1. Introduction
Triplin is a channel-forming structure found in
Escherichia coli. It forms a set of three pores whose effective size and conductance resemble OmpC, whereas its weak ion selectivity resembles OmpF [
1]. Triplin differs dramatically for all porins described to date (e.g., [
2,
3,
4,
5]) in that it displays steep voltage dependence [
6] comparable to that of the voltage-gated channels responsible for the electrical excitability of the mammalian nervous system. A unique property of Triplin that distinguishes it from all membrane channels and pores described to date is the remarkable interpore cooperative behavior in that pore 1 must close first, its closure then allows pore 2 to close and, in turn, pore 2 closure allows pore 3 to close [
6]. This is illustrated in
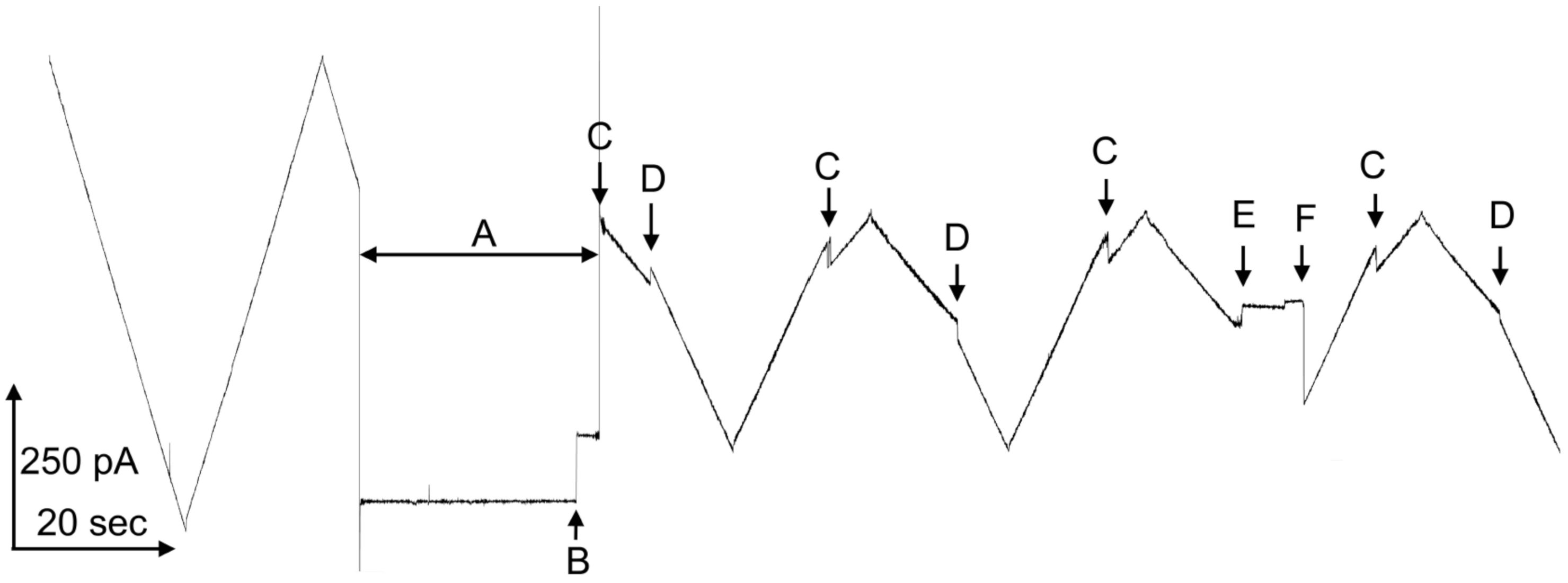
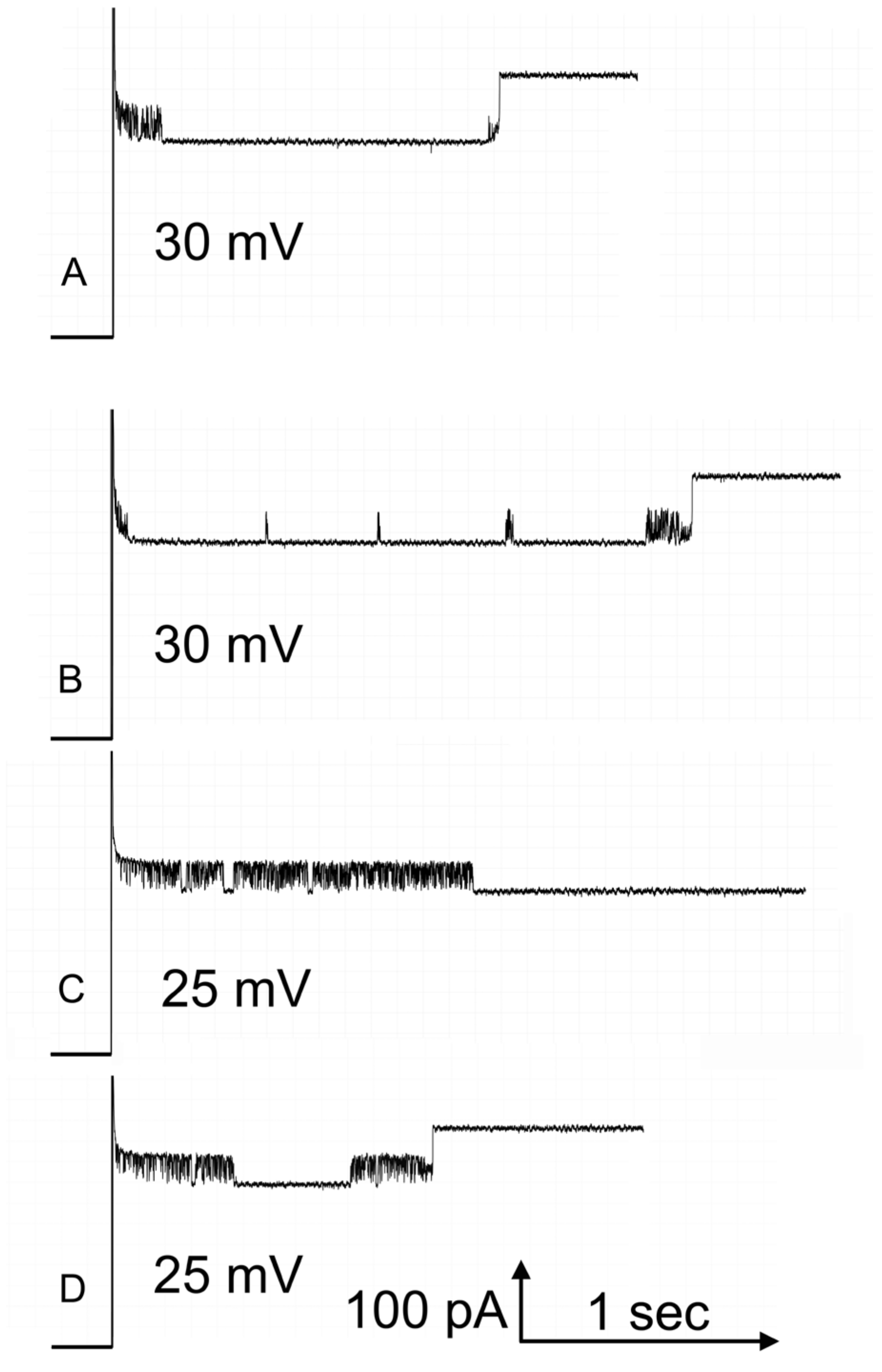
Figure 1. At left, a single Triplin shows no voltage-dependent gating. The application of a high positive potential during time interval “A”, closes pore 1 (at point “B”) and it remains closed. That closure allows pore 2 to close at negative voltages (at point “C”). The reopening of pore 2 (at point “D”) prevents the closing of pore 3 (at point “E”). A delayed reopening of pore 2 allows pore 3 to close at positive voltages.
