
Influence of Polyphenols on Adipose Tissue: Sirtuins as Pivotal Players in the Browning Process

 , , ,
, , , 

Abstract
:1. Introduction
2. Pathological Conditions of Adipose Tissue
3. Brown Adipose Tissue and Its Implication in Metabolic Processes
3.1. BAT
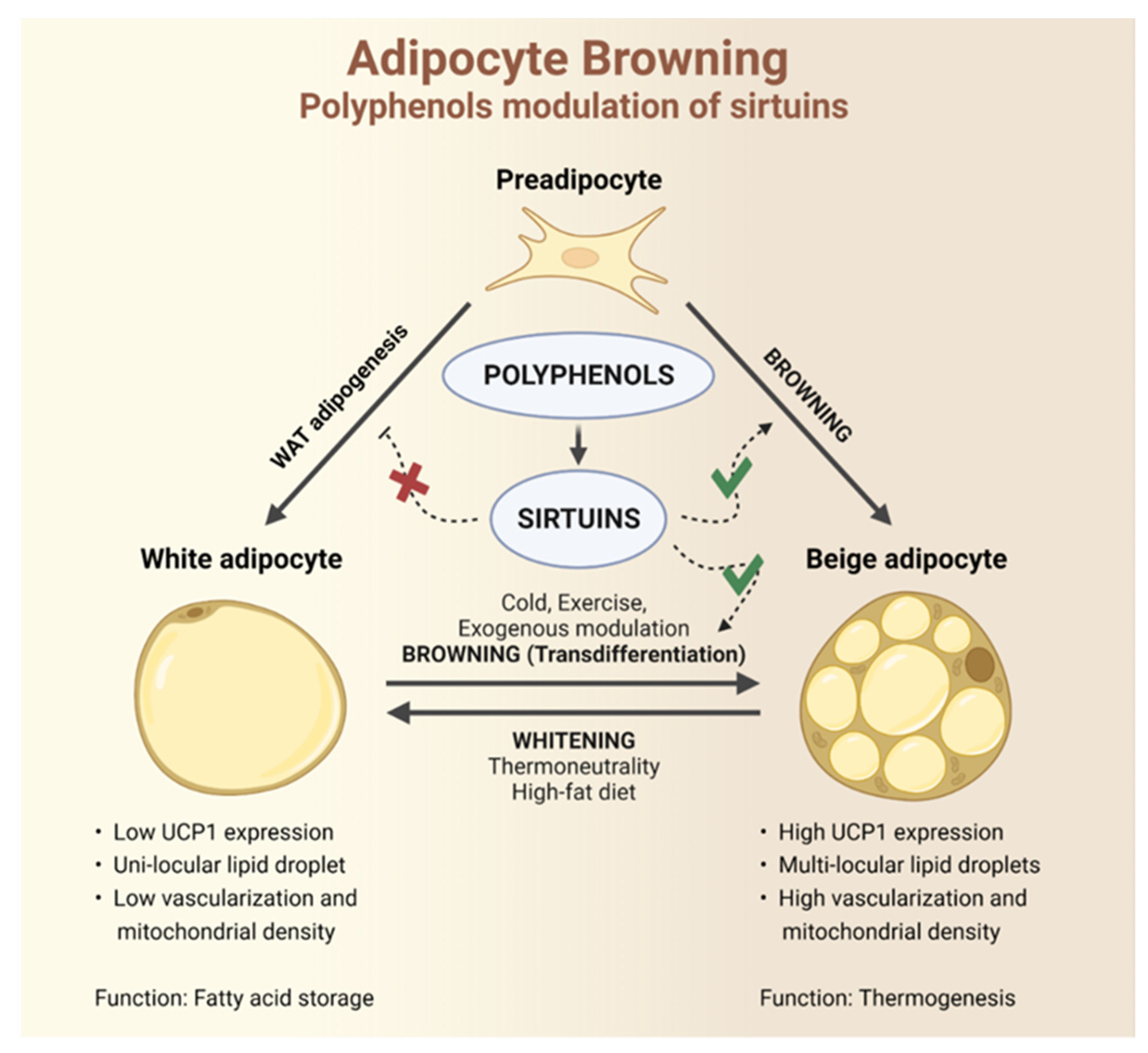
3.2. Browning
4. Polyphenols and Sirtuins Interaction as a Tool for Browning Stimulation
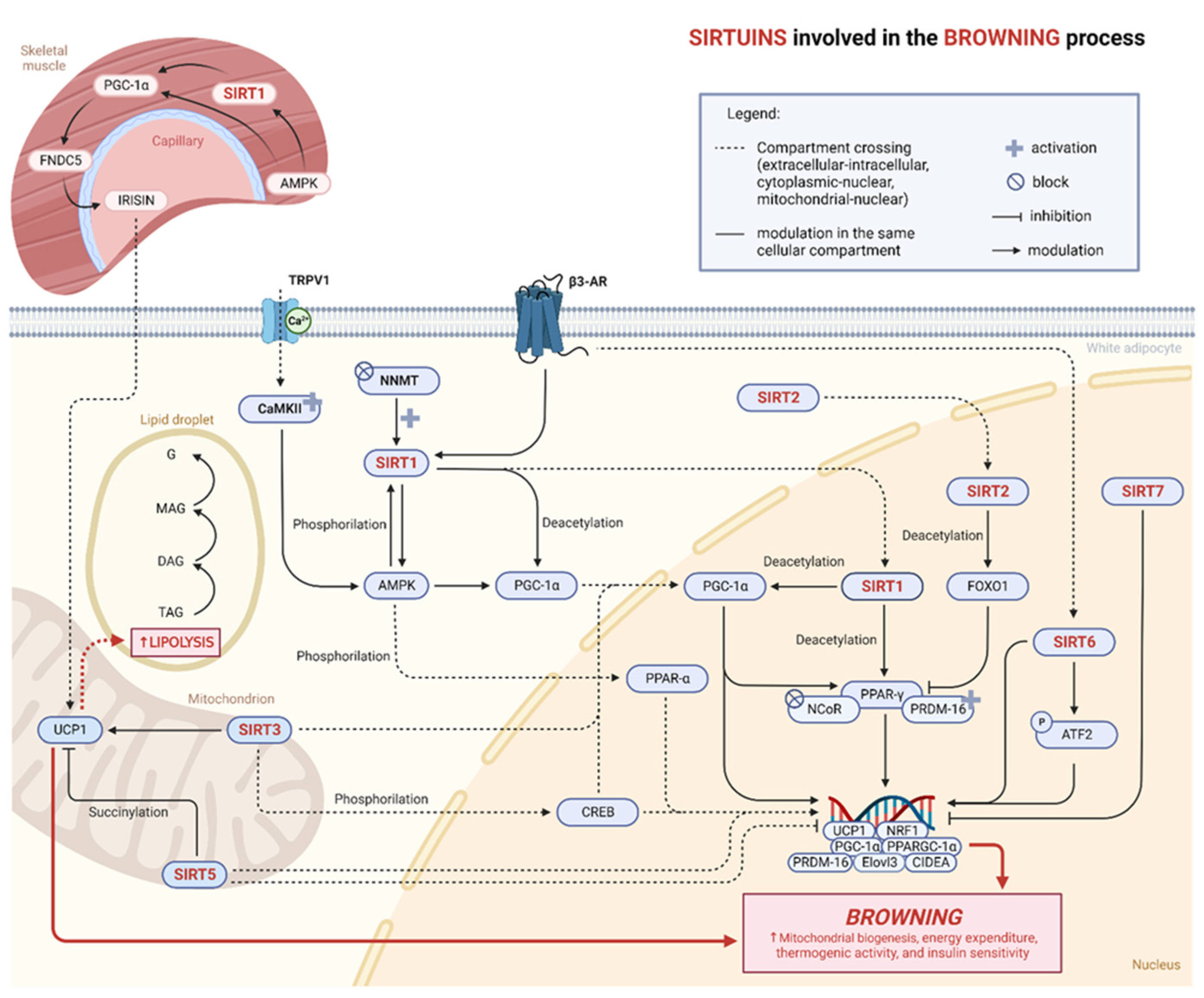
4.1. Sirtuins in Adipose Tissue
4.2. Sirtuins and Browning
4.2.1. SIRT1
4.2.2. SIRT2
4.2.3. SIRT3
4.2.4. SIRT5
4.2.5. SIRT6
4.2.6. SIRT7
5. Dietary Polyphenols and the Browning Process: Evidence from Pre-Clinical Studies
5.1. Polyphenols and Browning
5.2. SIRT1-Mediated Browning Effects of Polyphenols
6. Polyphenols and Human Obesity: The Still Unconsidered Role of Sirtuins
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cinti, S. The endocrine adipose organ. Rev. Endocr. Metab. Disord. 2022, 23, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Emont, M.P.; Jacobs, C.; Essene, A.L.; Pant, D.; Tenen, D.; Colleluori, G.; Di Vincenzo, A.; Jørgensen, A.M.; Dashti, H.; Stefek, A. A single-cell atlas of human and mouse white adipose tissue. Nature 2022, 603, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Arner, P. Fat tissue growth and development in humans. Recent Res. Nutr. Growth 2018, 89, 37–45. [Google Scholar]
- Sakers, A.; De Siqueira, M.K.; Seale, P.; Villanueva, C.J. Adipose-tissue plasticity in health and disease. Cell 2022, 185, 419–446. [Google Scholar] [CrossRef] [PubMed]
- Zwick, R.K.; Guerrero-Juarez, C.F.; Horsley, V.; Plikus, M.V. Anatomical, physiological, and functional diversity of adipose tissue. Cell Metab. 2018, 27, 68–83. [Google Scholar] [CrossRef]
- Frühbeck, G. Overview of adipose tissue and its role in obesity and metabolic disorders. In Adipose Tissue Protocols; Humana Press: Totowa, NJ, USA, 2008; pp. 1–22. [Google Scholar]
- Kwon, H.; Pessin, J.E. Adipokines mediate inflammation and insulin resistance. Front. Endocrinol. 2013, 4, 71. [Google Scholar] [CrossRef]
- Ahima, R.S.; Lazar, M.A. Adipokines and the peripheral and neural control of energy balance. Mol. Endocrinol. 2008, 22, 1023–1031. [Google Scholar] [CrossRef]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Meyer, L.K.; Ciaraldi, T.P.; Henry, R.R.; Wittgrove, A.C.; Phillips, S.A. Adipose tissue depot and cell size dependency of adiponectin synthesis and secretion in human obesity. Adipocyte 2013, 2, 217–226. [Google Scholar] [CrossRef]
- Xiao, L.; Yang, X.; Lin, Y.; Li, S.; Jiang, J.; Qian, S.; Tang, Q.; He, R.; Li, X. Large adipocytes function as antigen-presenting cells to activate CD4+ T cells via upregulating MHCII in obesity. Int. J. Obes. 2016, 40, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Crewe, C.; An, Y.A.; Scherer, P.E. The ominous triad of adipose tissue dysfunction: Inflammation, fibrosis, and impaired angiogenesis. J. Clin. Investig. 2017, 127, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.-C.; Hsieh, P.-S. The role of adipocyte hypertrophy and hypoxia in the development of obesity-associated adipose tissue inflammation and insulin resistance. In Adiposity Omics Molecular Understanding; IntechOpen: London, UK, 2017. [Google Scholar]
- Vishvanath, L.; Gupta, R.K. Contribution of adipogenesis to healthy adipose tissue expansion in obesity. J. Clin. Investig. 2019, 129, 4022–4031. [Google Scholar] [CrossRef]
- Pasarica, M.; Sereda, O.R.; Redman, L.M.; Albarado, D.C.; Hymel, D.T.; Roan, L.E.; Rood, J.C.; Burk, D.H.; Smith, S.R. Reduced adipose tissue oxygenation in human obesity: Evidence for rarefaction, macrophage chemotaxis, and inflammation without an angiogenic response. Diabetes 2009, 58, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Fujisaka, S.; Usui, I.; Ikutani, M.; Aminuddin, A.; Takikawa, A.; Tsuneyama, K.; Mahmood, A.; Goda, N.; Nagai, Y.; Takatsu, K.; et al. Adipose tissue hypoxia induces inflammatory M1 polarity of macrophages in an HIF-1α-dependent and HIF-1α-independent manner in obese mice. Diabetologia 2013, 56, 1403–1412. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-α: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef]
- Patsouris, D.; Li, P.-P.; Thapar, D.; Chapman, J.; Olefsky, J.M.; Neels, J.G. Ablation of CD11c-positive cells normalizes insulin sensitivity in obese insulin resistant animals. Cell Metab. 2008, 8, 301–309. [Google Scholar] [CrossRef]
- Marcelin, G.; Ferreira, A.; Liu, Y.; Atlan, M.; Aron-Wisnewsky, J.; Pelloux, V.; Botbol, Y.; Ambrosini, M.; Fradet, M.; Rouault, C.; et al. A PDGFRα-mediated switch toward CD9high adipocyte progenitors controls obesity-induced adipose tissue fibrosis. Cell Metab. 2017, 25, 673–685. [Google Scholar] [CrossRef]
- Sun, K.; Tordjman, J.; Clément, K.; Scherer, P.E. Fibrosis and adipose tissue dysfunction. Cell Metab. 2013, 18, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Pellegrinelli, V.; Heuvingh, J.; Du Roure, O.; Rouault, C.; Devulder, A.; Klein, C.; Lacasa, M.; Clément, E.; Lacasa, D.; Clément, K. Human adipocyte function is impacted by mechanical cues. J. Pathol. 2014, 233, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Ayala, J.E.; Lee-Young, R.S.; Zhang, Z.; James, F.D.; Neufer, P.D.; Pozzi, A.; Zutter, M.M.; Wasserman, D.H. Diet-induced muscle insulin resistance is associated with extracellular matrix remodeling and interaction with integrin α2β1 in mice. Diabetes 2011, 60, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Weyer, C.; Foley, J.; Bogardus, C.; Tataranni, P.; Pratley, R. Enlarged subcutaneous abdominal adipocyte size, but not obesity itself, predicts type II diabetes independent of insulin resistance. Diabetologia 2000, 43, 1498–1506. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Kajimura, S. Metabolic adaptation and maladaptation in adipose tissue. Nat. Metab. 2019, 1, 189–200. [Google Scholar] [CrossRef]
- Brownstein, A.J.; Veliova, M.; Acin-Perez, R.; Liesa, M.; Shirihai, O.S. ATP-consuming futile cycles as energy dissipating mechanisms to counteract obesity. Rev. Endocr. Metab. Disord. 2022, 23, 121–131. [Google Scholar] [CrossRef]
- Fedorenko, A.; Lishko, P.V.; Kirichok, Y. Mechanism of fatty-acid-dependent UCP1 uncoupling in brown fat mitochondria. Cell 2012, 151, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Buffolo, M.; Pires, K.M.; Pei, S.; Scherer, P.E.; Boudina, S. Adipocyte-specific deletion of manganese superoxide dismutase protects from diet-induced obesity through increased mitochondrial uncoupling and biogenesis. Diabetes 2016, 65, 2639–2651. [Google Scholar] [CrossRef]
