
Metabolic Dysregulation Explains the Diverse Impacts of Obesity in Males and Females with Gastrointestinal Cancers

 , , , ,
, , , , 
 ,
, 


Abstract
:1. Introduction
2. Results
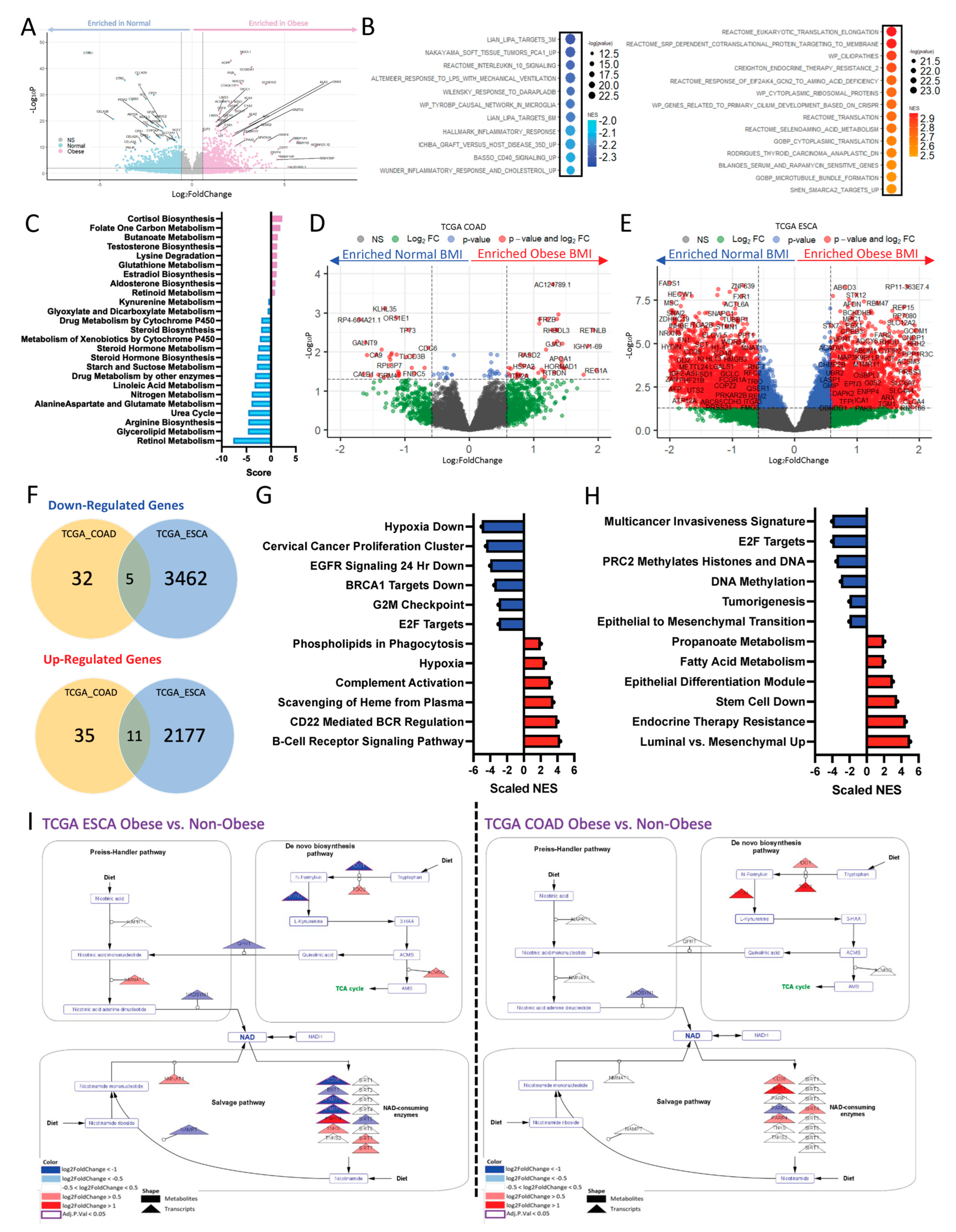
2.1. Obesity Drives Differential Transcriptomic Signature Associated with Metabolic Reprogramming in GI Cancers
2.2. Obesity Drives Differential Transcriptomic Signature in a Disease-Type-Specific Manner
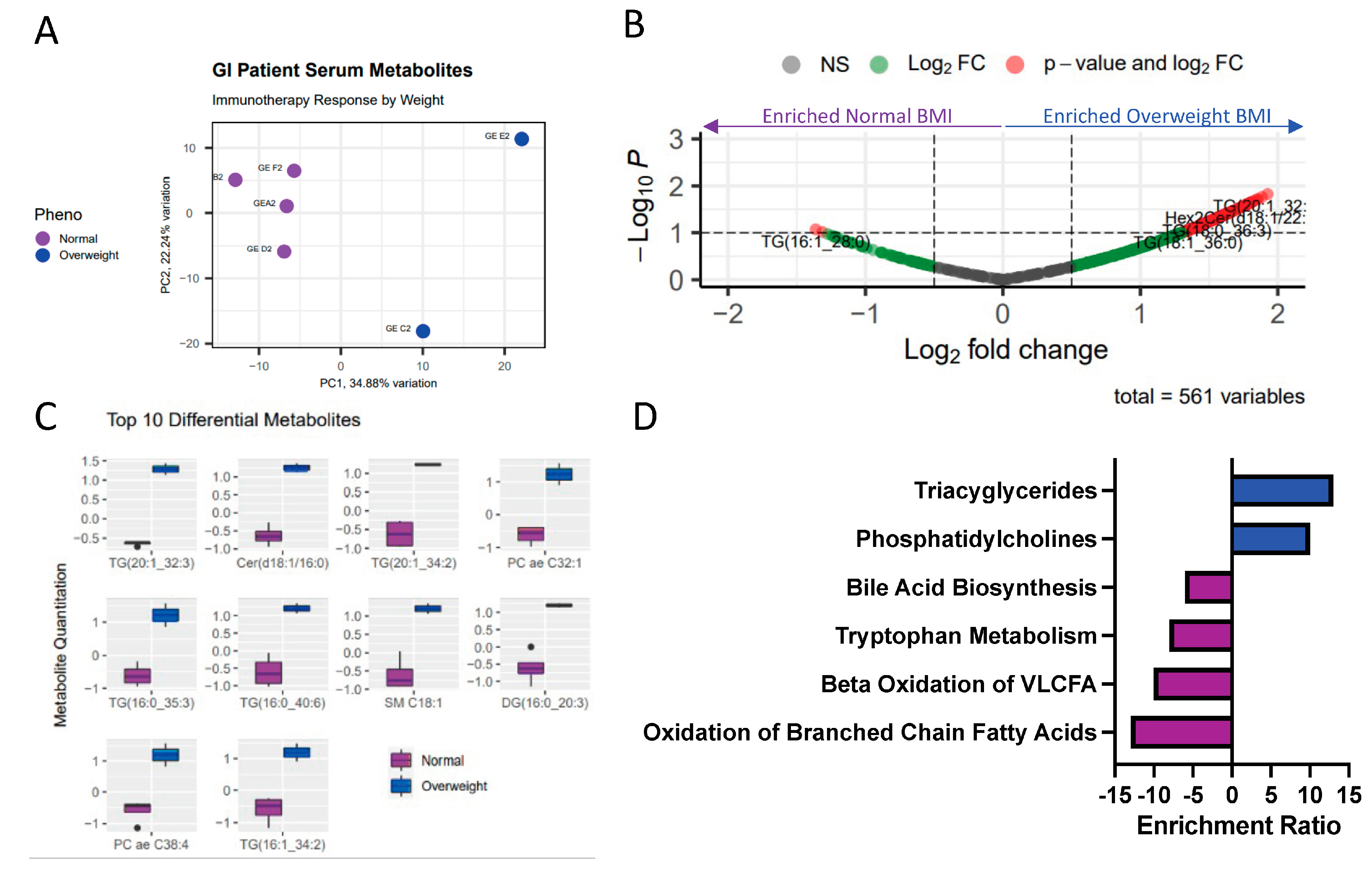
2.3. Obesity Associated with Differential Lipid and Amino Acid-Related Metabolic Programs
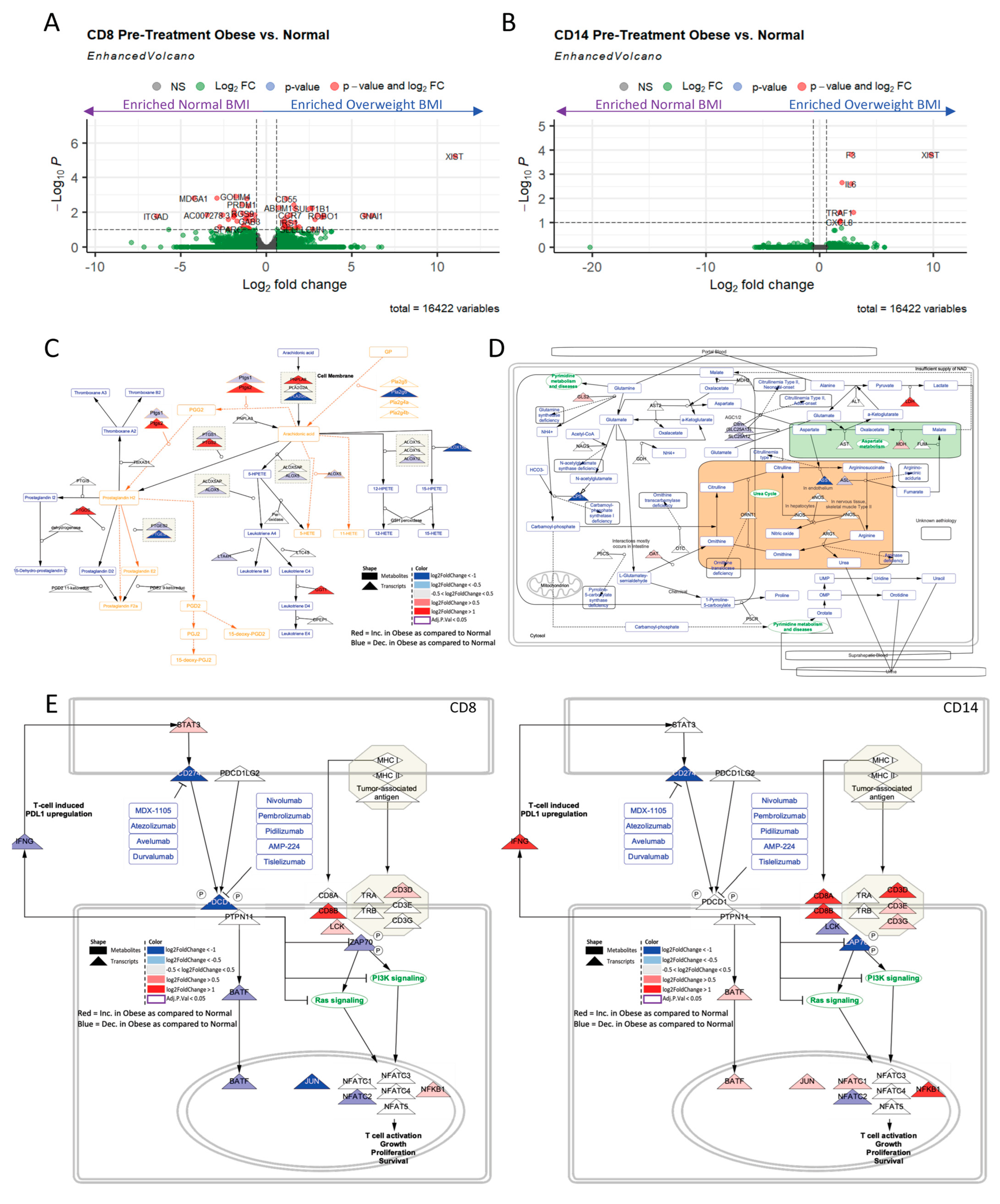
2.4. Obesity Drives Differential CD8+ and CD1+4 Metabolic and Immune Transcriptional Profiles Regardless of Sex
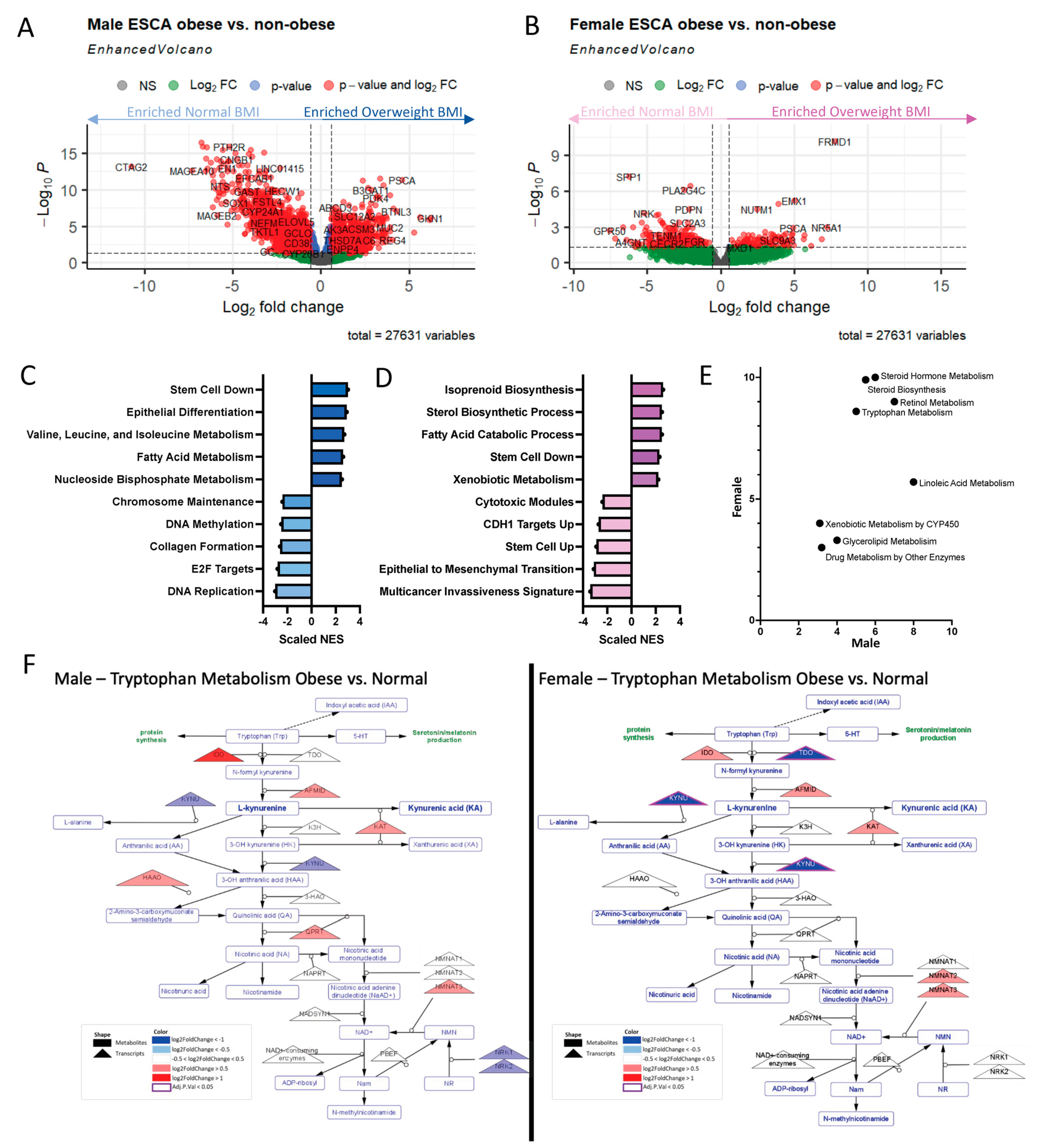
2.5. Obese Males and Females Dysregulate Similar Amino Acid and Lipid-Related Metabolic Pathways to Varying Extents
3. Discussion
4. Methods
4.1. Study Design
4.2. Sequencing Methods
4.3. The Cancer Genome Atlas Transcriptional Analysis
4.4. Metabolic Pipeline Analysis
4.5. Metabolomics—Biocrates Assays
4.6. Modeling with Cytoscape
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, S.E.; Llewellyn, C.H.; Smith, L. The obesity epidemic—Nature via nurture: A narrative review of high-income countries. SAGE Open Med. 2020, 8, 2050312120918265. [Google Scholar] [CrossRef] [PubMed]
- Kelly, T.; Yang, W.; Chen, C.-S.; Reynolds, K.; He, J. Global burden of obesity in 2005 and projections to 2030. Int. J. Obes. 2008, 32, 1431–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, S.; Chin, M.C.; Deurenberg-Yap, M. Review on epidemic of obesity. Ann. Acad. Med. Singap. 