

Cell-Type Specific Regulation of Cholesterogenesis by CYP46A1 Re-Expression in zQ175 HD Mouse Striatum
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
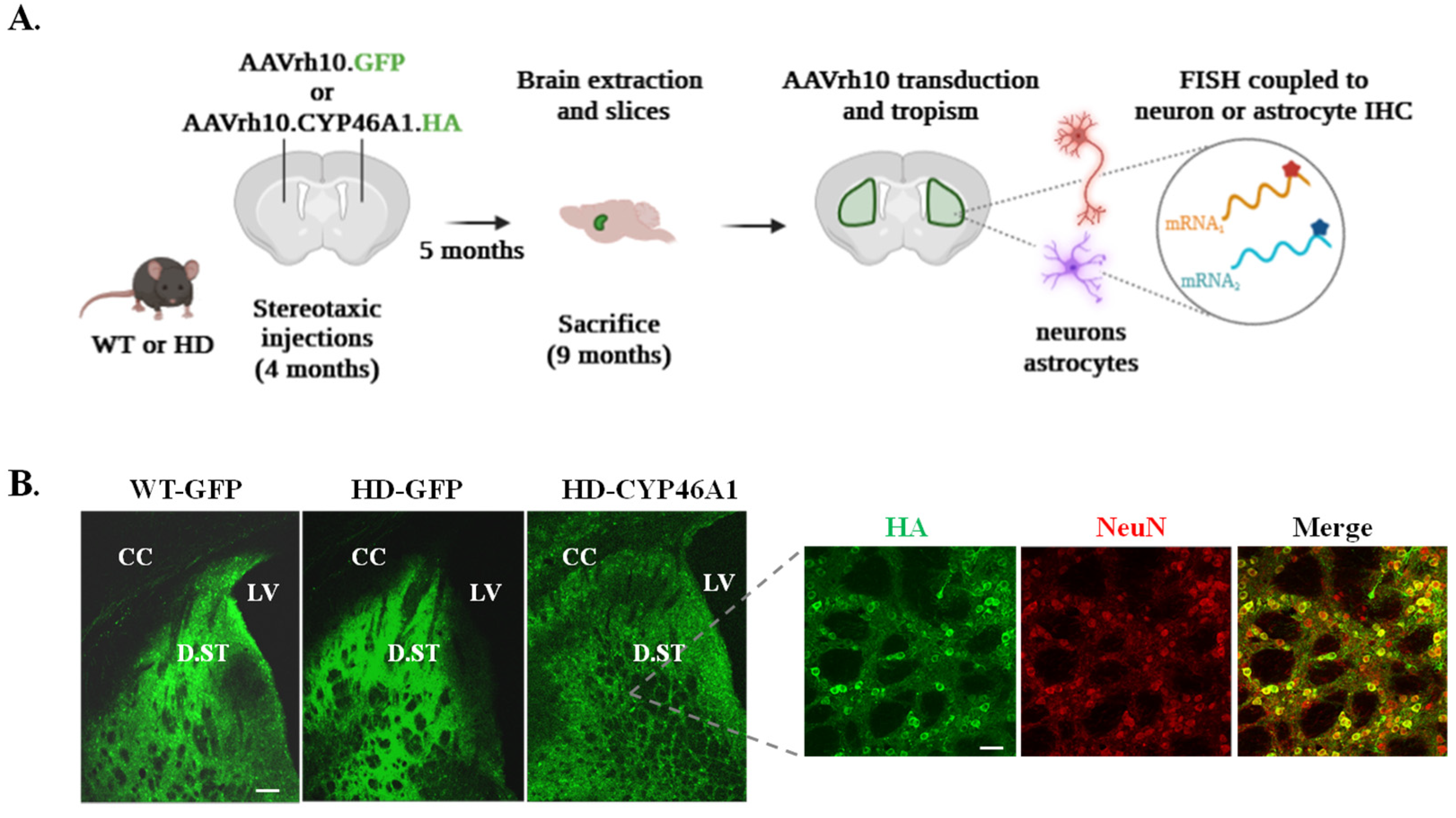
2.1. Validation of Virus-Mediated Expression of GFP and CYP46A1-HA in Dorsal Striatum and Validation of FISH Analysis Coupled with Cell-Specific Immunolabelling
2.1.1. Expression of CYP46A1 in the Striatum of HD Mice
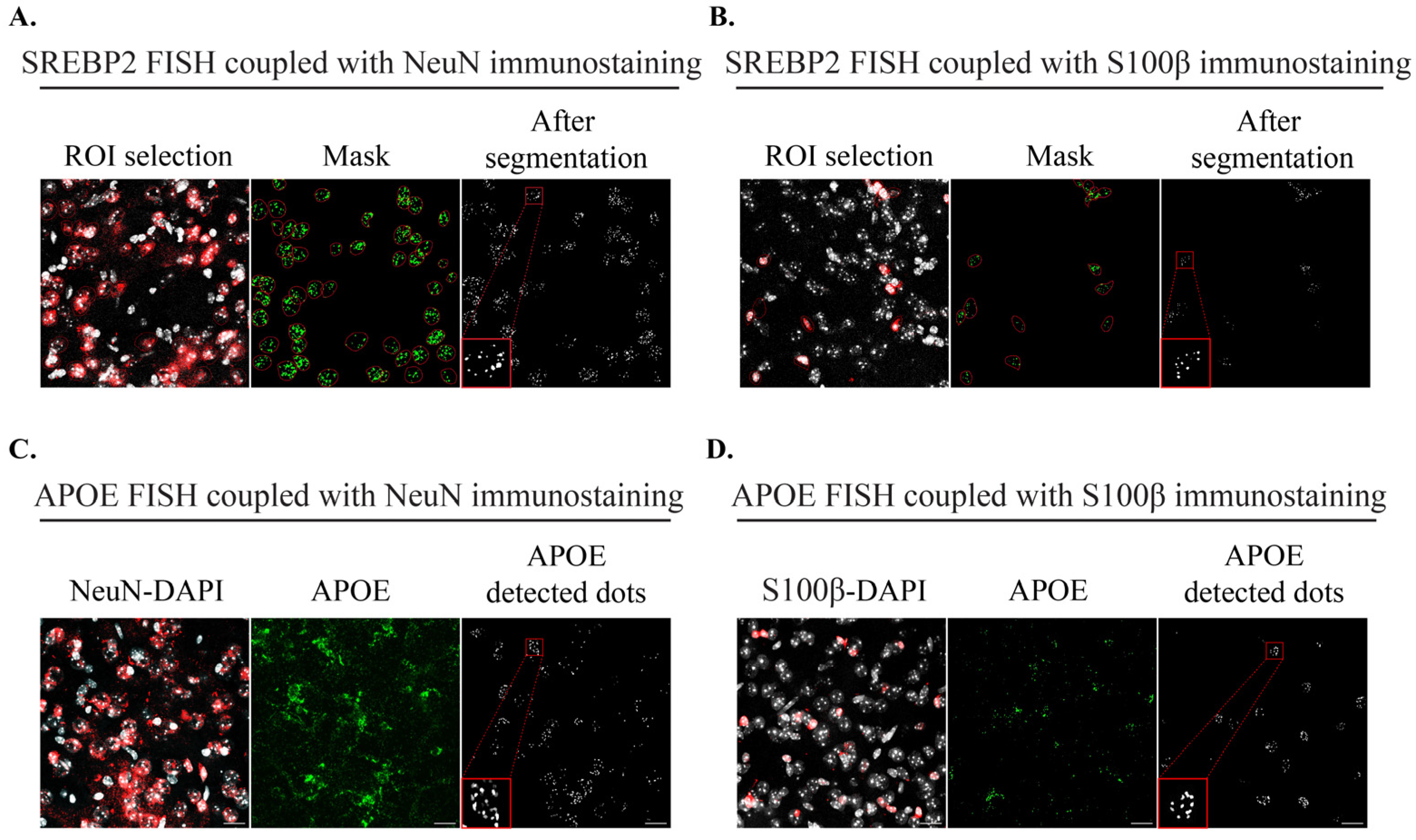
2.1.2. Imaging Tools to Quantify mRNA Signals after FISH Coupled with IHC
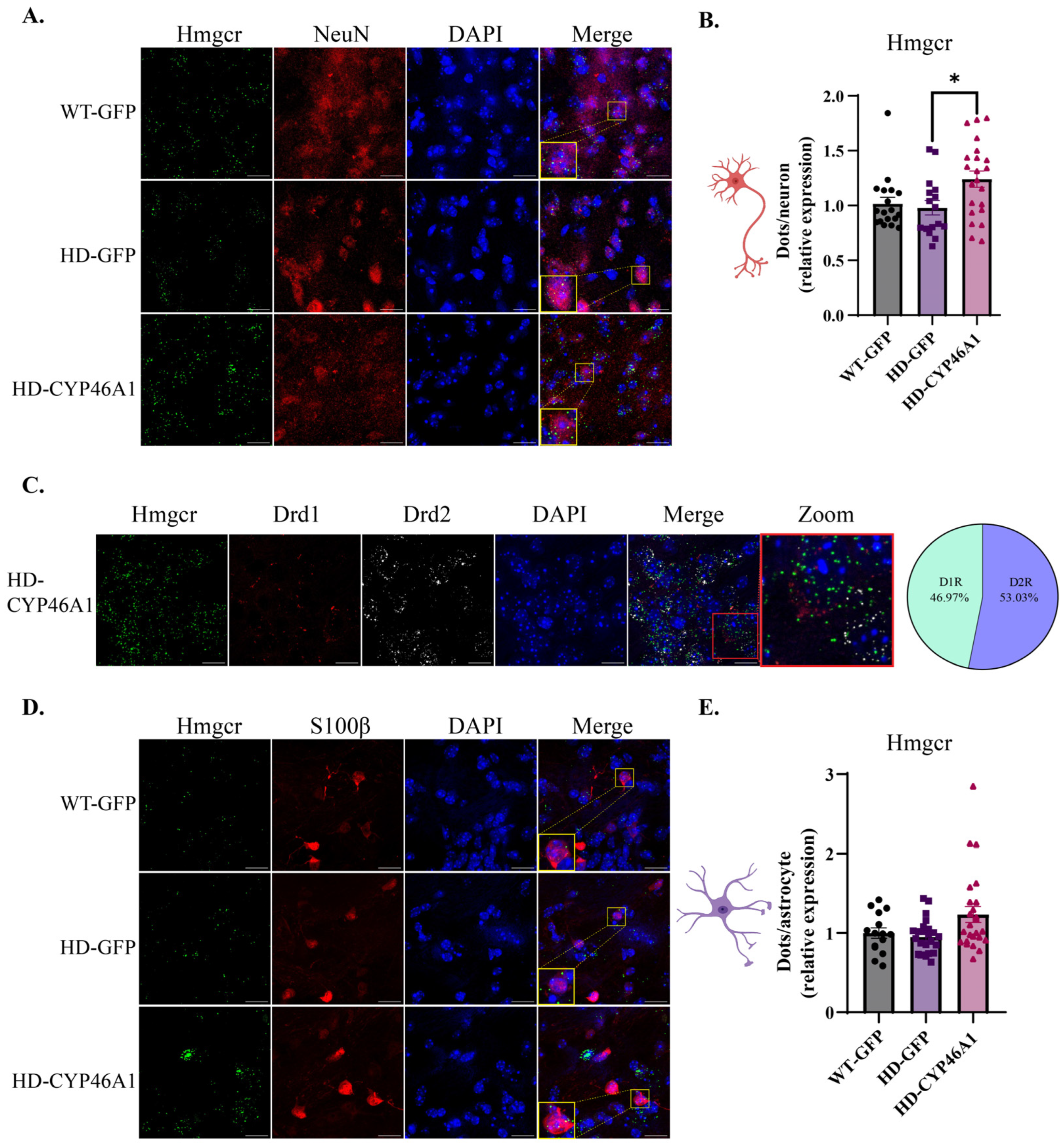
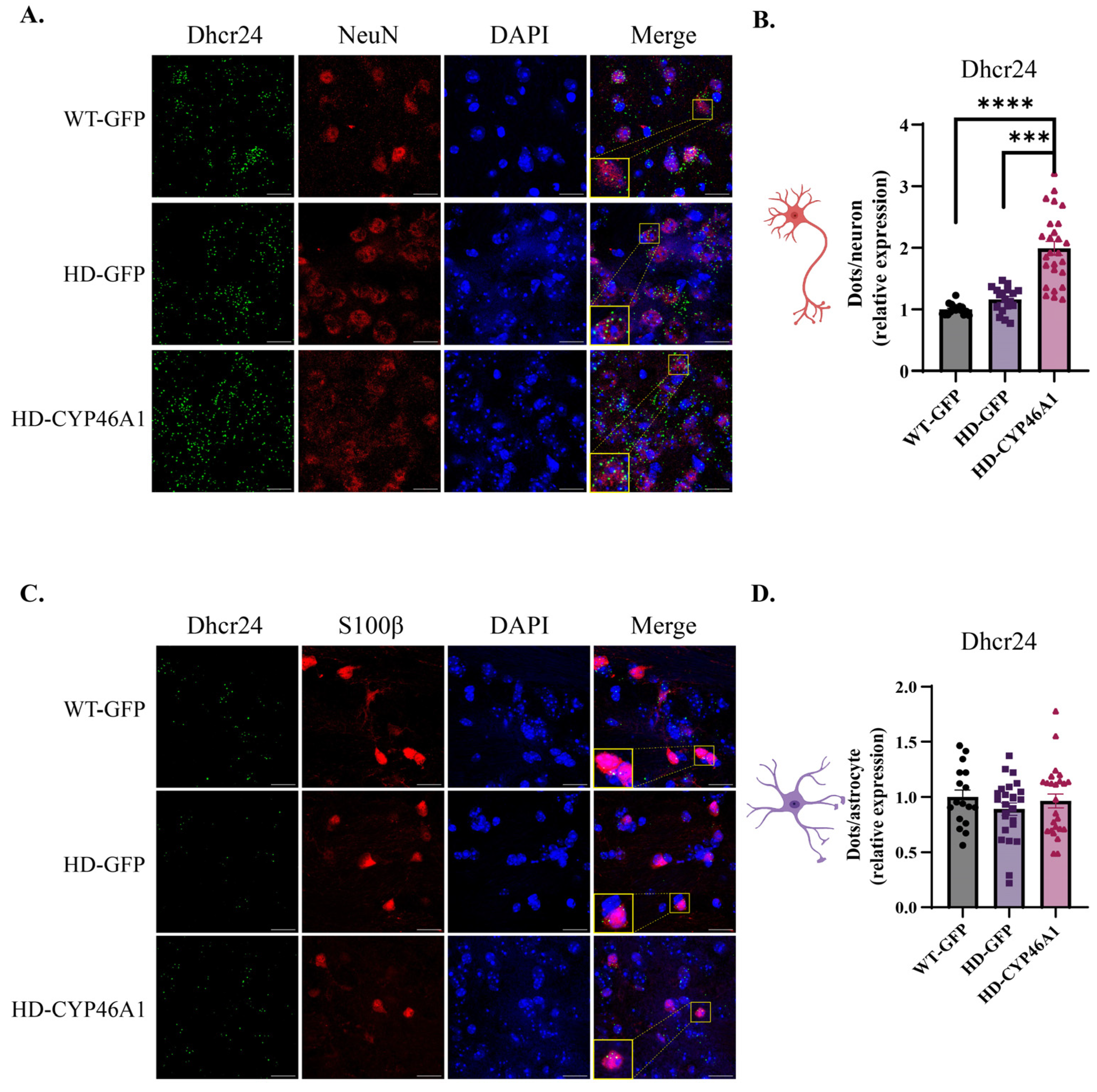
2.2. Analysis of Hmgcr and Dhcr24, Two Key Cholesterol Synthesis Genes, in Neurons and Astrocytes after CYP46A1-HA Expression in HD Mice
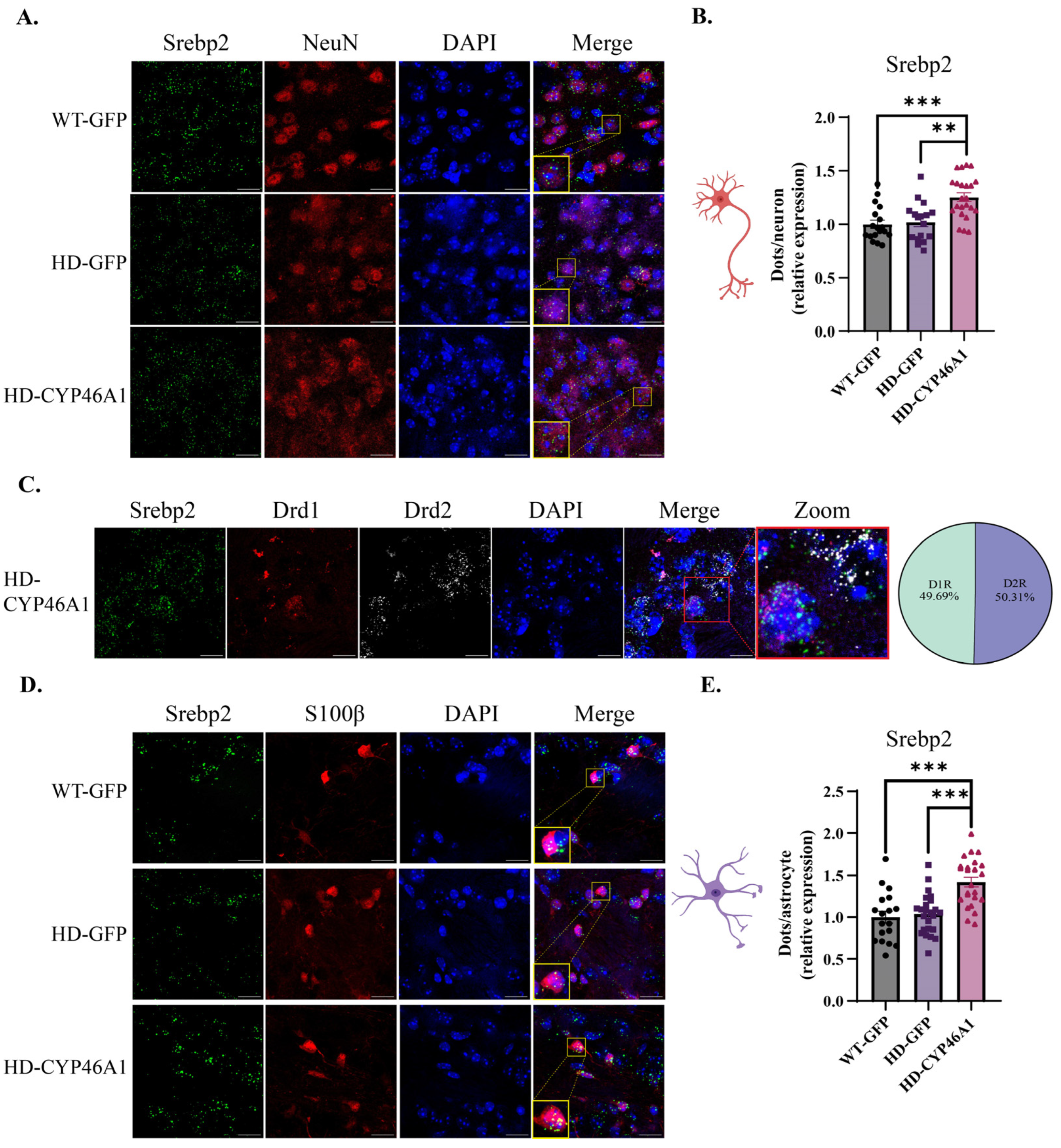
2.3. Regulation of Srebp2 Transcription Factor Gene in Neurons and Astrocytes after CYP46A1-HA Expression in HD Mice
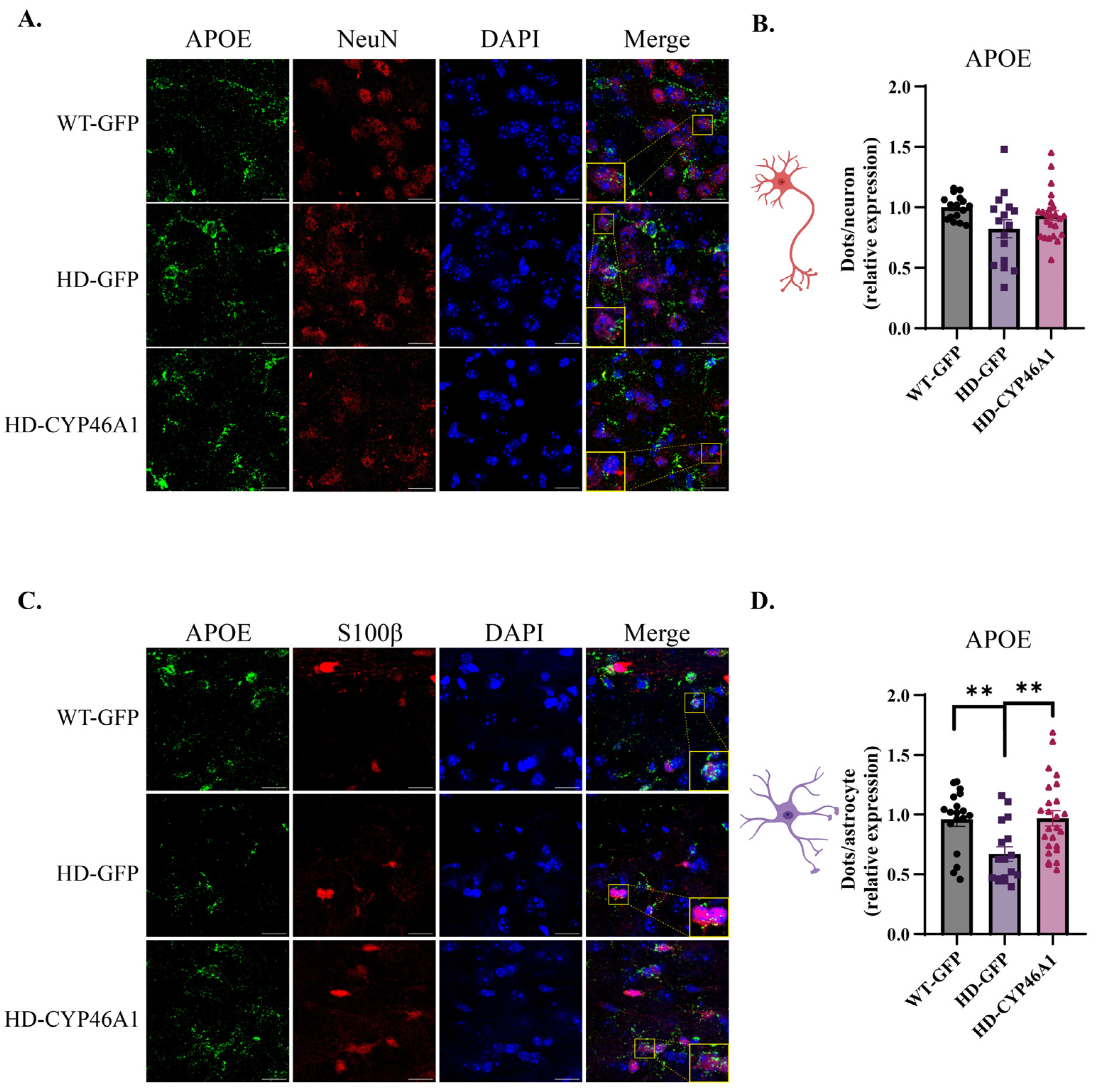
2.4. Neuronal CYP46A1 Expression Effect on ApoE Cholesterol Efflux Gene
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Production and Stereotaxic Injection of AAVrh10.GFP and AAVrh10.CYP46A1.HA
4.3. Brain Section Preparation
4.4. Immunostaining
4.5. FISH Coupled with Immunostaining
4.6. Image Acquisition and Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aylward, E.H.; Codori, A.M.; Rosenblatt, A.; Sherr, M.; Brandt, J.; Stine, O.C.; Barta, P.E.; Pearlson, G.D.; Ross, C.A. Rate of Caudate Atrophy in Presymptomatic and Symptomatic Stages of Huntington’s Disease. Mov. Disord. 2000, 15, 552–560. [Google Scholar] [CrossRef]