- Gesta, S.; Tseng, Y.-H.; Kahn, C.R. Developmental origin of fat: Tracking obesity to its source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef]
- Leitner, B.P.; Huang, S.; Brychta, R.J.; Duckworth, C.J.; Baskin, A.S.; McGehee, S.; Tal, I.; Dieckmann, W.; Gupta, G.; Kolodny, G.M.; et al. Mapping of human brown adipose tissue in lean and obese young men. Proc. Natl. Acad. Sci. USA 2017, 114, 8649–8654. [Google Scholar] [CrossRef]
- Pfannenberg, C.; Werner, M.K.; Ripkens, S.; Stef, I.; Deckert, A.; Schmadl, M.; Reimold, M.; Häring, H.-U.; Claussen, C.D.; Stefan, N. Impact of age on the relationships of brown adipose tissue with sex and adiposity in humans. Diabetes 2010, 59, 1789–1793. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.-H.; Doria, A.; et al. Identification and importance of brown adipose tissue in adult humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Yeung, H.W.; Grewal, R.K.; Gonen, M.; Schöder, H.; Larson, S.M. Patterns of 18F-FDG uptake in adipose tissue and muscle: A potential source of false-positives for PET. J. Nucl. Med. 2003, 44, 1789–1796. [Google Scholar] [PubMed]
- Truong, M.T.; Erasmus, J.J.; Munden, R.F.; Marom, E.M.; Sabloff, B.S.; Gladish, G.W.; Podoloff, D.A.; Macapinlac, H.A. Focal FDG uptake in mediastinal brown fat mimicking malignancy: A potential pitfall resolved on PET/CT. Am. J. Roentgenol. 2004, 183, 1127–1132. [Google Scholar] [CrossRef]
- Aloo, S.-O.; Ofosu, F.K.; Kim, N.-H.; Kilonzi, S.M.; Oh, D.-H. Insights on Dietary Polyphenols as Agents against Metabolic Disorders: Obesity as a Target Disease. Antioxidants 2023, 12, 416. [Google Scholar] [CrossRef]
- Ouellet, V.; Routhier-Labadie, A.; Bellemare, W.; Lakhal-Chaieb, L.; Turcotte, E.; Carpentier, A.C.; Richard, D. Outdoor temperature, age, sex, body mass index, and diabetic status determine the prevalence, mass, and glucose-uptake activity of 18F-FDG-detected BAT in humans. J. Clin. Endocrinol. Metab. 2011, 96, 192–199. [Google Scholar] [CrossRef]
- Cinti, S. The adipose organ at a glance. Dis. Model. Mech. 2012, 5, 588–594. [Google Scholar] [CrossRef]
- Bartelt, A.; Heeren, J. Adipose tissue browning and metabolic health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Klingenspor, M.; Herzig, S.; Pfeifer, A. Brown fat develops a brite future. Obes. Facts 2012, 5, 890–896. [Google Scholar] [CrossRef]
- Hammarstedt, A.; Gogg, S.; Hedjazifar, S.; Nerstedt, A.; Smith, U. Impaired adipogenesis and dysfunctional adipose tissue in human hypertrophic obesity. Physiol. Rev. 2018, 98, 1911–1941. [Google Scholar] [CrossRef]
- Sidossis, L.; Kajimura, S. Brown and beige fat in humans: Thermogenic adipocytes that control energy and glucose homeostasis. J. Clin. Investig. 2015, 125, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Seale, P. Control of brown and beige fat development. Nat. Rev. Mol. Cell Biol. 2016, 17, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scime, A.; Devarakonda, S.; Conroe, H.M.; Erdjument-Bromage, H.; et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008, 454, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Schulz, T.J.; Huang, T.L.; Tran, T.T.; Zhang, H.; Townsend, K.L.; Shadrach, J.L.; Cerletti, M.; McDougall, L.E.; Giorgadze, N.; Tchkonia, T.; et al. Identification of inducible brown adipocyte progenitors residing in skeletal muscle and white fat. Proc. Natl. Acad. Sci. USA 2011, 108, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Petkova, A.P.; Mottillo, E.P.; Granneman, J.G. In vivo identification of bipotential adipocyte progenitors recruited by β3-adrenoceptor activation and high-fat feeding. Cell Metab. 2012, 15, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Barbatelli, G.; Murano, I.; Madsen, L.; Hao, Q.; Jimenez, M.; Kristiansen, K.; Giacobino, J.; De Matteis, R.; Cinti, S. The emergence of cold-induced brown adipocytes in mouse white fat depots is determined predominantly by white to brown adipocyte transdifferentiation. Am. J. Physiol. -Endocrinol. Metab. 2010, 298, E1244–E1253. [Google Scholar] [CrossRef]
- Flori, L.; Testai, L.; Calderone, V. The “irisin system”: From biological roles to pharmacological and nutraceutical perspectives. Life Sci. 2021, 267, 118954. [Google Scholar] [CrossRef]
- Rosenwald, M.; Perdikari, A.; Rülicke, T.; Wolfrum, C. Bi-directional interconversion of brite and white adipocytes. Nat. Cell Biol. 2013, 15, 659–667. [Google Scholar] [CrossRef]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose tissue remodeling: Its role in energy metabolism and metabolic disorders. Front. Endocrinol. 2016, 7, 30. [Google Scholar] [CrossRef]
- Wang, S.; Pan, M.-H.; Hung, W.-L.; Tung, Y.-C.; Ho, C.-T. From white to beige adipocytes: Therapeutic potential of dietary molecules against obesity and their molecular mechanisms. Food Funct. 2019, 10, 1263–1279. [Google Scholar] [CrossRef] [PubMed]
- Villarroya, F.; Cereijo, R.; Gavaldà-Navarro, A.; Villarroya, J.; Giralt, M. Inflammation of brown/beige adipose tissues in obesity and metabolic disease. J. Intern. Med. 2018, 284, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; Piragine, E.; Piano, I.; Flori, L.; Da Pozzo, E.; Miragliotta, V.; Pirone, A.; Citi, V.; Di Cesare Mannelli, L.; Brogi, S.; et al. The citrus flavonoid naringenin protects the myocardium from ageing-dependent dysfunction: Potential role of SIRT1. Oxidative Med. Cell. Longev. 2020, 2020, 4650207. [Google Scholar] [CrossRef] [PubMed]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-inflammatory action and mechanisms of resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef]
- Martelli, A.; Flori, L.; Gorica, E.; Piragine, E.; Saviano, A.; Annunziata, G.; Di Minno, M.N.D.; Ciampaglia, R.; Calcaterra, I.; Maione, F. Vascular effects of the polyphenolic nutraceutical supplement taurisolo®: Focus on the protection of the endothelial function. Nutrients 2021, 13, 1540. [Google Scholar] [CrossRef]
- Ungurianu, A.; Zanfirescu, A.; Margină, D. Regulation of Gene Expression through Food—Curcumin as a Sirtuin Activity Modulator. Plants 2022, 11, 1741. [Google Scholar] [CrossRef]