2009, 38, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Mokdad, A.H.; Ford, E.S.; Bowman, B.A.; Dietz, W.H.; Vinicor, F.; Bales, V.S.; Marks, J.S. Prevalence of Obesity, Diabetes, and Obesity-Related Health Risk Factors, 2001. JAMA 2003, 289, 76–79. [Google Scholar] [CrossRef]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K. Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, M.C.; Pastore, C.A.; Orlandi, S.P.; Heymsfield, S.B. Obesity paradox in cancer: New insights provided by body composition. Am. J. Clin. Nutr. 2014, 99, 999–1005. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Aguilar, E.G.; Luna, J.I.; Dunai, C.; Khuat, L.T.; Le, C.T.; Mirsoian, A.; Minnar, C.M.; Stoffel, K.M.; Sturgill, I.R.; et al. Paradoxical effects of obesity on T cell function during tumor progression and PD-1 checkpoint blockade. Nat. Med. 2018, 25, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Naik, G.S.; Waikar, S.S.; Johnson, A.E.W.; Buchbinder, E.I.; Haq, R.; Hodi, F.S.; Schoenfeld, J.D.; Ott, P.A. Complex inter-relationship of body mass index, gender and serum creatinine on survival: Exploring the obesity paradox in melanoma patients treated with checkpoint inhibition. J. Immunother. Cancer 2019, 7, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagheri, M.; Speakman, J.R.; Shemirani, F.; Djafarian, K. Renal cell carcinoma survival and body mass index: A dose–response meta-analysis reveals another potential paradox within a paradox. Int. J. Obes. 2016, 40, 1817–1822. [Google Scholar] [CrossRef]