- MacDonald, M.E.; Ambrose, C.M.; Duyao, M.P.; Myers, R.H.; Lin, C.; Srinidhi, L.; Barnes, G.; Taylor, S.A.; James, M.; Groot, N.; et al. A Novel Gene Containing a Trinucleotide Repeat that Is Expanded and Unstable on Huntington’s Disease Chromosomes. The Huntington’s Disease Collaborative Research Group. Cell 1993, 72, 971–983. [Google Scholar] [CrossRef]
- Saudou, F.; Humbert, S. The Biology of Huntingtin. Neuron 2016, 89, 910–926. [Google Scholar] [CrossRef] [Green Version]
- Tabrizi, S.J.; Flower, M.D.; Ross, C.A.; Wild, E.J. Huntington Disease: New Insights into Molecular Pathogenesis and Therapeutic Opportunities. Nat. Rev. Neurol. 2020, 16, 529–546. [Google Scholar] [CrossRef]
- Duan, W.; Urani, E.; Mattson, M.P. The Potential of Gene Editing for Huntington’s Disease. Trends Neurosci. 2023, 46, 365–376. [Google Scholar] [CrossRef]
- Tabrizi, S.J.; Estevez-Fraga, C.; van Roon-Mom, W.M.C.; Flower, M.D.; Scahill, R.I.; Wild, E.J.; Muñoz-Sanjuan, I.; Sampaio, C.; Rosser, A.E.; Leavitt, B.R. Potential Disease-Modifying Therapies for Huntington’s Disease: Lessons Learned and Future Opportunities. Lancet Neurol. 2022, 21, 645–658. [Google Scholar] [CrossRef]
- Kacher, R.; Mounier, C.; Caboche, J.; Betuing, S. Altered Cholesterol Homeostasis in Huntington’s Disease. Front. Aging Neurosci. 2022, 14, 797220. [Google Scholar] [CrossRef]
- Valenza, M.; Cattaneo, E. Emerging Roles for Cholesterol in Huntington’s Disease. Trends Neurosci. 2011, 34, 474–486. [Google Scholar] [CrossRef]
- Lund, E.G.; Guileyardo, J.M.; Russell, D.W. CDNA Cloning of Cholesterol 24-Hydroxylase, a Mediator of Cholesterol Homeostasis in the Brain. Proc. Natl. Acad. Sci. USA 1999, 96, 7238–7243. [Google Scholar] [CrossRef]