- Dai, H.; Sinclair, D.A.; Ellis, J.L.; Steegborn, C. Sirtuin activators and inhibitors: Promises, achievements, and challenges. Pharmacol. Ther. 2018, 188, 140–154. [Google Scholar] [CrossRef]
- Yessenkyzy, A.; Saliev, T.; Zhanaliyeva, M.; Masoud, A.-R.; Umbayev, B.; Sergazy, S.; Krivykh, E.; Gulyayev, A.; Nurgozhin, T. Polyphenols as caloric-restriction mimetics and autophagy inducers in aging research. Nutrients 2020, 12, 1344. [Google Scholar] [CrossRef]
- Lee, S.-H.; Lee, J.-H.; Lee, H.-Y.; Min, K.-J. Sirtuin signaling in cellular senescence and aging. BMB Rep. 2019, 52, 24–34. [Google Scholar] [CrossRef]
- Ma, J.; Zheng, Y.; Tang, W.; Yan, W.; Nie, H.; Fang, J.; Liu, G. Dietary polyphenols in lipid metabolism: A role of gut microbiome. Anim. Nutr. 2020, 6, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Wang, M.; Qiu, X.; Liu, D.; Jiang, H.; Yang, N.; Xu, R.-M. Structural basis for allosteric, substrate-dependent stimulation of SIRT1 activity by resveratrol. Genes Dev. 2015, 29, 1316–1325. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.-I.; Armstrong, C.M.; Kaeberlein, M.; Guarente, L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature 2000, 403, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Auwerx, J. The role of sirtuins in the control of metabolic homeostasis. Ann. N. Y. Acad. Sci. 2009, 1173, E10–E19. [Google Scholar] [CrossRef]
- Hwang, J.-W.; Yao, H.; Caito, S.; Sundar, I.K.; Rahman, I. Redox regulation of SIRT1 in inflammation and cellular senescence. Free Radic. Biol. Med. 2013, 61, 95–110. [Google Scholar] [CrossRef]
- Gomes, P.; Outeiro, T.F.; Cavadas, C. Emerging role of sirtuin 2 in the regulation of mammalian metabolism. Trends Pharmacol. Sci. 2015, 36, 756–768. [Google Scholar] [CrossRef]
- Zhou, S.; Tang, X.; Chen, H.-Z. Sirtuins and insulin resistance. Front. Endocrinol. 2018, 9, 748. [Google Scholar] [CrossRef]
- Shi, T.; Fan, G.Q.; Xiao, S.D. SIRT3 reduces lipid accumulation via AMPK activation in human hepatic cells. J. Dig. Dis. 2010, 11, 55–62. [Google Scholar] [CrossRef]
- Wu, Y.; Gao, W.-N.; Xue, Y.-N.; Zhang, L.-C.; Zhang, J.-J.; Lu, S.-Y.; Yan, X.-Y.; Yu, H.-M.; Su, J.; Sun, L.-K. SIRT3 aggravates metformin-induced energy stress and apoptosis in ovarian cancer cells. Exp. Cell Res. 2018, 367, 137–149. [Google Scholar] [CrossRef]
- Shi, T.; Wang, F.; Stieren, E.; Tong, Q. SIRT3, a mitochondrial sirtuin deacetylase, regulates mitochondrial function and thermogenesis in brown adipocytes. J. Biol. Chem. 2005, 280, 13560–13567. [Google Scholar] [CrossRef]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- Vachharajani, V.T.; Liu, T.; Wang, X.; Hoth, J.J.; Yoza, B.K.; McCall, C.E. Sirtuins link inflammation and metabolism. J. Immunol. Res. 2016, 2016, 8167273. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Li, M.; Hou, T.; Gao, T.; Zhu, W.-G.; Yang, Y. Sirtuins in glucose and lipid metabolism. Oncotarget 2017, 8, 1845–1859. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lou, R.; Zhou, F.; Li, D.; Peng, C.; Lin, L. Sirtuins: Key players in obesity-associated adipose tissue remodeling. Front. Immunol. 2022, 13, 1068986. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Burk, D.; Gao, Z.; Yin, J.; Zhang, X.; Weng, J.; Ye, J. Angiogenic deficiency and adipose tissue dysfunction are associated with macrophage malfunction in SIRT1−/− mice. Endocrinology 2012, 153, 1706–1716. [Google Scholar] [CrossRef]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; Machado de Oliveira, R.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-γ. Nature 2004, 429, 771–776. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Z.; Ke, L.; Sun, Y.; Li, W.; Feng, X.; Zhu, W.; Chen, S. Resveratrol promotes white adipocytes browning and improves metabolic disorders in Sirt1-dependent manner in mice. FASEB J. 2020, 34, 4527–4539. [Google Scholar] [CrossRef]
- Yoshizaki, T.; Milne, J.C.; Imamura, T.; Schenk, S.; Sonoda, N.; Babendure, J.L.; Lu, J.-C.; Smith, J.J.; Jirousek, M.R.; Olefsky, J.M. SIRT1 exerts anti-inflammatory effects and improves insulin sensitivity in adipocytes. Mol. Cell. Biol. 2009, 29, 1363–1374. [Google Scholar] [CrossRef]
- Sun, Y.; Li, J.; Xiao, N.; Wang, M.; Kou, J.; Qi, L.; Huang, F.; Liu, B.; Liu, K. Pharmacological activation of AMPK ameliorates perivascular adipose/endothelial dysfunction in a manner interdependent on AMPK and SIRT1. Pharmacol. Res. 2014, 89, 19–28. [Google Scholar] [CrossRef]
- Wang, F.; Tong, Q. SIRT2 suppresses adipocyte differentiation by deacetylating FOXO1 and enhancing FOXO1’s repressive interaction with PPARγ. Mol. Biol. Cell 2009, 20, 801–808. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, J.; Tong, Q.; Lin, L. SIRT3 acts as a positive autophagy regulator to promote lipid mobilization in adipocytes via activating AMPK. Int. J. Mol. Sci. 2020, 21, 372. [Google Scholar] [CrossRef] [PubMed]
- Laurent, G.; German, N.J.; Saha, A.K.; de Boer, V.C.; Davies, M.; Koves, T.R.; Dephoure, N.; Fischer, F.; Boanca, G.; Vaitheesvaran, B.; et al. SIRT4 coordinates the balance between lipid synthesis and catabolism by repressing malonyl CoA decarboxylase. Mol. Cell 2013, 50, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Meyer, J.G.; Cai, W.; Softic, S.; Li, M.E.; Verdin, E.; Newgard, C.; Schilling, B.; Kahn, C.R. Regulation of UCP1 and mitochondrial metabolism in brown adipose tissue by reversible succinylation. Mol. Cell 2019, 74, 844–857.e7. [Google Scholar] [CrossRef] [PubMed]
- Bober, E.; Fang, J.; Smolka, C.; Ianni, A.; Vakhrusheva, O.; Krüger, M.; Braun, T. Sirt7 promotes adipogenesis by binding to and inhibiting Sirt1. BMC Proc. 2012, 6, P57. [Google Scholar] [CrossRef]
- Xu, F.; Zheng, X.; Lin, B.; Liang, H.; Cai, M.; Cao, H.; Ye, J.; Weng, J. Diet-induced obesity and insulin resistance are associated with brown fat degeneration in SIRT 1-deficient mice. Obesity 2016, 24, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, T.; Hu, J.; Ma, R.; He, B.; Wang, M.; Wang, Y. Adiponectin/SIRT1 axis induces white adipose browning after vertical sleeve gastrectomy of obese rats with type 2 diabetes. Obes. Surg. 2020, 30, 1392–1403. [Google Scholar] [CrossRef]
- Gao, P.; Jiang, Y.; Wu, H.; Sun, F.; Li, Y.; He, H.; Wang, B.; Lu, Z.; Hu, Y.; Wei, X.; et al. Inhibition of mitochondrial calcium overload by SIRT3 prevents obesity-or age-related whitening of brown adipose tissue. Diabetes 2020, 69, 165–180. [Google Scholar] [CrossRef]