- Toro, A.L.; Costantino, N.S.; Shriver, C.D.; Ellsworth, D.L.; Ellsworth, R.E. Effect of obesity on molecular characteristics of invasive breast tumors: Gene expression analysis in a large cohort of female patients. BMC Obes. 2016, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Naik, A.; Monjazeb, A.M.; Decock, J. The Obesity Paradox in Cancer, Tumor Immunology, and Immunotherapy: Potential Therapeutic Implications in Triple Negative Breast Cancer. Front. Immunol. 2019, 10, 1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanter, R.; Caballero, B. Global Gender Disparities in Obesity: A Review. Adv. Nutr. Int. Rev. J. 2012, 3, 491–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Y.; Ren, Y.; Zhang, Q.; Yi, P.; Cheng, C. Metabolic modulation of immune checkpoints and novel therapeutic strategies in cancer. Semin. Cancer Biol. 2022, 86, 542–565. [Google Scholar] [CrossRef] [PubMed]
- Cheung, O.K.-W.; Cheng, A.S.-L. Gender Differences in Adipocyte Metabolism and Liver Cancer Progression. Front. Genet. 2016, 7, 168. [Google Scholar] [CrossRef]
- Rosendahl, J.; Kirsten, H.; Hegyi, E.; Kovacs, P.; Weiss, F.U.; Laumen, H.; Lichtner, P.; Ruffert, C.; Chen, J.-M.; Masson, E.; et al. Genome-wide association study identifies inversion in the CTRB1-CTRB2 locus to modify risk for alcoholic and non-alcoholic chronic pancreatitis. Gut 2017, 67, 1855–1863. [Google Scholar] [CrossRef] [Green Version]
- Li, L.O.; Klett, E.L.; Coleman, R.A. Acyl-CoA synthesis, lipid metabolism and lipotoxicity. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2010, 1801, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayakumar, A.; Yakar, S.; LeRoith, D. The Intricate Role of Growth Hormone in Metabolism. Front. Endocrinol. 