- Leoni, V.; Mariotti, C.; Tabrizi, S.J.; Valenza, M.; Wild, E.J.; Henley, S.M.D.; Hobbs, N.Z.; Mandelli, M.L.; Grisoli, M.; Björkhem, I.; et al. Plasma 24S-Hydroxycholesterol and Caudate MRI in Pre-Manifest and Early Huntington’s Disease. Brain 2008, 131, 2851–2859. [Google Scholar] [CrossRef] [Green Version]
- Leoni, V.; Long, J.D.; Mills, J.A.; Di Donato, S.; Paulsen, J.S.; PREDICT-HD Study Group. Plasma 24S-Hydroxycholesterol Correlation with Markers of Huntington Disease Progression. Neurobiol. Dis. 2013, 55, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Valenza, M.; Leoni, V.; Tarditi, A.; Mariotti, C.; Björkhem, I.; Di Donato, S.; Cattaneo, E. Progressive Dysfunction of the Cholesterol Biosynthesis Pathway in the R6/2 Mouse Model of Huntington’s Disease. Neurobiol. Dis. 2007, 28, 133–142. [Google Scholar] [CrossRef]
- Valenza, M.; Carroll, J.B.; Leoni, V.; Bertram, L.N.; Björkhem, I.; Singaraja, R.R.; Di Donato, S.; Lutjohann, D.; Hayden, M.R.; Cattaneo, E. Cholesterol Biosynthesis Pathway Is Disturbed in YAC128 Mice and Is Modulated by Huntingtin Mutation. Hum. Mol. Genet. 2007, 16, 2187–2198. [Google Scholar] [CrossRef] [Green Version]
- Leoni, V.; Mariotti, C.; Nanetti, L.; Salvatore, E.; Squitieri, F.; Bentivoglio, A.R.; Bandettini di Poggio, M.; Bandettini Del Poggio, M.; Piacentini, S.; Monza, D.; et al. Whole Body Cholesterol Metabolism Is Impaired in Huntington’s Disease. Neurosci. Lett. 2011, 494, 245–249. [Google Scholar] [CrossRef]
- Kreilaus, F.; Spiro, A.S.; Hannan, A.J.; Garner, B.; Jenner, A.M. Brain Cholesterol Synthesis and Metabolism Is Progressively Disturbed in the R6/1 Mouse Model of Huntington’s Disease: A Targeted GC-MS/MS Sterol Analysis. J. Huntingt. Dis. 2015, 4, 305–318. [Google Scholar] [CrossRef] [Green Version]