- Shuai, L.; Zhang, L.-N.; Li, B.-H.; Tang, C.-L.; Wu, L.-Y.; Li, J.; Li, J.-Y. SIRT5 regulates brown adipocyte differentiation and browning of subcutaneous white adipose tissue. Diabetes 2019, 68, 1449–1461. [Google Scholar] [CrossRef]
- Wu, D.; Bang, I.H.; Park, B.-H.; Bae, E.J. Loss of Sirt6 in adipocytes impairs the ability of adipose tissue to adapt to intermittent fasting. Exp. Mol. Med. 2021, 53, 1298–1306. [Google Scholar] [CrossRef]
- Qiang, L.; Wang, L.; Kon, N.; Zhao, W.; Lee, S.; Zhang, Y.; Rosenbaum, M.; Zhao, Y.; Gu, W.; Farmer, S.R.; et al. Brown remodeling of white adipose tissue by SirT1-dependent deacetylation of Pparγ. Cell 2012, 150, 620–632. [Google Scholar] [CrossRef]
- Cantó, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Bouviere, J.; Fortunato, R.S.; Dupuy, C.; Werneck-de-Castro, J.P.; Carvalho, D.P.; Louzada, R.A. Exercise-stimulated ROS sensitive signaling pathways in skeletal muscle. Antioxidants 2021, 10, 537. [Google Scholar] [CrossRef] [PubMed]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Zu, Y.-X.; Lu, H.-Y.; Liu, W.-W.; Jiang, X.-W.; Huang, Y.; Li, X.; Zhao, Q.-C.; Xu, Z.-H. Jiang Gui Fang activated interscapular brown adipose tissue and induced epididymal white adipose tissue browning through the PPARγ/SIRT1-PGC1α pathway. J. Ethnopharmacol. 2020, 248, 112271. [Google Scholar] [CrossRef]
- Zhou, J.; Poudel, A.; Chandramani-Shivalingappa, P.; Xu, B.; Welchko, R.; Li, L. Liraglutide induces beige fat development and promotes mitochondrial function in diet induced obesity mice partially through AMPK-SIRT-1-PGC1-α cell signaling pathway. Endocrine 2019, 64, 271–283. [Google Scholar] [CrossRef]
- Xu, Y.; Yu, T.; Ma, G.; Zheng, L.; Jiang, X.; Yang, F.; Wang, Z.; Li, N.; He, Z.; Song, X.; et al. Berberine modulates deacetylation of PPARγ to promote adipose tissue remodeling and thermogenesis via AMPK/SIRT1 pathway. Int. J. Biol. Sci. 2021, 17, 3173–3187. [Google Scholar] [CrossRef]
- Yao, E.; Yang, X.; Huang, X.; Mi, Y.; Wu, X.; Fang, M.; Huang, J.; Qiu, Y.; Hong, X.; Peng, L.; et al. Phytochemical wedelolactone reverses obesity by prompting adipose browning through SIRT1/AMPK/PPARα pathway via targeting nicotinamide N-methyltransferase. Phytomedicine 2022, 94, 153843. [Google Scholar] [CrossRef]
- Baskaran, P.; Krishnan, V.; Ren, J.; Thyagarajan, B. Capsaicin induces browning of white adipose tissue and counters obesity by activating TRPV1 channel-dependent mechanisms. Br. J. Pharmacol. 2016, 173, 2369–2389. [Google Scholar] [CrossRef]
- Chen, C.-C.; Kuo, C.-H.; Leu, Y.-L.; Wang, S.-H. Corylin reduces obesity and insulin resistance and promotes adipose tissue browning through SIRT-1 and β3-AR activation. Pharmacol. Res. 2021, 164, 105291. [Google Scholar] [CrossRef]
- Kong, S.; Cai, B.; Nie, Q. PGC-1α affects skeletal muscle and adipose tissue development by regulating mitochondrial biogenesis. Mol. Genet. Genom. 2022, 297, 621–633. [Google Scholar] [CrossRef]
- Milton-Laskibar, I.; Aguirre, L.; Etxeberria, U.; Milagro, F.I.; Martínez, J.A.; Portillo, M.P. Do the effects of resveratrol on thermogenic and oxidative capacities in IBAT and skeletal muscle depend on feeding conditions? Nutrients 2018, 10, 1446. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Curry, B.J.; Brown, P.L.; Zemel, M.B. Leucine modulates mitochondrial biogenesis and SIRT1-AMPK signaling in C2C12 myotubes. J. Nutr. Metab. 2014, 2014, 239750. [Google Scholar] [CrossRef] [PubMed]
- Perrini, S.; Porro, S.; Nigro, P.; Cignarelli, A.; Caccioppoli, C.; Genchi, V.A.; Martines, G.; De Fazio, M.; Capuano, P.; Natalicchio, A.; et al. Reduced SIRT1 and SIRT2 expression promotes adipogenesis of human visceral adipose stem cells and associates with accumulation of visceral fat in human obesity. Int. J. Obes. 2020, 44, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Sebaa, R.; Johnson, J.; Pileggi, C.; Norgren, M.; Xuan, J.; Sai, Y.; Tong, Q.; Krystkowiak, I.; Bondy-Chorney, E.; Davey, N.E.; et al. SIRT3 controls brown fat thermogenesis by deacetylation regulation of pathways upstream of UCP1. Mol. Metab. 2019, 25, 35–49. [Google Scholar] [CrossRef]
- Wei, T.; Huang, G.; Liu, P.; Gao, J.; Huang, C.; Sun, M.; Shen, W. Sirtuin 3-mediated pyruvate dehydrogenase activity determines brown adipocytes phenotype under high-salt conditions. Cell Death Dis. 2019, 10, 614. [Google Scholar] [CrossRef]
- Giralt, A.; Hondares, E.; Villena, J.A.; Ribas, F.; Díaz-Delfín, J.; Giralt, M.; Iglesias, R.; Villarroya, F. Peroxisome proliferator-activated receptor-γ coactivator-1α controls transcription of the Sirt3 gene, an essential component of the thermogenic brown adipocyte phenotype. J. Biol. Chem. 2011, 286, 16958–16966. [Google Scholar] [CrossRef]
- Cheng, C.-Y.; Yang, A.-J.; Ekambaranellore, P.; Huang, K.-C.; Lin, W.-W. Anti-obesity action of INDUS810, a natural compound from Trigonella foenum-graecum: AMPK-dependent lipolysis effect in adipocytes. Obes. Res. Clin. Pract. 2018, 12, 562–569. [Google Scholar] [CrossRef]
- Lone, J.; Choi, J.H.; Kim, S.W.; Yun, J.W. Curcumin induces brown fat-like phenotype in 3T3-L1 and primary white adipocytes. J. Nutr. Biochem. 2016, 27, 193–202. [Google Scholar] [CrossRef]
- Park, W.Y.; Park, J.; Ahn, K.S.; Kwak, H.J.; Um, J.Y. Ellagic acid induces beige remodeling of white adipose tissue by controlling mitochondrial dynamics and SIRT3. FASEB J. 2021, 35, e21548. [Google Scholar] [CrossRef]
- Molinari, F.; Feraco, A.; Mirabilii, S.; Saladini, S.; Sansone, L.; Vernucci, E.; Tomaselli, G.; Marzolla, V.; Rotili, D.; Russo, M.A.; et al. SIRT5 inhibition induces brown fat-like phenotype in 3T3-L1 preadipocytes. Cells 2021, 10, 1126. [Google Scholar] [CrossRef]
- Zhang, C.; He, X.; Sheng, Y.; Xu, J.; Yang, C.; Zheng, S.; Liu, J.; Li, H.; Ge, J.; Yang, M.; et al. Allicin regulates energy homeostasis through brown adipose tissue. IScience 2020, 23, 101113. [Google Scholar] [CrossRef] [PubMed]
- Bae, E.J.; Park, B.-H. Multiple Roles of Sirtuin 6 in Adipose Tissue Inflammation. Korean Diabetes J. 2023, 47, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Cui, X.; Chen, Q.; Yang, X.; Fang, F.; Zhang, J.; Liu, G.; Jin, W.; Chang, Y. Cold-inducible SIRT6 regulates thermogenesis of brown and beige fat. Cell Rep. 2017, 20, 641–654. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Sato, Y.; Sobuz, S.U.; Mizumoto, T.; Tsuyama, T.; Karim, M.F.; Miyata, K.; Tasaki, M.; Yamazaki, M.; Kariba, Y.; et al. SIRT7 suppresses energy expenditure and thermogenesis by regulating brown adipose tissue functions in mice. Nat. Commun. 2022, 13, 7439. [Google Scholar] [CrossRef]