Insights into the molecular basis for this complex were obtained by probing the nature and dynamics of the voltage sensors of Triplin [
1]. This led the authors to propose the following model.
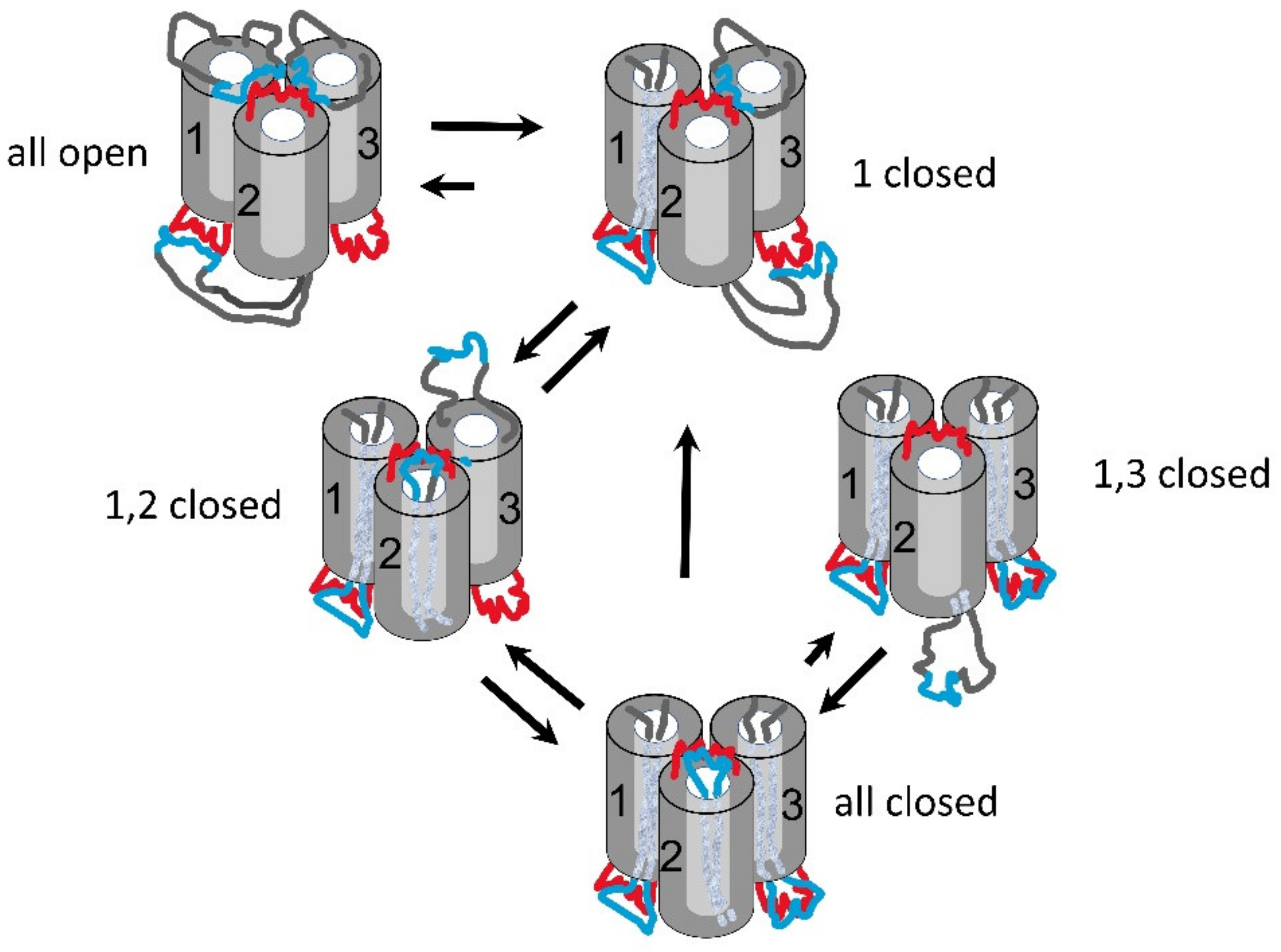
Triplin is proposed to form three beta barrel pores, similar to those of other three-pore porins, but one pore-forming structure is oriented opposite to the others [
1]. In the model (
Figure 2), each pore-forming subunit has a positively charged voltage sensor (blue region) located in external loops on one end of the pore. Pore closure is proposed to result from the entry of the sensor into the pore, thus blocking ion flow. When the sensor is in the pore, it is proposed to interact with a negative domain located at the other end of the pore, with the negative domain also being responsible for the pore’s weak cation selectivity and rectification [
1]. No evidence was presented to support this closing mechanism, and this paper addresses that issue.