- Boussicault, L.; Alves, S.; Lamazière, A.; Planques, A.; Heck, N.; Moumné, L.; Despres, G.; Bolte, S.; Hu, A.; Pagès, C.; et al. CYP46A1, the Rate-Limiting Enzyme for Cholesterol Degradation, Is Neuroprotective in Huntington’s Disease. Brain 2016, 139, 953–970. [Google Scholar] [CrossRef] [Green Version]
- Kacher, R.; Lamazière, A.; Heck, N.; Kappes, V.; Mounier, C.; Despres, G.; Dembitskaya, Y.; Perrin, E.; Christaller, W.; Sasidharan Nair, S.; et al. CYP46A1 Gene Therapy Deciphers the Role of Brain Cholesterol Metabolism in Huntington’s Disease. Brain 2019, 142, 2432–2450. [Google Scholar] [CrossRef]
- Sipione, S.; Rigamonti, D.; Valenza, M.; Zuccato, C.; Conti, L.; Pritchard, J.; Kooperberg, C.; Olson, J.M.; Cattaneo, E. Early Transcriptional Profiles in Huntingtin-Inducible Striatal Cells by Microarray Analyses. Hum. Mol. Genet. 2002, 11, 1953–1965. [Google Scholar] [CrossRef] [Green Version]
- Valenza, M.; Rigamonti, D.; Goffredo, D.; Zuccato, C.; Fenu, S.; Jamot, L.; Strand, A.; Tarditi, A.; Woodman, B.; Racchi, M.; et al. Dysfunction of the Cholesterol Biosynthetic Pathway in Huntington’s Disease. J. Neurosci. 2005, 25, 9932–9939. [Google Scholar] [CrossRef] [Green Version]
- Samara, A.; Galbiati, M.; Luciani, P.; Deledda, C.; Messi, E.; Peri, A.; Maggi, R. Altered Expression of 3-Betahydroxysterol Delta-24-Reductase/Selective Alzheimer’s Disease Indicator-1 Gene in Huntington’s Disease Models. J. Endocrinol. Investig. 2014, 37, 729–737. [Google Scholar] [CrossRef]