2011, 2, 32. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, A.; Sainz, J.; Hein, R.; Hoffmeister, M.; Frank, B.; Försti, A.; Brenner, H.; Hemminki, K.; Chang-Claude, J. Modification of menopausal hormone therapy-associated colorectal cancer risk by polymorphisms in sex steroid signaling, metabolism and transport related genes. Endocr.-Relat. Cancer 2011, 18, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Mabrouk, N.; Lecoeur, B.; Bettaieb, A.; Paul, C.; Végran, F. Impact of Lipid Metabolism on Antitumor Immune Response. Cancers 2022, 14, 1850. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Lei, Q.; Yang, L.; Qin, G.; Liu, S.; Wang, D.; Ping, Y.; Zhang, Y. Contradictory roles of lipid metabolism in immune response within the tumor microenvironment. J. Hematol. Oncol. 2021, 14, 187. [Google Scholar] [CrossRef]
- Ringel, A.E.; Drijvers, J.M.; Baker, G.J.; Catozzi, A.; García-Cañaveras, J.C.; Gassaway, B.M.; Miller, B.C.; Juneja, V.R.; Nguyen, T.H.; Joshi, S.; et al. Obesity Shapes Metabolism in the Tumor Microenvironment to Suppress Anti-Tumor Immunity. Cell 2020, 183, 1848–1866.e26. [Google Scholar] [CrossRef] [PubMed]
- McQuade, J.L.; Daniel, C.R.; Hess, K.R.; Mak, C.; Wang, D.Y.; Rai, R.R.; Park, J.J.; Haydu, L.E.; Spencer, C.; Wongchenko, M.; et al. Association of body-mass index and outcomes in patients with metastatic melanoma treated with targeted therapy, immunotherapy, or chemotherapy: A retrospective, multicohort analysis. Lancet Oncol. 