1 |
| Others | 25 (60%) 1 |
| Total SR | SA | SE | SE/SA | |
|---|---|---|---|---|
| Cemented | 25 | 3 | 6 | 2 |
| Uncemented | 111 | 8 | 22 | 2.75 |
| Ra (µm) | Rq (µm) | Āmax (µm) | |
|---|---|---|---|
| Smooth | 0.025 ± 0.008 | 0.042 ± 0.008 | 0.090 ± 0.010 |
| Rough | 2.240 ± 0.070 | 2.980 ± 0.070 | 3100 ± 0.095 |
| Cement | 0.110 ± 0.020 | 0.150 ± 0.020 | 0.250 ± 0.050 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belgiovine, C.; Pellegrino, L.; Bulgarelli, A.; Lauta, F.C.; Di Claudio, A.; Ciceri, R.; Cancellara, A.; Calcaterra, F.; Mavilio, D.; Grappiolo, G.; et al. Interaction of Bacteria, Immune Cells, and Surface Topography in Periprosthetic Joint Infections. Int. J. Mol. Sci. 2023, 24, 9028. https://doi.org/10.3390/ijms24109028
Belgiovine C, Pellegrino L, Bulgarelli A, Lauta FC, Di Claudio A, Ciceri R, Cancellara A, Calcaterra F, Mavilio D, Grappiolo G, et al. Interaction of Bacteria, Immune Cells, and Surface Topography in Periprosthetic Joint Infections. International Journal of Molecular Sciences. 2023; 24(10):9028. https://doi.org/10.3390/ijms24109028
Chicago/Turabian StyleBelgiovine, Cristina, Luca Pellegrino, Alberto Bulgarelli, Francesca Cecilia Lauta, Alessia Di Claudio, Roberta Ciceri, Assunta Cancellara, Francesca Calcaterra, Domenico Mavilio, Guido Grappiolo, and et al. 2023. "Interaction of Bacteria, Immune Cells, and Surface Topography in Periprosthetic Joint Infections" International Journal of Molecular Sciences 24, no. 10: 9028. https://doi.org/10.3390/ijms24109028
APA StyleBelgiovine, C., Pellegrino, L., Bulgarelli, A., Lauta, F. C., Di Claudio, A., Ciceri, R., Cancellara, A., Calcaterra, F., Mavilio, D., Grappiolo, G., Chiappetta, K., Loppini, M., & Rusconi, R. (2023). Interaction of Bacteria, Immune Cells, and Surface Topography in Periprosthetic Joint Infections. International Journal of Molecular Sciences, 24(10), 9028. https://doi.org/10.3390/ijms24109028