- Chang, C.-C.; Lin, K.-Y.; Peng, K.-Y.; Day, Y.-J.; Hung, L.-M. Resveratrol exerts anti-obesity effects in high-fat diet obese mice and displays differential dosage effects on cytotoxicity, differentiation, and lipolysis in 3T3-L1 cells. Endocr. J. 2016, 63, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Bouzar, C.; Cottet-Rousselle, C.; Zagotta, I.; Lamarche, F.; Wabitsch, M.; Tokarska-Schlattner, M.; Fischer-Posovszky, P.; Schlattner, U.; Rousseau, D. Resveratrol inhibits lipogenesis of 3T3-L1 and SGBS cells by inhibition of insulin signaling and mitochondrial mass increase. Biochim. Biophys. Acta (BBA)-Bioenerg. 2016, 1857, 643–652. [Google Scholar] [CrossRef]
- Deng, X.; Chen, B.; Luo, Q.; Zao, X.; Liu, H.; Li, Y. Hulless barley polyphenol extract inhibits adipogenesis in 3T3-L1 cells and obesity related-enzymes. Front. Nutr. 2022, 9, 933068. [Google Scholar] [CrossRef]
- Hong, S.Y.; Ha, A.W.; Kim, W. Effects of quercetin on cell differentiation and adipogenesis in 3T3-L1 adipocytes. Nutr. Res. Pract. 2021, 15, 444–455. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, L.; Dong, L.; Hu, X.; Feng, F.; Chen, F. 6-gingerol, a functional polyphenol of ginger, promotes browning through an AMPK-dependent pathway in 3T3-L1 adipocytes. J. Agric. Food Chem. 2019, 67, 14056–14065. [Google Scholar] [CrossRef]
- Aranaz, P.; Navarro-Herrera, D.; Zabala, M.; Miguéliz, I.; Romo-Hualde, A.; López-Yoldi, M.; Martínez, J.A.; Vizmanos, J.L.; Milagro, F.I.; González-Navarro, C.J. Phenolic compounds inhibit 3T3-L1 adipogenesis depending on the stage of differentiation and their binding affinity to PPARγ. Molecules 2019, 24, 1045. [Google Scholar] [CrossRef]
- Rayalam, S.; Yang, J.Y.; Ambati, S.; Della-Fera, M.A.; Baile, C.A. Resveratrol induces apoptosis and inhibits adipogenesis in 3T3-L1 adipocytes. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2008, 22, 1367–1371. [Google Scholar] [CrossRef] [PubMed]
- Dayarathne, L.A.; Ranaweera, S.S.; Natraj, P.; Rajan, P.; Lee, Y.J.; Han, C.-H. Restoration of the adipogenic gene expression by naringenin and naringin in 3T3-L1 adipocytes. J. Vet. Sci. 2021, 22, e55. [Google Scholar] [CrossRef] [PubMed]
- Pacifici, F.; Malatesta, G.; Mammi, C.; Pastore, D.; Marzolla, V.; Ricordi, C.; Chiereghin, F.; Infante, M.; Donadel, G.; Curcio, F.; et al. A Novel Mix of Polyphenols and Micronutrients Reduces Adipogenesis and Promotes White Adipose Tissue Browning via UCP1 Expression and AMPK Activation. Cells 2023, 12, 714. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liao, W.; Yin, X.; Zheng, X.; Li, Q.; Zhang, H.; Zheng, L.; Feng, X. Resveratrol-induced brown fat-like phenotype in 3T3-L1 adipocytes partly via mTOR pathway. Food Nutr. Res. 2020, 64, 3565. [Google Scholar] [CrossRef] [PubMed]
- Rebello, C.J.; Greenway, F.L.; Lau, F.H.; Lin, Y.; Stephens, J.M.; Johnson, W.D.; Coulter, A.A. Naringenin promotes thermogenic gene expression in human white adipose tissue. Obesity 2019, 27, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Song, N.-J.; Lee, A.R.; Lee, D.H.; Seo, M.-J.; Kim, S.; Chang, S.-H.; Yang, D.K.; Hwang, Y.-J.; Hwang, K.-A. Oxyresveratrol increases energy expenditure through Foxo3a-mediated Ucp1 induction in high-fat-diet-induced obese mice. Int. J. Mol. Sci. 2018, 20, 26. [Google Scholar] [CrossRef]
- Choi, H.; Kim, C.-S.; Yu, R. Quercetin upregulates uncoupling protein 1 in white/brown adipose tissues through sympathetic stimulation. J. Obes. Metab. Syndr. 2018, 27, 102–109. [Google Scholar] [CrossRef]
- Wang, S.; Liang, X.; Yang, Q.; Fu, X.; Rogers, C.J.; Zhu, M.; Rodgers, B.; Jiang, Q.; Dodson, M.V.; Du, M. Resveratrol induces brown-like adipocyte formation in white fat through activation of AMP-activated protein kinase (AMPK) α1. Int. J. Obes. 2015, 39, 967–976. [Google Scholar] [CrossRef]
- Testai, L.; De Leo, M.; Flori, L.; Polini, B.; Braca, A.; Nieri, P.; Pistelli, L.; Calderone, V. Contribution of irisin pathway in protective effects of mandarin juice (Citrus reticulata Blanco) on metabolic syndrome in rats fed with high fat diet. Phytother. Res. 2021, 35, 4324–4333. [Google Scholar] [CrossRef]
- De Leo, M.; Piragine, E.; Pirone, A.; Braca, A.; Pistelli, L.; Calderone, V.; Miragliotta, V.; Testai, L. Protective effects of bergamot (Citrus bergamia risso & poiteau) juice in rats fed with high-fat diet. Planta Med. 2020, 86, 180–189. [Google Scholar]
- Qiao, Y.; Sun, J.; Xia, S.; Tang, X.; Shi, Y.; Le, G. Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct. 2014, 5, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-J.; Jung, U.J.; Choi, M.-S. Differential effects of low-dose resveratrol on adiposity and hepatic steatosis in diet-induced obese mice. Br. J. Nutr. 2012, 108, 2166–2175. [Google Scholar] [CrossRef] [PubMed]
- Fukuchi, Y.; Hiramitsu, M.; Okada, M.; Hayashi, S.; Nabeno, Y.; Osawa, T.; Naito, M. Lemon polyphenols suppress diet-induced obesity by up-regulation of mRNA levels of the enzymes involved in β-oxidation in mouse white adipose tissue. J. Clin. Biochem. Nutr. 2008, 43, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Hui, S.; Liu, Y.; Huang, L.; Zheng, L.; Zhou, M.; Lang, H.; Wang, X.; Yi, L.; Mi, M. Resveratrol enhances brown adipose tissue activity and white adipose tissue browning in part by regulating bile acid metabolism via gut microbiota remodeling. Int. J. Obes. 2020, 44, 1678–1690. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, M.; Wang, S. Browning Subcutaneous White Adipose Tissue Using Transdermal Delivery of ASC-Targeting Peptide and Resveratrol Conjugate. Curr. Dev. Nutr. 2022, 6, 1042. [Google Scholar] [CrossRef]
- Crespillo, A.; Alonso, M.; Vida, M.; Pavón, F.; Serrano, A.; Rivera, P.; Romero-Zerbo, Y.; Fernández-Llebrez, P.; Martínez, A.; Pérez-Valero, V. Reduction of body weight, liver steatosis and expression of stearoyl-CoA desaturase 1 by the isoflavone daidzein in diet-induced obesity. Br. J. Pharmacol. 2011, 164, 1899–1915. [Google Scholar] [CrossRef] [PubMed]