2018, 19, 310–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vick, L.; Wang, Z.; Collins, C.; Khuat, L.; Dunai, C.; Stoffel, K.; Yendamuri, S.; Mukherjee, S.; Rosario, S.; Canter, R.; et al. 534 Sex-linked differences in obesity markedly impact the anti-tumor efficacy of PD-1 blockade. J. ImmunoTherapy Cancer 2022, 10. [Google Scholar] [CrossRef]
- Rong, J.; Chen, S.; Song, C.; Wang, H.; Zhao, Q.; Zhao, R.; He, Y.; Yan, L.; Song, Y.; Wang, F.; et al. The prognostic value of gender in gastric gastrointestinal stromal tumors: A propensity score matching analysis. Biol. Sex Differ. 2020, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Abancens, M.; Bustos, V.; Harvey, H.; McBryan, J.; Harvey, B.J. Sexual Dimorphism in Colon Cancer. Front. Oncol. 2020, 10, 607909. [Google Scholar] [CrossRef]
- Wang, S.; Cowley, L.A.; Liu, X.-S. Sex Differences in Cancer Immunotherapy Efficacy, Biomarkers, and Therapeutic Strategy. Molecules 2019, 24, 3214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosario, S.R.; Long, M.D.; Affronti, H.C.; Rowsam, A.M.; Eng, K.H.; Smiraglia, D.J. Pan-cancer analysis of transcriptional metabolic dysregulation using The Cancer Genome Atlas. Nat. Commun. 2018, 9, 5330. [Google Scholar] [CrossRef] [Green Version]
- Rosario, S.R.; Smith RJJr Patnaik, S.K.; Liu, S.; Barbi, J.; Yendamuri, S. Altered acetyl-CoA metabolism presents a new potential immunotherapy target in the obese lung microenvironment. Cancer Metab. 2022, 10, 17. [Google Scholar] [CrossRef]
- Clish, C.B. Metabolomics: An emerging but powerful tool for precision medicine. Mol. Case Stud. 2015, 1, a000588. [Google Scholar] [CrossRef] [Green Version]