- Valenza, M.; Marullo, M.; Di Paolo, E.; Cesana, E.; Zuccato, C.; Biella, G.; Cattaneo, E. Disruption of Astrocyte-Neuron Cholesterol Cross Talk Affects Neuronal Function in Huntington’s Disease. Cell Death Differ. 2015, 22, 690–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abildayeva, K.; Jansen, P.J.; Hirsch-Reinshagen, V.; Bloks, V.W.; Bakker, A.H.F.; Ramaekers, F.C.S.; de Vente, J.; Groen, A.K.; Wellington, C.L.; Kuipers, F.; et al. 24(S)-Hydroxycholesterol Participates in a Liver X Receptor-Controlled Pathway in Astrocytes That Regulates Apolipoprotein E-Mediated Cholesterol Efflux. J. Biol. Chem. 2006, 281, 12799–12808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangi, A.F.; Schnabel, J.A.; Davatzikos, C.; Alberola-López, C.; Fichtinger, G. Medical Image Computing and Computer Assisted Intervention–MICCAI 2018. In Proceedings of the 21st International Conference, Granada, Spain, 16–20 September 2018; Springer: Berlin/Heidelberg, Germany, 2018; Volume 11073. [Google Scholar]
- Heck, N.; Dos Santos, M.; Amairi, B.; Salery, M.; Besnard, A.; Herzog, E.; Boudier, T.; Vanhoutte, P.; Caboche, J. A New Automated 3D Detection of Synaptic Contacts Reveals the Formation of Cortico-Striatal Synapses upon Cocaine Treatment in Vivo. Brain Struct. Funct. 2015, 220, 2953–2966. [Google Scholar] [CrossRef] [PubMed]
- Ollion, J.; Cochennec, J.; Loll, F.; Escudé, C.; Boudier, T. TANGO: A Generic Tool for High-Throughput 3D Image Analysis for Studying Nuclear Organization. Bioinformatics 2013, 29, 1840–1841. [Google Scholar] [CrossRef] [Green Version]
- Surmeier, D.J.; Song, W.J.; Yan, Z. Coordinated Expression of Dopamine Receptors in Neostriatal Medium Spiny Neurons. J. Neurosci. 1996, 16, 6579–6591. [Google Scholar] [CrossRef] [Green Version]