- Oi-Kano, Y.; Kawada, T.; Watanabe, T.; Koyama, F.; Watanabe, K.; Senbongi, R.; Iwai, K. Oleuropein, a phenolic compound in extra virgin olive oil, increases uncoupling protein 1 content in brown adipose tissue and enhances noradrenaline and adrenaline secretions in rats. J. Nutr. Sci. Vitaminol. 2008, 54, 363–370. [Google Scholar] [CrossRef]
- Mezhibovsky, E.; Knowles, K.A.; He, Q.; Sui, K.; Tveter, K.M.; Duran, R.M.; Roopchand, D.E. Grape polyphenols attenuate diet-induced obesity and hepatic steatosis in mice in association with reduced butyrate and increased markers of intestinal carbohydrate oxidation. Front. Nutr. 2021, 8, 675267. [Google Scholar] [CrossRef]
- Kudo, N.; Arai, Y.; Suhara, Y.; Ishii, T.; Nakayama, T.; Osakabe, N. A single oral administration of theaflavins increases energy expenditure and the expression of metabolic genes. PLoS ONE 2015, 10, e0137809. [Google Scholar] [CrossRef]
- Zhou, J.; Mao, L.; Xu, P.; Wang, Y. Effects of (−)-epigallocatechin gallate (EGCG) on energy expenditure and microglia-mediated hypothalamic inflammation in mice fed a high-fat diet. Nutrients 2018, 10, 1681. [Google Scholar] [CrossRef]
- Osakabe, N.; Hoshi, J.; Kudo, N.; Shibata, M. The flavan-3-ol fraction of cocoa powder suppressed changes associated with early-stage metabolic syndrome in high-fat diet-fed rats. Life Sci. 2014, 114, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Osborn, L.J.; Schultz, K.; Massey, W.; DeLucia, B.; Choucair, I.; Varadharajan, V.; Banerjee, R.; Fung, K.; Horak III, A.J.; Orabi, D. A gut microbial metabolite of dietary polyphenols reverses obesity-driven hepatic steatosis. Proc. Natl. Acad. Sci. USA 2022, 119, e2202934119. [Google Scholar] [CrossRef] [PubMed]
- Tamura, Y.; Tomiya, S.; Takegaki, J.; Kouzaki, K.; Tsutaki, A.; Nakazato, K. Apple polyphenols induce browning of white adipose tissue. J. Nutr. Biochem. 2020, 77, 108299. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Okabe, M.; Natsume, M.; Ashida, H. Prevention mechanisms of glucose intolerance and obesity by cacao liquor procyanidin extract in high-fat diet-fed C57BL/6 mice. Arch. Biochem. Biophys. 2012, 527, 95–104. [Google Scholar] [CrossRef]
- Abedi-Taleb, E.; Vahabi, Z.; Sekhavati-Moghadam, E.; Khedmat, L.; Jazayeri, S.; Saboor-Yaraghi, A.A. Upregulation of FNDC5 gene expression in C2C12 cells after single and combined treatments of resveratrol and ATRA. Lipids Health Dis. 2019, 18, 181. [Google Scholar] [CrossRef] [PubMed]
- Mele, L.; Bidault, G.; Mena, P.; Crozier, A.; Brighenti, F.; Vidal-Puig, A.; Del Rio, D. Dietary (Poly) phenols, brown adipose tissue activation, and energy expenditure: A narrative review. Adv. Nutr. 2017, 8, 694–704. [Google Scholar] [CrossRef]
- Ciccone, L.; Piragine, E.; Brogi, S.; Camodeca, C.; Fucci, R.; Calderone, V.; Nencetti, S.; Martelli, A.; Orlandini, E. Resveratrol-like Compounds as SIRT1 Activators. Int. J. Mol. Sci. 2022, 23, 15105. [Google Scholar] [CrossRef]
- Sarubbo, F.; Moranta, D.; Asensio, V.J.; Miralles, A.; Esteban, S. Effects of resveratrol and other polyphenols on the most common brain age-related diseases. Curr. Med. Chem. 2017, 24, 4245–4266. [Google Scholar] [CrossRef]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Alberdi, G.; Rodríguez, V.M.; Miranda, J.; Macarulla, M.T.; Churruca, I.; Portillo, M.P. Thermogenesis is involved in the body-fat lowering effects of resveratrol in rats. Food Chem. 2013, 141, 1530–1535. [Google Scholar] [CrossRef]
- Andrade, J.M.O.; Frade, A.C.M.; Guimaraes, J.B.; Freitas, K.M.; Lopes, M.T.P.; Guimarães, A.L.S.; De Paula, A.M.B.; Coimbra, C.C.; Santos, S.H.S. Resveratrol increases brown adipose tissue thermogenesis markers by increasing SIRT1 and energy expenditure and decreasing fat accumulation in adipose tissue of mice fed a standard diet. Eur. J. Nutr. 2014, 53, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Ai, H.; Yuan, S.; Cao, H.; Liang, H.; Weng, J.; Xu, F. Effect of SIRT1 deficiency on function of brown adipose tissue in obese mice. Zhonghua Yi Xue Za Zhi 2016, 96, 1859–1862. [Google Scholar] [PubMed]
- Jimenez-Gomez, Y.; Mattison, J.A.; Pearson, K.J.; Martin-Montalvo, A.; Palacios, H.H.; Sossong, A.M.; Ward, T.M.; Younts, C.M.; Lewis, K.; Allard, J.S.; et al. Resveratrol improves adipose insulin signaling and reduces the inflammatory response in adipose tissue of rhesus monkeys on high-fat, high-sugar diet. Cell Metab. 2013, 18, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Aziz, S.A.; Wakeling, L.A.; Miwa, S.; Alberdi, G.; Hesketh, J.E.; Ford, D. Metabolic programming of a beige adipocyte phenotype by genistein. Mol. Nutr. Food Res. 2017, 61, 1600574. [Google Scholar] [CrossRef]
- Gutiérrez-Salmeán, G.; Ortiz-Vilchis, P.; Vacaseydel, C.M.; Garduño-Siciliano, L.; Chamorro-Cevallos, G.; Meaney, E.; Villafaña, S.; Villarreal, F.; Ceballos, G.; Ramírez-Sánchez, I. Effects of (−)-epicatechin on a diet-induced rat model of cardiometabolic risk factors. Eur. J. Pharmacol. 2014, 728, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhang, X.; Zhang, L.; Bian, H.-X.; Xu, N.; Bao, B.; Liu, J. Quercetin reduces obesity-associated ATM infiltration and inflammation in mice: A mechanism including AMPKα1/SIRT1 [S]. J. Lipid Res. 2014, 55, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Cen, F.; Tian, F.; Li, M.-J.; Zhang, Q.; Shen, H.Y.; Shen, X.-C.; Zhou, M.M.; Du, J. Combination treatment with quercetin and resveratrol attenuates high fat diet-induced obesity and associated inflammation in rats via the AMPKα1/SIRT1 signaling pathway. Exp. Ther. Med. 2017, 14, 5942–5948. [Google Scholar] [CrossRef]
- Brown Fat Activation and Browning Efficiency Augmented by Chronic Cold and Nutraceuticals for Brown Adipose Tissue-mediated Effect Against Metabolic Syndrome (BEACON BEAMS Study). ClinicalTrials.gov Identifier: NCT04595006. 2021. Available online: https://ClinicalTrials.gov/show/NCT04595006 (accessed on 13 April 2023).
- Goh, K.P.; Lee, H.Y.; Lau, D.P.; Supaat, W.; Chan, Y.H.; Koh, A.F.Y. Effects of resveratrol in patients with type 2 diabetes mellitus on skeletal muscle SIRT1 expression and energy expenditure. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 2–13. [Google Scholar] [CrossRef]
- Assessing the Value of Red Grapes Polyphenol Supplementation on Metabolic Parameters in Obese Insulinoresistant Subjects (POLYGIR Study). ClinicalTrials.gov Identifier: NCT02633150. 2015. Available online: https://ClinicalTrials.gov/show/NCT02633150 (accessed on 13 April 2023).