3. Discussion
Triplin is a three-pore complex, each pore possessing a positively charged voltage sensor consisting of 14 net charges [
1,
6]. Each sensor domain translocates across the membrane in a steeply voltage-dependent manner, and this translocation is coupled with the closure of each pore [
1]. The proposed molecular mechanism by which sensor translocation results in pore closure is for the sensor to enter the pore and obstruct the flow of ions [
1]. With an estimated pore diameter of 0.9 nm [
1], there is enough space for surface loops to enter the pore and obstruct it. Indeed, it is not uncommon for porin to normally have a surface loop inserted into the pore, reducing its effective diameter (e.g., loop 3 in OmpF [
5]). Other mechanisms are possible [
7], but the experimental evidence presented here supports the proposed mechanism.
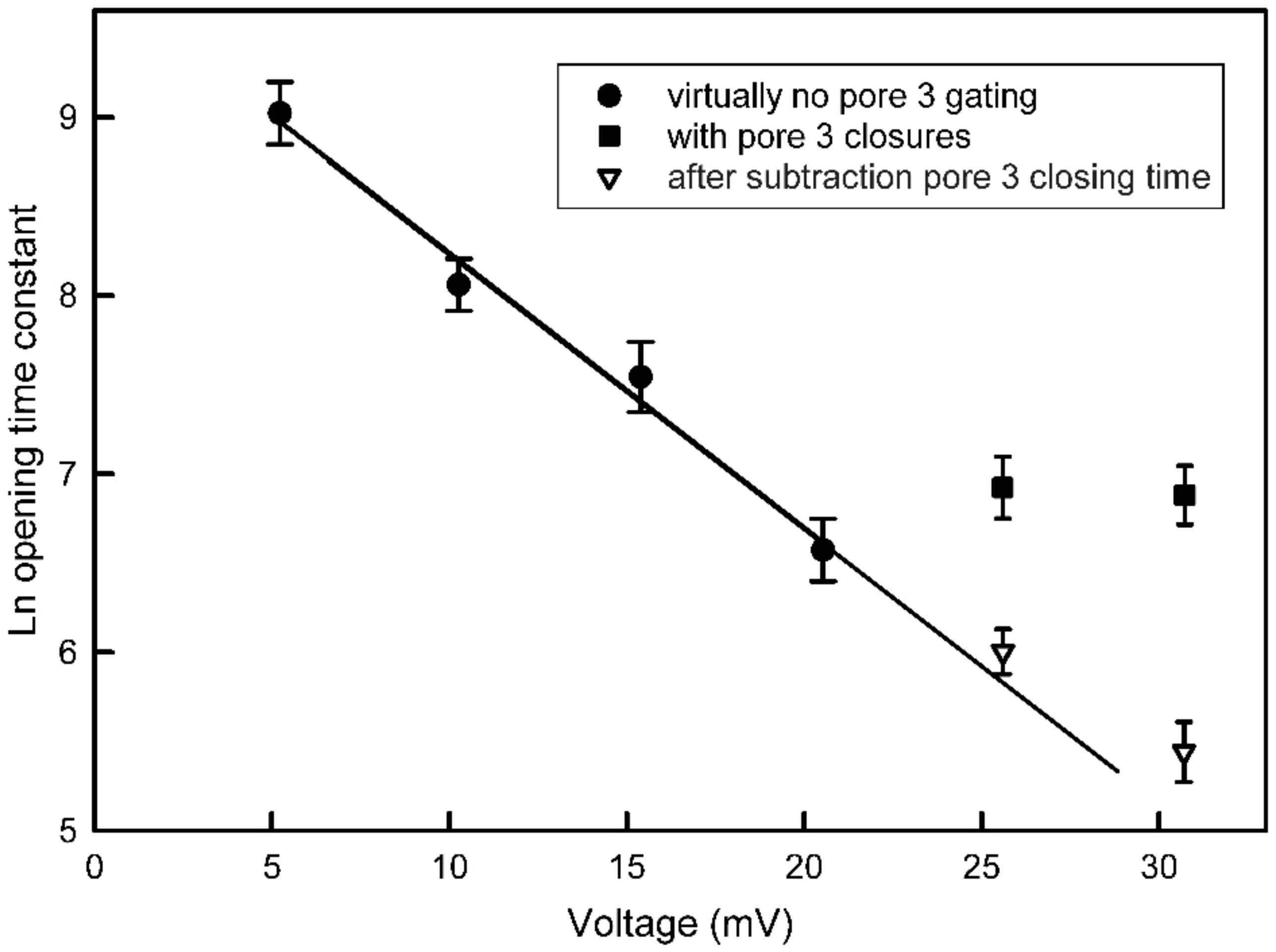
Kinetic measurements can not only provide insight into the molecular mechanism by which voltage-gating takes place but, more importantly, can provide compelling evidence against the proposed mechanism. The kinetics of a well-studied beta barrel channel former, VDAC, provide useful contrasting kinetic properties to Triplin. For Triplin, the rates of pore closure for pores 2 and 3 are fast whereas the rates of reopening are slow [
6]. For VDAC, pore closure is slow (τ ≈ 400 s) and voltage dependent whereas reopening is fast (τ ≈ 2.5 ms) and voltage independent [
8]. The voltage-dependent effective closure of VDAC (actually a reduction in pore size and selectivity inversion that results in no flux of ATP) is achieved by the translocation of portions of the beta barrel being moved to the membrane surface [
8]. The slow closure rate of VDAC can be readily understood by the need to break at least two sets of hydrogen bonds that anchor the mobile domain to the rest of the transmembrane beta barrel before they can translocate to the membrane surface. Reopening involves moving this mobile region from the membrane surface and reinserting it into the beta barrel. Half as many hydrogen bonds in the beta barrel would need to be broken for the reinsertion of the mobile domain to take place.