- Goodliffe, J.W.; Song, H.; Rubakovic, A.; Chang, W.; Medalla, M.; Weaver, C.M.; Luebke, J.I. Differential Changes to D1 and D2 Medium Spiny Neurons in the 12-Month-Old Q175+/- Mouse Model of Huntington’s Disease. PLoS ONE 2018, 13, e0200626. [Google Scholar] [CrossRef]
- Megret, L.; Gris, B.; Sasidharan Nair, S.; Cevost, J.; Wertz, M.; Aaronson, J.; Rosinski, J.; Vogt, T.F.; Wilkinson, H.; Heiman, M.; et al. Shape Deformation Analysis Reveals the Temporal Dynamics of Cell-Type-Specific Homeostatic and Pathogenic Responses to Mutant Huntingtin. Elife 2021, 10, e64984. [Google Scholar] [CrossRef]
- Matsushima, A.; Pineda, S.S.; Crittenden, J.R.; Lee, H.; Galani, K.; Mantero, J.; Tombaugh, G.; Kellis, M.; Heiman, M.; Graybiel, A.M. Transcriptional Vulnerabilities of Striatal Neurons in Human and Rodent Models of Huntington’s Disease. Nat. Commun. 2023, 14, 282. [Google Scholar] [CrossRef]
- Jeon, T.-I.; Osborne, T.F. SREBPs: Metabolic Integrators in Physiology and Metabolism. Trends Endocrinol. Metab. 2012, 23, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Birolini, G.; Verlengia, G.; Talpo, F.; Maniezzi, C.; Zentilin, L.; Giacca, M.; Conforti, P.; Cordiglieri, C.; Caccia, C.; Leoni, V.; et al. SREBP2 Gene Therapy Targeting Striatal Astrocytes Ameliorates Huntington’s Disease Phenotypes. Brain 2021, 144, 3175–3190. [Google Scholar] [CrossRef]
- Zhou, X.; Fu, A.K.; Ip, N.Y. APOE Signaling in Neurodegenerative Diseases: An Integrative Approach Targeting APOE Coding and Noncoding Variants for Disease Intervention. Curr. Opin. Neurobiol. 2021, 69, 58–67. [Google Scholar] [CrossRef]
- Valenza, M.; Leoni, V.; Karasinska, J.M.; Petricca, L.; Fan, J.; Carroll, J.; Pouladi, M.A.; Fossale, E.; Nguyen, H.P.; Riess, O.; et al. Cholesterol Defect Is Marked across Multiple Rodent Models of Huntington’s Disease and Is Manifest in Astrocytes. J. Neurosci. 2010, 30, 10844–10850. [Google Scholar] [CrossRef] [Green Version]