- Qian, B.-Z.; Pollard, J.W. Macrophage Diversity Enhances Tumor Progression and Metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castoldi, A.; Naffah de Souza, C.; Camara, N.O.S.; Moraes-Vieira, P.M. The Macrophage Switch in Obesity Development. Front. Immunol. 2015, 6, 637. [Google Scholar] [CrossRef] [Green Version]
- Stine, Z.E.; Schug, Z.T.; Salvino, J.M.; Dang, C.V. Targeting cancer metabolism in the era of precision oncology. Nat. Rev. Drug Discov. 2021, 21, 141–162. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdóttir, H.; Tamayo, P.; Mesirov, J.P. Molecular signatures database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [Green Version]
- Killcoyne, S.; Carter, G.W.; Smith, J.; Boyle, J. Cytoscape: A Community-Based Framework for Network Modeling. Method Mol. Biol. 2009, 563, 219–239. [Google Scholar]
- Slenter, D.N.; Kutmon, M.; Hanspers, K.; Riutta, A.; Windsor, J.; Nunes, N.; Mélius, J.; Cirillo, E.; Coort, S.L.; Digles, D.; et al. WikiPathways: A multifaceted pathway database bridging metabolomics to other omics research. Nucleic Acids Res. 2018, 46, D661–D667. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORIEN CRC | |||
|---|---|---|---|
| Variable | Age > 50 | Age <= 50 | p |
| n = 364 (n%) | n = 96 (n%) | ||
| Race | 0.621 | ||
| White | 313 (86) | 80 (83) | |
| Non-white | 51(14) | 16 (17) | |
| Not reported | 0 (0) | 0 (0) | |
| Gender | 0.801 | ||
| Female | 163 (45) | 45 (47) | |
| Male | 201 (55) | 51 (53) | |
| Tumor stage | 0.009 | ||
| I (IA, IB) | 21 (5) | 3 (3) | |
| II (IIA/B/C) | 90 (24) | 10 (10) | |
| III (IIIA/B/C)+ | 104 (29) | 34 (35) | |
| IV (IVA/B/C)+ | 87 (24) | 31 (32) | |