For Triplin, the proposed mechanism would reverse the energetics. Closure of the pores in Triplin is proposed to result from the entry of a surface loop-like domain into the pore led by the positively charged region (blue;
Figure 11). Once in the pore, this mobile sensor domain is proposed to bind to a negatively charged domain (red) on the other end of the pore. The breaking of that electrostatic interaction then would be responsible for the slow reopening kinetics.
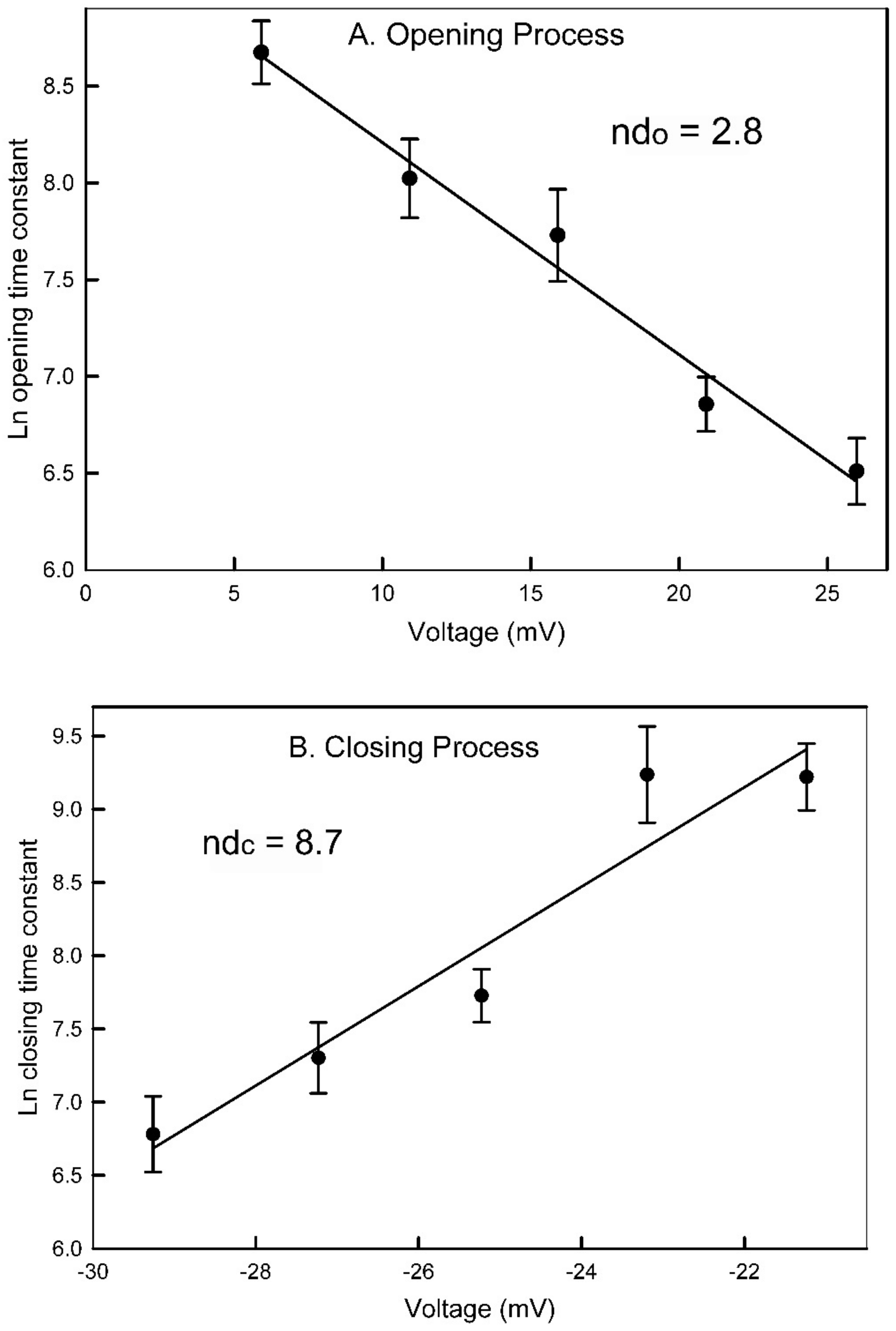
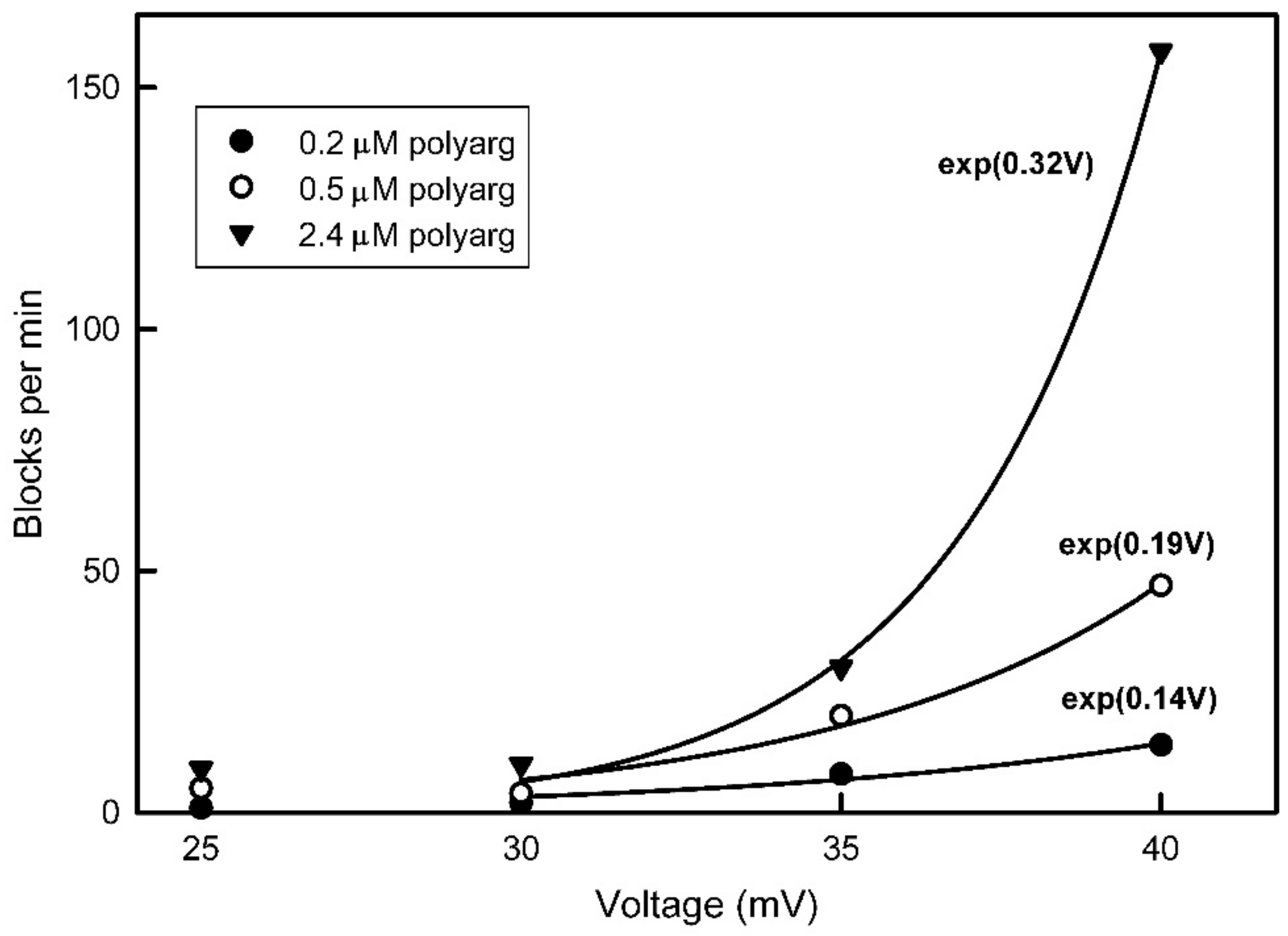
The measured voltage dependence of pore-closing and opening time constants provides independent support for the pore-blocking closure mechanism. The pore-closing process is four times more voltage dependent than the opening process indicating that the energy barrier for the process is deep within the pore. Therefore, the closing process would require the sensor to move through most of the electric field before reaching the peak of the energy barrier, and the reverse would be the case for the reopening process. Focusing on the gating model, what aspect of the structure might correspond to the energy barrier? All surfaces, especially charged surfaces, have