- Pikuleva, I.A.; Cartier, N. Cholesterol Hydroxylating Cytochrome P450 46A1: From Mechanisms of Action to Clinical Applications. Front. Aging Neurosci. 2021, 13, 696778. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.-C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.-T.; Ma, X.-J.; Luo, Y. RNAscope: A Novel in Situ RNA Analysis Platform for Formalin-Fixed, Paraffin-Embedded Tissues. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Fenster, R.J.; Pineda, S.S.; Gibbs, W.S.; Mohammadi, S.; Davila-Velderrain, J.; Garcia, F.J.; Therrien, M.; Novis, H.S.; Gao, F.; et al. Cell Type-Specific Transcriptomics Reveals That Mutant Huntingtin Leads to Mitochondrial RNA Release and Neuronal Innate Immune Activation. Neuron 2020, 107, 891–908. [Google Scholar] [CrossRef] [PubMed]
- Benraiss, A.; Mariani, J.N.; Osipovitch, M.; Cornwell, A.; Windrem, M.S.; Villanueva, C.B.; Chandler-Militello, D.; Goldman, S.A. Cell-Intrinsic Glial Pathology Is Conserved across Human and Murine Models of Huntington’s Disease. Cell Rep. 2021, 36, 109308. [Google Scholar] [CrossRef] [PubMed]
- Gangwani, M.R.; Soto, J.S.; Jami-Alahmadi, Y.; Tiwari, S.; Kawaguchi, R.; Wohlschlegel, J.A.; Khakh, B.S. Neuronal and Astrocytic Contributions to Huntington’s Disease Dissected with Zinc Finger Protein Transcriptional Repressors. Cell Rep. 2023, 42, 111953. [Google Scholar] [CrossRef] [PubMed]