| Not reported | 62 (17) | 18 (18) | |
| BMI | 0.624 | ||
| BMI < 18 | 4 (1) | 1 (1) | |
| BMI 18–24.9 | 70 (19) | 24 (25) | |
| BMI 24.9–30 | 111 (30) | 25 (26) | |
| BMI >= 30 | 109 (30) | 28 (29) | |
| Missing | 70 (19) | 18 (18) | |
| Characteristics by Gender in TCGA-COAD, 438 Cases | ||||
|---|---|---|---|---|
| Variable | Levels | Female (n = 208) | Males (n = 230) | p |
| race | asian | 2 (1) | 5 (2) | 0.4 |
| black or african american | 31 (15) | 28 (12) | ||
| not reported | 71 (34) | 92 (40) | ||
| white | 104 (50) | 1005 (46) | ||
| tumor stage | I (IA, IB) | 33 (16) | 42 (18) | 0.6 |
| II (IIA, IIB) | 70 (34) | 66 (29) | ||
| III (IIIA, /B/B)+ | 100 (48) | 115 (50) | ||
| no report | 5 (2) | 7 (3) | ||
| bmicat2 | BMI >= 30 | 41 (36) | 37 (33) | 0.3 |
| BMI 18–24.9 | 39 (34) | 32 (28) | ||
| BMI 24.9–30 | 34 (30) | 44 (39) | ||
| median (range) | ||||
| Age at initial diagnosis | 67 (34–90) | 68 (31–90) | 0.2 | |
| Characteristics by Gender in TCGA-ESCA, 87 Cases | ||||
| Variable | Levels | Female (n = 13) | Males (n = 74) | p |
| race | asian | 0 (0) | 1 (1) | 0.9 |
| black or african american | 0 (0) | 0 (0) | ||
| not reported | 2 (15) | 14 (19) | ||
| white | 11 (85) | 59 (80) | ||
| tumor stage | I (IA,IB) | 6 (46) | 7 (9) | 0.007 |
| II (IIA,IIB) | 4 (31) | 20 (27) | ||
| III (IIIA,/B/B)+ | 2 (15) | 31 (42) | ||
| no report | 1 (8) | 16 (22) | ||
| bmicat2 | BMI >= 30 | 4 (33) | 19 (27) | 0.8 |
| BMI 18–24.9 | 4 (33) | 31 (44) | ||
| BMI 24.9–30 | 4 (33) | 44 (39) | ||
| median (range) | ||||