- Hughes, L.A.; Arts, I.C.; Ambergen, T.; Brants, H.A.; Dagnelie, P.C.; Goldbohm, R.A.; van den Brandt, P.A.; Weijenberg, M.P. Higher dietary flavone, flavonol, and catechin intakes are associated with less of an increase in BMI over time in women: A longitudinal analysis from the Netherlands Cohort Study. Am. J. Clin. Nutr. 2008, 88, 1341–1352. [Google Scholar]
- Aali, Y.; Ebrahimi, S.; Shiraseb, F.; Mirzaei, K. The association between dietary polyphenol intake and cardiometabolic factors in overweight and obese women: A cross-sectional study. BMC Endocr. Disord. 2022, 22, 120. [Google Scholar] [CrossRef]
- Suliburska, J.; Bogdanski, P.; Szulinska, M.; Stepien, M.; Pupek-Musialik, D.; Jablecka, A. Effects of green tea supplementation on elements, total antioxidants, lipids, and glucose values in the serum of obese patients. Biol. Trace Elem. Res. 2012, 149, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-J.; Liu, C.-Y.; Chiu, J.-P.; Hsu, C.-H. Therapeutic effect of high-dose green tea extract on weight reduction: A randomized, double-blind, placebo-controlled clinical trial. Clin. Nutr. 2016, 35, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Dow, C.A.; Going, S.B.; Chow, H.-H.S.; Patil, B.S.; Thomson, C.A. The effects of daily consumption of grapefruit on body weight, lipids, and blood pressure in healthy, overweight adults. Metabolism 2012, 61, 1026–1035. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Cha, Y.-J.; Lee, K.-H.; Yim, J.-E. Onion peel extract reduces the percentage of body fat in overweight and obese subjects: A 12-week, randomized, double-blind, placebo-controlled study. Nutr. Res. Pract. 2016, 10, 175–181. [Google Scholar] [CrossRef]
- Saraf-Bank, S.; Ahmadi, A.; Paknahad, Z.; Maracy, M.; Nourian, M. Effects of curcumin on cardiovascular risk factors in obese and overweight adolescent girls: A randomized clinical trial. Sao Paulo Med. J. 2019, 137, 414–422. [Google Scholar] [CrossRef]
- Tabrizi, R.; Tamtaji, O.R.; Lankarani, K.B.; Akbari, M.; Dadgostar, E.; Dabbaghmanesh, M.H.; Kolahdooz, F.; Shamshirian, A.; Momen-Heravi, M.; Asemi, Z. The effects of resveratrol intake on weight loss: A systematic review and meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 375–390. [Google Scholar] [CrossRef]
- Delpino, F.M.; Figueiredo, L.M.; Caputo, E.L.; Mintem, G.C.; Gigante, D.P. What is the effect of resveratrol on obesity? A systematic review and meta-analysis. Clin. Nutr. ESPEN 2021, 41, 59–67. [Google Scholar] [CrossRef]
- Mousavi, S.; Milajerdi, A.; Sheikhi, A.; Kord-Varkaneh, H.; Feinle-Bisset, C.; Larijani, B.; Esmaillzadeh, A. Resveratrol supplementation significantly influences obesity measures: A systematic review and dose–response meta-analysis of randomized controlled trials. Obes. Rev. 2019, 20, 487–498. [Google Scholar] [CrossRef]
- Galet, C.; Gollapudi, K.; Stepanian, S.; Byrd, J.B.; Henning, S.M.; Grogan, T.; Elashoff, D.; Heber, D.; Said, J.; Cohen, P. Effect of a low-fat fish oil diet on proinflammatory eicosanoids and cell-cycle progression score in men undergoing radical prostatectomylow-fat fish oil diet, serum eicosanoids, and prostate cancer. Cancer Prev. Res. 2014, 7, 97–104. [Google Scholar] [CrossRef]
- Johnson, J.J.; Nihal, M.; Siddiqui, I.A.; Scarlett, C.O.; Bailey, H.H.; Mukhtar, H.; Ahmad, N. Enhancing the bioavailability of resveratrol by combining it with piperine. Mol. Nutr. Food Res. 2011, 55, 1169–1176. [Google Scholar] [CrossRef]
- Tolve, R.; Galgano, F.; Condelli, N.; Cela, N.; Lucini, L.; Caruso, M.C. Optimization model of phenolics encapsulation conditions for biofortification in fatty acids of animal food products. Foods 2021, 10, 881. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Dong, Y.; Wang, F.; Zhang, Y. Nanoformulations to enhance the bioavailability and physiological functions of polyphenols. Molecules 2020, 25, 4613. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| First Author, Year, Reference | Cell Line | Compound, Concentration | Time | Potential Mechanism | Main Effects |
|---|---|---|---|---|---|
| Aranaz, 2019, [122] | 3T3-L1 | Phenolic compounds 1, 10–100 µM | 8 days *,** | ↓ PPAR-γ a at the initial stage of differentiation | ↓ lipid accumulation depending on the stage of differentiation |
| Chang, 2016, [117,129] | 3T3-L1 | Resveratrol, 0.03–10 µM | 8 days *; 24 h ** | ↓ PPAR-γ b | ↓ preadipocyte differentiation ↑ mature adipocyte lipolysis |
| Choi, 2018, [129] | 3T3-L1 | Quercetin, 10 µM | 6 days * | ↑ UCP1 b | - |
| Choi, 2018, [128] | 3T3-L1, C3H10T1/2 | Oxyresveratrol, 100 µM | 6 days *; 24 h ** | ↓ PPAR-γ a ↑ UCP1 a | ↓ lipid accumulation and adipocyte differentiation ↑ mitochondrial mass |
| Dayarathne, 2021, [124] | 3T3-L1 | Naringenin and naringin, 20–100 µM | 8 days * | ↑ p-AMPK b, p-ACC b | ↓ lipid and triglyceride accumulation |
| Deng, 2022, [119] | 3T3-L1 | Polyphenol extract, 5–25 μg FAE/ml | 48 h * | ↓ PPAR-γ b, FAS a,b | ↓ lipid and triglyceride accumulation ↓ oxidative stress |
| Hong, 2021, [120] | 3T3-L1 | Quercetin, 5–20 µM | 24 h ** | ↓ PPAR-γ a | ↓ fat accumulation |
| Li, 2016, [118] | 3T3-L1, SGBS | Resveratrol, 50–100 µM | 4 **–8 * days (3T3-L1), 6 **–12 * days (SGBS) | ↓ AMPK b, ACC b | ↓ lipid accumulation |
| Liu, 2020, [126] | 3T3-L1 | Resveratrol, 10–40 µM | 6–8 days * | ↑ PGC-1α b, PPAR-γ b, UCP1b | ↓ lipid accumulation |
| Pacifici, 2023, [125] | 3T3-L1 | Mix of polyphenols 2, N/A | 10 days * | ↑ CIDEA a.b, p-AMPK b, UCP1 a,b | ↓ lipid accumulation ↓ inflammation ↑ fatty acid oxidation |
| Rayalam, 2008, [123] | 3T3-L1 | Resveratrol, 12.5–50 µM | 6 days * | ↓ PPAR-γ a, FAS a ↑ UCP1 a | ↓ lipid accumulation |
| Rebello, 2019, [127] | hADSC and pWAT from overweight patients | Naringenin, 8 µM | 7–14 days * | ↑ PGC-1α a, PGC-1β a, p-AMPK b, UCP1 a,b | - |
| Wang, 2015, [130] | Differentiated iWAT SVCs | Resveratrol, 10–40 µM | 7 days * | ↑ PRDM16 a,b, PGC-1α a, CIDEA a, p-AMPK b, UCP1 a,b | ↓ lipid accumulation ↑ O2 consumption |
| Wang, 2019, [121] | 3T3-L1 | 6-Gingerol, 20 μg/ml | Until differentiation * | ↑ PGC-1α a,b, PRDM-16 a,b, p-AMPK b, UCP1 a,b ↓ PPAR-γ a | ↓ lipid content of mature adipocytes ↑ mitochondrial respiration and energy metabolism |
| First Author, Year, Reference | Animal Model | Number (Control/ Treated) | Compound, Dosage, Route of Administration | Weeks of Treatment | Potential Mechanism | Main Effects |
|---|---|---|---|---|---|---|
| Abbasi, 2022, [137] | C57BL/6J mice # | 5/5 | Peptide-resveratrol conjugate, N/A, transdermal | 4 | ↑ UCP1 a in WAT | ↓ body weight |
| Chang, 2016, [117] | C57BL/6C mice # | 10–11/ 10–11 | Resveratrol, 1–30 mg/kg, orally | 10 | - | ↓ diet-induced body weight ↓ total WAT weight 1 |