a layer of bound water of hydration. This is known to be a critical factor in determining the binding energy between two surfaces and thus critical to determining the binding selectivity as demonstrated by George Eisenman [
9]. Thus, the location of the energy barrier could very well be at a point where the sensor approaches the complementary binding domain. If the proposed negatively charged region at the opposite end of the pore was part of that complementary surface, it would explain (correlate with) the results of the kinetic studies.
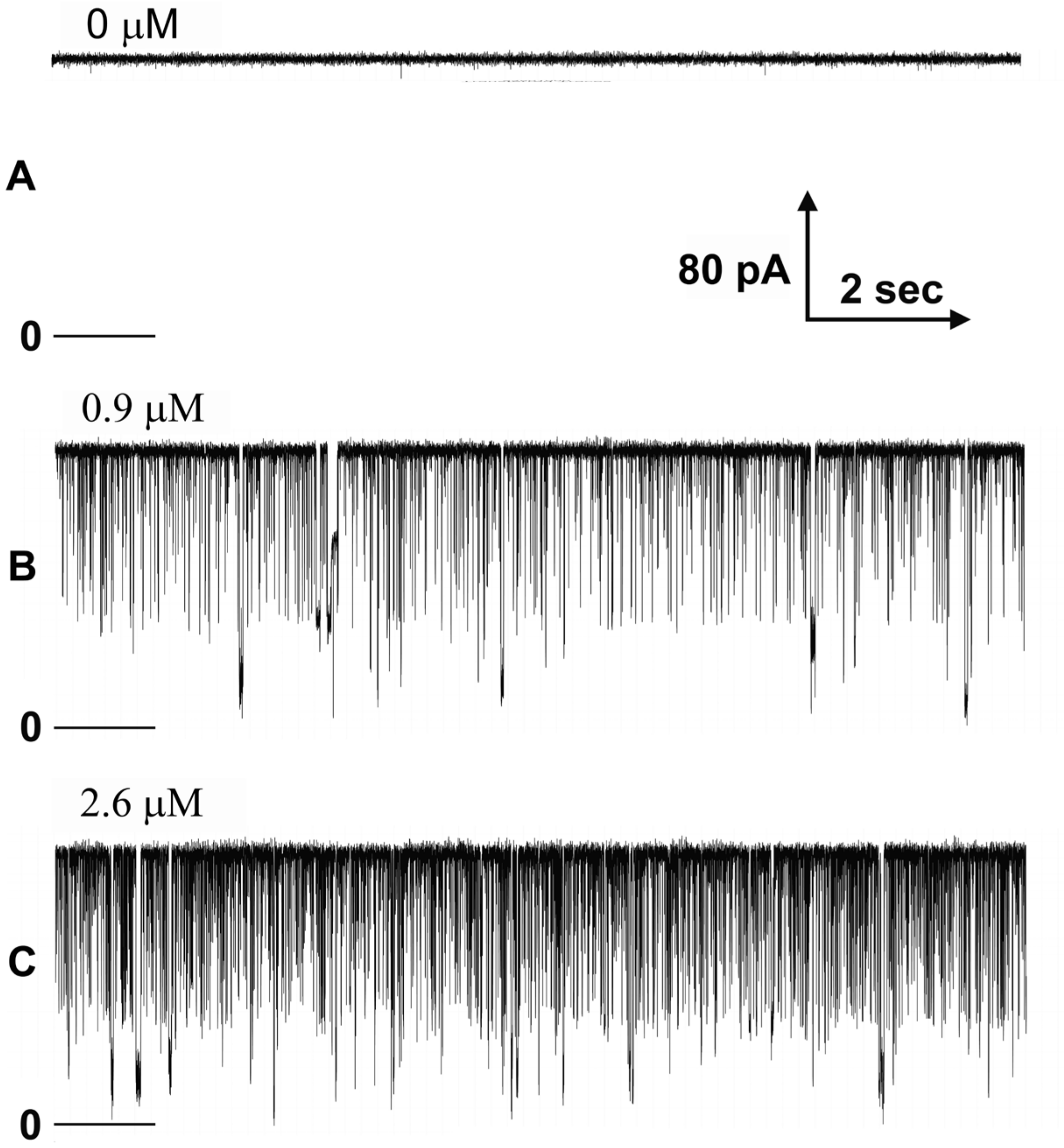
The existence of a binding region complementary to the sensor domain is strongly supported by the specificity of the polyarginine blocking. The differential ability to block the pore by polyarginine as opposed to polylysine is greater than 500-fold since no detectable blocking by polylysine was observed. This specificity of the blocking of polyarginine over polylysine is not related to any propensity for secondary structure formation as both have an extended conformation under the experimental conditions used [
10,
11]. Thus, the very strong preference for polyarginine indicates a specific interacting surface in Triplin that goes beyond just an electrostatic interaction.