- Boyles, J.K.; Pitas, R.E.; Wilson, E.; Mahley, R.W.; Taylor, J.M. Apolipoprotein E Associated with Astrocytic Glia of the Central Nervous System and with Nonmyelinating Glia of the Peripheral Nervous System. J. Clin. Investig. 1985, 76, 1501–1513. [Google Scholar] [CrossRef] [Green Version]
- Pitas, R.E.; Boyles, J.K.; Lee, S.H.; Foss, D.; Mahley, R.W. Astrocytes Synthesize Apolipoprotein E and Metabolize Apolipoprotein E-Containing Lipoproteins. Biochim. Biophys. Acta 1987, 917, 148–161. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinchaud, K.; Masson, C.; Dayre, B.; Mounier, C.; Gilles, J.-F.; Vanhoutte, P.; Caboche, J.; Betuing, S. Cell-Type Specific Regulation of Cholesterogenesis by CYP46A1 Re-Expression in zQ175 HD Mouse Striatum. Int. J. Mol. Sci. 2023, 24, 11001. https://doi.org/10.3390/ijms241311001
Pinchaud K, Masson C, Dayre B, Mounier C, Gilles J-F, Vanhoutte P, Caboche J, Betuing S. Cell-Type Specific Regulation of Cholesterogenesis by CYP46A1 Re-Expression in zQ175 HD Mouse Striatum. International Journal of Molecular Sciences. 2023; 24(13):11001. https://doi.org/10.3390/ijms241311001
Chicago/Turabian StylePinchaud, Katleen, Chloé Masson, Baptiste Dayre, Coline Mounier, Jean-François Gilles, Peter Vanhoutte, Jocelyne Caboche, and Sandrine Betuing. 2023. "Cell-Type Specific Regulation of Cholesterogenesis by CYP46A1 Re-Expression in zQ175 HD Mouse Striatum" International Journal of Molecular Sciences 24, no. 13: 11001. https://doi.org/10.3390/ijms241311001
APA StylePinchaud, K., Masson, C., Dayre, B., Mounier, C., Gilles, J. -F., Vanhoutte, P., Caboche, J., & Betuing, S. (2023). Cell-Type Specific Regulation of Cholesterogenesis by CYP46A1 Re-Expression in zQ175 HD Mouse Striatum. International Journal of Molecular Sciences, 24(13), 11001. https://doi.org/10.3390/ijms241311001