| Cho, 2012, [134] | C57BL/6J mice # | 10/10 | Resveratrol, 0.005–0.02% w/w, orally | 10 | ↓ FAS activity in eWAT ↑ β-oxidation in eWAT | ↓ body weight ↓ total WAT weight 2 ↓ epididymal adipocyte size |
| Choi, 2018, [129] | C57BL/6J mice # | 6/6 | Quercetin, 0.05% w/w, orally | 9 | ↑ PRDM16 a, CIDEA a, PPAR-γ b, PGC-1α b, UCP1 a,b in WAT ↑ UCP1 in BAT b | ↓ WAT weight and adipocyte size |
| Choi, 2018, [128] | C57BL/6N mice # | 6/7 | Oxyresveratrol, 7.5–15 mg/kg, i.p. | 8 | ↑ PRDM16 a,b, PGC-1α a,b, UCP-1 a,b in iWAT | ↓ body weight ↓ eWAT weight ↑ energy expenditure |
| Crespillo, 2011, [138] | Wistar rats # | 16/16 | Daidzein, 50 mg/kg, i.p. | 2 | ↑ UCP1 b in BAT | ↓ body weight ↓ fat depots in the liver |
| De Leo, 2020, [132] | Wistar rats # | 6/6 | Bergamot juice, 12% v/v, orally | 3 | - | ↓ body weight ↓ fat depots in the liver |
| Fukuchi, 2008, [135] | C57BL6/J mice # | 6/6 | Lemon polyphenols, 0.5% w/w, orally | 12 | ↑ FAS a and ACO a in eWAT | ↓ body weight ↓ insulin resistance ↓ total WAT weight 3 |
| Hui, 2020, [136] | db/db mice | 8/8 | Resveratrol, 0.4% w/w, orally | 6 | ↑ PRDM16 a, CIDEA a, UCP1 a,b in iWAT/BAT ↑ PPAR-γ a in iWAT | ↓ eWAT and iWAT ↓ sizes of lipid droplets in iWAT and BAT |
| Kudo, 2015, [141] | ICR mice | 8/8 | Theaflavins, 10 mg/kg, orally | Acute treatment (2–20 h) | ↑ UCP1 a in BAT ↑ p-AMPK b, UCP3 a in SKM ↑ PGC-1α a in BAT/SKM | ↑ energy expenditure |
| Mezhibovsy, 2021, [140] | C57BL/6J mice # | 6–8/6–8 | Grape polyphenols, 1.0% w/w, orally | 23 | Inverse relationship between body weight and UCP1 a in BAT | ↓ body weight ↓ FAT mass ↑ energy expenditure ↓ fat depots in the liver |
| Oi-Kano, 2008, [139] | Sprague-Dawley rats # | 6–7/6–7 | Oleuropein, 1–4 mg/kg, orally | 4 | ↑ UCP1 b in iBAT | ↓ body weight ↓ eWAT and pWAT |
| Osakabe, 2014, [143] | Wistar rats # | 7/8 | Flavan-3-ols from cocoa powder, 0.2% w/w, orally | 4 | ↑ UCP1 b in BAT ↑ UCP3 b in the SKM | ↓ eWAT |
| Osborn, 2022_I, [144] | C57BL/6 mice # | 9–10/9–10 | Flavonoid composites, 1% w/w, orally | 16 | - | ↓ body weight and fat mass ↑ lean mass |
| Osborn, 2022_II, [144] | C57BL/6 mice # | 9–10/9–10 | 4-HPAA 350 µg/day, transdermal | 6 | ↑ PGC-1α a and UCP1 a in BAT | ↓ fat depots in the liver |
| Qiao, 2014, [133] | Kunming mice # | 8/8 | Resveratrol, 200 mg/kg, orally | 12 | ↓ PPAR-γ a, ACC a, FAS a in eWAT | ↓ body weight ↓ visceral and sWAT weight |
| Tamura, 2020, [145] | C57BL/6J mice # | 7/7 | Apple polyphenols, 5.0% w/w, orally | 4 | ↑ OXPHOS b in eWAT ↑ OXPHOS b, PGC-1α b, FGF21 a,b, p-FRS2α b, p-Erk1/2 b, p-MAPK b, UCP1 b in iWAT | ↓ body weight ↓ eWAT, iWAT and BAT weight ↑ thermogenic adaptations |
| Testai, 2021, [131] | Wistar rats # | 6/6 | Bergamot juice, 24% v/v, orally | 3 | ↑ PGC-1α a and UCP1 a in WAT ↑ plasma levels of irisin | ↓ body weight ↓ WAT weight |
| Wang, 2015, [130] | CD1 mice # | 6/6 | Resveratrol, 0.1% w/w, orally | 4 | ↑ PRDM16 b, p-AMPK b and UCP1 b in iWAT | ↓ body weight ↓ iWAT weight and adipocyte size ↑ browning in iWAT ↑ O2 consumption in iWAT |
| Yamashita, 2012, [146] | C57BL6/J mice # | 6/6 | Cacao liquor procyanidins, 0.5–2.0% w/w, orally | 13 | ↑ p-AMPK b, PGC-1α a and UCP2 a,b in WAT ↑ UCP1 a,b in BAT ↑ UCP3 a,b in SKM | ↓ body weight ↑ lean body mass ↓ total WAT weight 4 |
| Zhou, 2018, [142] | C57BL/6J mice # | 8/8 | EGCG, 1.0% w/w, orally | 4 | ↑ PRDM16 a, PGC-1α a and UCP1 a in BAT | ↓ body weight ↓ pWAT, eWAT, sWAT ↓ adipocyte size in WAT and lipid droplets in BAT ↑ thermogenic adaptations |
| First Author, Year, Reference | Experimental Model | Concentration or Dose | Compound | Potential Mechanism | Main Effects |
|---|---|---|---|---|---|
| Lagouge, 2006, [151] | C2C12 * | 50 µM | Resveratrol | ↑ SIRT1, PGC-1α, UCP1 | ↑ oxygen consumption, metabolic consumption |
| KKay mice | 400 mpk | ||||
| HF-fed C57Bl/6J mice | 200, 400 mpk | ||||
| Alberdi, 2013, [152] | HF-fed rat | 30 mpk | Resveratrol | ↑ SIRT1, PGC-1α, UCP1, TFAM, COX2, UCP3 (in muscles) | ↑ thermogenesis, whole-body energy dissipation |
| Li, 2020, [79] | 3T3-L1 * | 5–20 µM | Resveratrol | ↓ PPAR-γ, CEBPα ↑ SIRT1, ATGL, FNDC5, UCP1, PRDM16, CIDEA | ↑ Browning-like feature in white adipose tissue |
| KKay mice | 200 or 400 mpk | ||||
| HF-fed C57Bl/6J mice | 200 or 400 mpk | ||||
| Andrade, 2014, [153] | HF-fed C57Bl/6J mice | 400 mpk | Resveratrol | ↑ SIRT1, UCP1, PGC-1α | ↑ oxygen consumption ↓ epididymal and retroperitoneal adipose tissue, cholesterol, glucose |
| Zheng, 2016, [154] | HF-fed SIRT1 (+/−) mice | - | - | ↓ UCP1, mtDNA | ↓ oxygen consumption and heat production, UCP1, mtDNA ↑ white adipose tissue |
| Jimenez-Gomez, 2013, [155] | 3T3-L1 * | Serum of treated monkeys | Resveratrol | ↑ SIRT1, UCP1, PGC-1α | ↑ number of adipocytes ↓ inflammation |
| HF-fed rhesus monkey | 80 or 480 mpk | ||||
| Aziz, 2017, [156] | 3T3-L1 * | 10, 50, or 100 µM | Genistein | ↑ SIRT1, UCP1, PGC-1α | ↑ Browning-like feature in white adipose tissue, oxygen consumption |
| Gutiérrez-Salmeán, 2014, [157] | HF-fed mice | 1 mpk | Epicatechin | ↑ SIRT1, UCP1, PGC-1α | ↓ triglycerides, weight gain, cardiometabolic risk factors |
| Dong, 2014, [158] | HF-fed C57Bl/6J mice | 0.1 % w/w | Quercetin | ↑ SIRT1/AMPK, PGC-1α, UCP1 | ↑ Browning-like feature in white adipose tissue ↓ inflammation, body weight gain |
| Zhao, 2017, [159] | HF-fed rats | 120 mpk + 240 mpk | Resveratrol + quercetin | ↑ SIRT1/AMPK | ↓ inflammation, body weight gain, cholesterol, triglycerides |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flori, L.; Piragine, E.; Spezzini, J.; Citi, V.; Calderone, V.; Martelli, A. Influence of Polyphenols on Adipose Tissue: Sirtuins as Pivotal Players in the Browning Process. Int. J. Mol. Sci. 2023, 24, 9276. https://doi.org/10.3390/ijms24119276
Flori L, Piragine E, Spezzini J, Citi V, Calderone V, Martelli A. Influence of Polyphenols on Adipose Tissue: Sirtuins as Pivotal Players in the Browning Process. International Journal of Molecular Sciences. 2023; 24(11):9276. https://doi.org/10.3390/ijms24119276
Chicago/Turabian StyleFlori, Lorenzo, Eugenia Piragine, Jacopo Spezzini, Valentina Citi, Vincenzo Calderone, and Alma Martelli. 2023. "Influence of Polyphenols on Adipose Tissue: Sirtuins as Pivotal Players in the Browning Process" International Journal of Molecular Sciences 24, no. 11: 9276. https://doi.org/10.3390/ijms24119276
APA StyleFlori, L., Piragine, E., Spezzini, J., Citi, V., Calderone, V., & Martelli, A. (2023). Influence of Polyphenols on Adipose Tissue: Sirtuins as Pivotal Players in the Browning Process. International Journal of Molecular Sciences, 24(11), 9276. https://doi.org/10.3390/ijms24119276