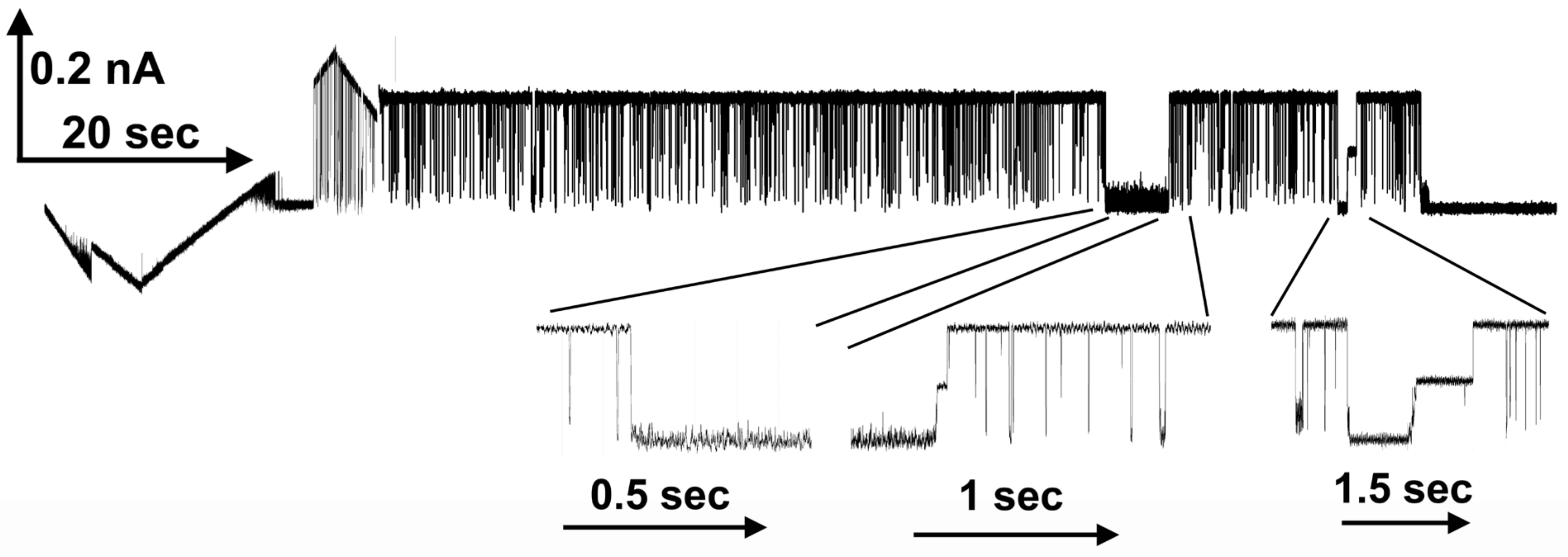
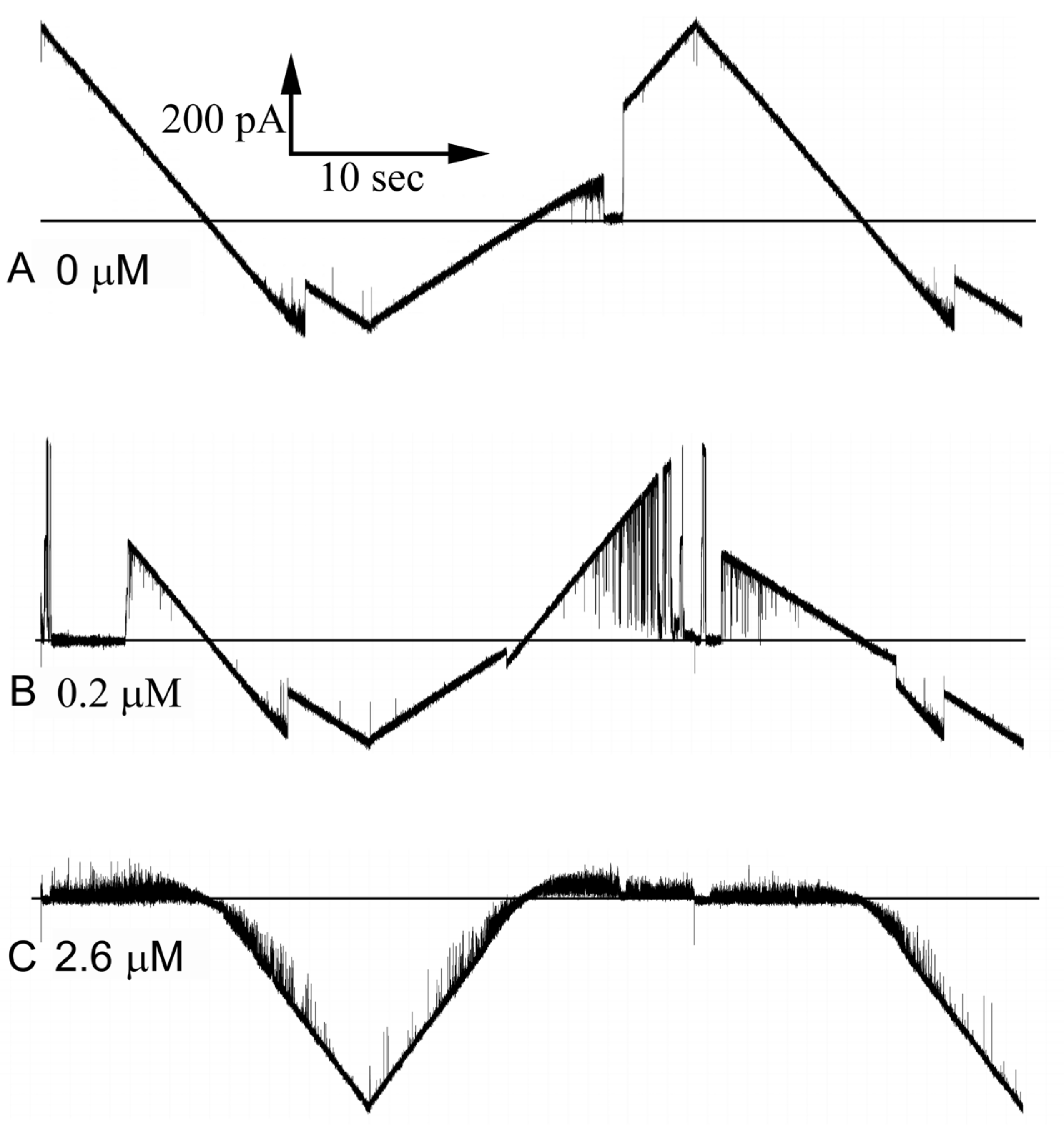
Is the blockage of ion flow through the Triplin pores really mimicking the blockage produced by the sensor domain? Whereas the long-lived blockages must be physical blockages, the transient ones might be the result of polyarginine translocating through the pore as it moves from one side of the membrane to the other. However, calculations indicate that an extended polyarginine chain driven through the pore with an electric field should travel at a rate that is 104 times faster than that observed for the typical duration of the fast current blocking (flickering) events. Thus, it is more likely that the flickering is a result of transient blocking of the pore. This conclusion is supported by the observation that frequently all the conductance is blocked simultaneously, and thus, all open pores of a Triplin are blocked simultaneously. That would be a very unlikely event if the conductance drops were due to polyarginine translocating through the pores.
An observation that would seem to indicate that the polyarginine is translocating through the pores rather than blocking is the finding that the flickering rate increases with an increase in the aqueous concentration of polyarginine. However, the interaction between polyarginine and Triplin may be quite labile, undergoing a dynamic equilibrium with dissolved polymer. Thus, increasing the concentration of polyarginine would increase the blocking frequency.
The properties of the polyarginine block are consistent with the model of the gating mechanism. The long, highly charged polypeptide chain should easily flow through a simple cylindrical pore, especially when driven by an electric field. Published work indicates that polyarginine should have a primarily extended conformation under the conditions of neutral pH used in these experiments [
10]. Thus, the observation of blockage indicates some stabilizing interaction within the pore and/or interference to flow by the proposed negatively charged domain at one end of each pore.
4. Materials and Methods
4.1. Sources of Materials Used
All chemicals used were reagent grade. The phospholipids were obtained from Avanti Polar Lipids (Alabaster, AL, USA). Cholesterol, polylysine, polyarginine, and spermine were purchased from Sigma (St. Louis, MO, USA).