| Age at initial diagnosis | 74 (28–84) | 65 (28–84) | 0.03 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosario, S.R.; Dong, B.; Zhang, Y.; Hsiao, H.-H.; Isenhart, E.; Wang, J.; Siegel, E.M.; Monjazeb, A.M.; Owen, D.H.; Dey, P.; et al. Metabolic Dysregulation Explains the Diverse Impacts of Obesity in Males and Females with Gastrointestinal Cancers. Int. J. Mol. Sci. 2023, 24, 10847. https://doi.org/10.3390/ijms241310847
Rosario SR, Dong B, Zhang Y, Hsiao H-H, Isenhart E, Wang J, Siegel EM, Monjazeb AM, Owen DH, Dey P, et al. Metabolic Dysregulation Explains the Diverse Impacts of Obesity in Males and Females with Gastrointestinal Cancers. International Journal of Molecular Sciences. 2023; 24(13):10847. https://doi.org/10.3390/ijms241310847
Chicago/Turabian StyleRosario, Spencer R., Bowen Dong, Yali Zhang, Hua-Hsin Hsiao, Emily Isenhart, Jianmin Wang, Erin M. Siegel, Arta M. Monjazeb, Dwight H. Owen, Prasenjit Dey, and et al. 2023. "Metabolic Dysregulation Explains the Diverse Impacts of Obesity in Males and Females with Gastrointestinal Cancers" International Journal of Molecular Sciences 24, no. 13: 10847. https://doi.org/10.3390/ijms241310847
APA StyleRosario, S. R., Dong, B., Zhang, Y., Hsiao, H. -H., Isenhart, E., Wang, J., Siegel, E. M., Monjazeb, A. M., Owen, D. H., Dey, P., Tabung, F. K., Spakowicz, D. J., Murphy, W. J., Edge, S., Yendamuri, S., Ibrahimi, S., Kolesar, J. M., McDonald, P. H., Vadehra, D., ... Mukherjee, S. (2023). Metabolic Dysregulation Explains the Diverse Impacts of Obesity in Males and Females with Gastrointestinal Cancers. International Journal of Molecular Sciences, 24(13), 10847. https://doi.org/10.3390/ijms241310847