4.2. Electrophysiological Recordings
All experiments were performed on Triplin reconstituted into planar phospholipid membranes made from monolayers as described previously [
12,
13]. In brief, the membrane was formed across a 0.1 mm hole in a thin polyvinylidene chloride partition separating two aqueous compartments containing 5 mL of 1.0 M KCl, 1 mM MgCl
2, buffered with 10 mM HEPES, pH 7.8. The monolayers were formed from a solution of 0.5% (
w/
v) diphytanoylphosphatidylcholine, 0.5% (
w/
v) polar extract of soybean phospholipids, and 0.05% (
w/
v) cholesterol in hexane. The hexane was allowed to evaporate prior to membrane formation, and thus, no solvents were present in the membrane. This membrane is identical to a natural cell membrane but lacks proteins or carbohydrates. Samples containing Triplin were generated as previously described [
6], i.e., flash-frozen in 0.1 mL aliquots and stored at −80 °C. After thawing, β-octyl-glucoside was added to a final concentration of 1% (
w/
v) and kept on ice during the experiment. Typically, 10 µL of the sample was dispersed into one aqueous compartment (designated “
cis”), and, with time, Triplin would insert into the membrane. The membrane voltage was clamped, and the current recorded using Clampex 10.3 software. Calomel electrodes were used to interface the solution with the electronics. Voltages was applied to the
cis side, the
trans held at virtual ground. The signal was low-pass filtered at 500 Hz. The sample containing Triplin was always added to the
cis compartment. All measurements were performed at room temperature (23 to 24 °C)
4.3. Kinetic Measurements
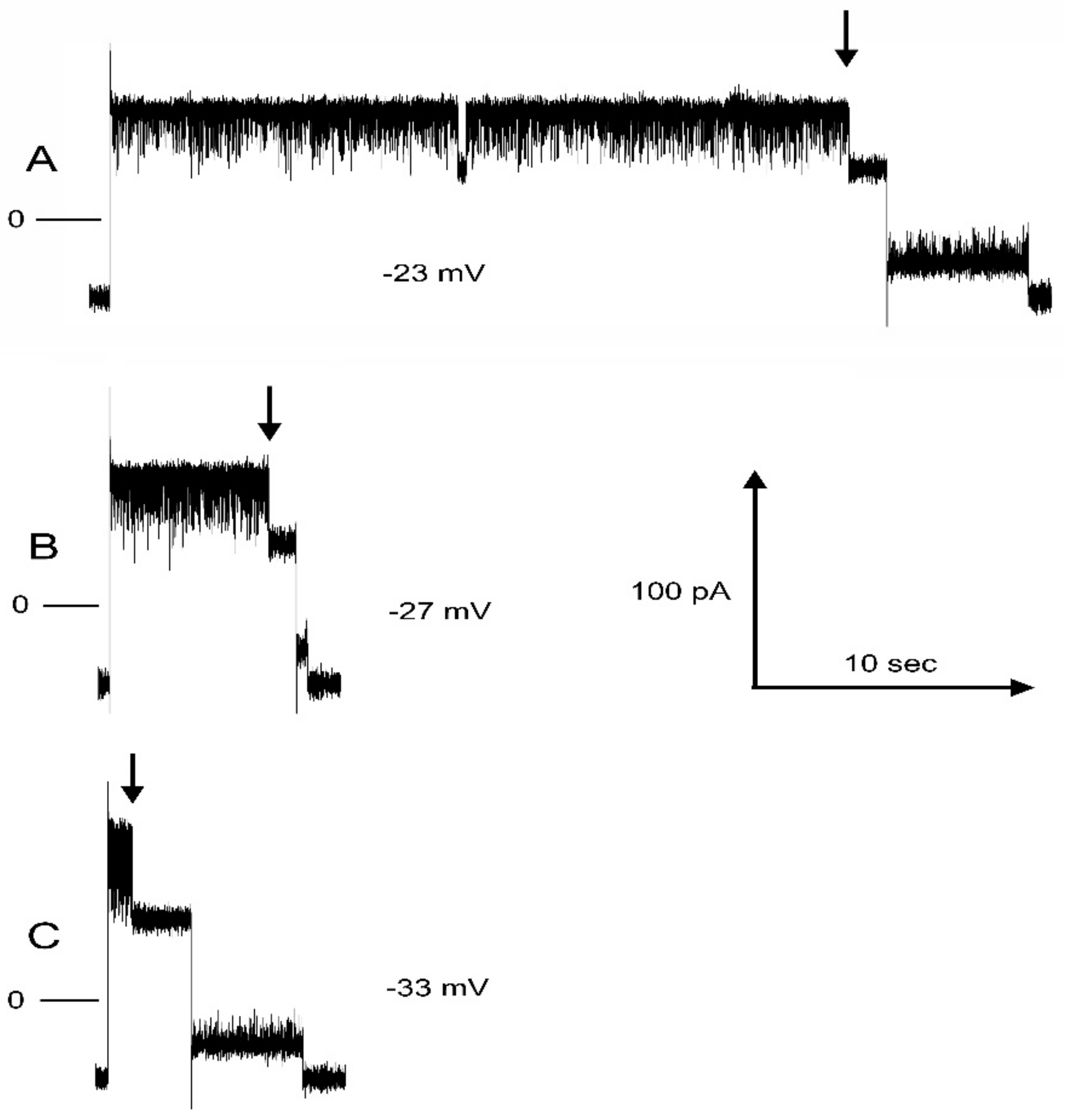
Kinetic measurements were only made on pore 2 because the conditions necessary for making those measurements for pore 3 often resulted in the spontaneous opening of pore 2 and thus interfering with the collection of sufficient data to achieve valid measurements. Recordings were made on membranes containing a single Triplin so that the meaning of any conductance change was clearly defined. For the closing kinetics, pore 2 was held in the open state at 10 mV and then switched to a negative voltage until pore 2 closed, and thus, the time required for closure was recorded. This process was repeated 15 to 20 times, depending on the experiment, for negative voltages ranging from −20 to −30 mV. For the opening process, pore 2 was closed by applying −40 mV, and then, the voltage was switched to a positive value until the pore reopened. Again, the process was repeated 15 to 20 times, depending on the experiment, for positive voltages ranging from 5 to 40 mV. At the higher voltages, pore 3 would frequently close and reopen during the waiting period for pore 2 to reopen.
Theoretical Basis for Estimating the Fraction of the Electric Field Traversed Prior to Reaching the Peak of the Energy Barrier
Using Eyring Rate Theory, one can obtain an estimate of the fraction of the transmembrane electric field that the sensor needed to traverse prior to reaching the peak of the energy barrier.
where the forward reaction rate constant is
ko and the reverse is
kcFrom Eyring rate theory:
where
h is Plank’s constant,
kB is Boltzmann’s constant,
n is the number of charges on the voltage sensor,
do is the fraction of the electric field that the sensor must traverse to move from the closed state to the peak of the energy barrier whereas
dc is from the open state to the barrier peak.
V is the transmembrane voltage, and
Vo is the voltage at which half the pores are open and half are closed.
R,
T, and
F have their usual meanings.
After log transforming and simplifying:
and



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










