

Adaptation of the Th-MYCN Mouse Model of Neuroblastoma for Evaluation of Disseminated Disease

 ,
,
Abstract
:1. Introduction
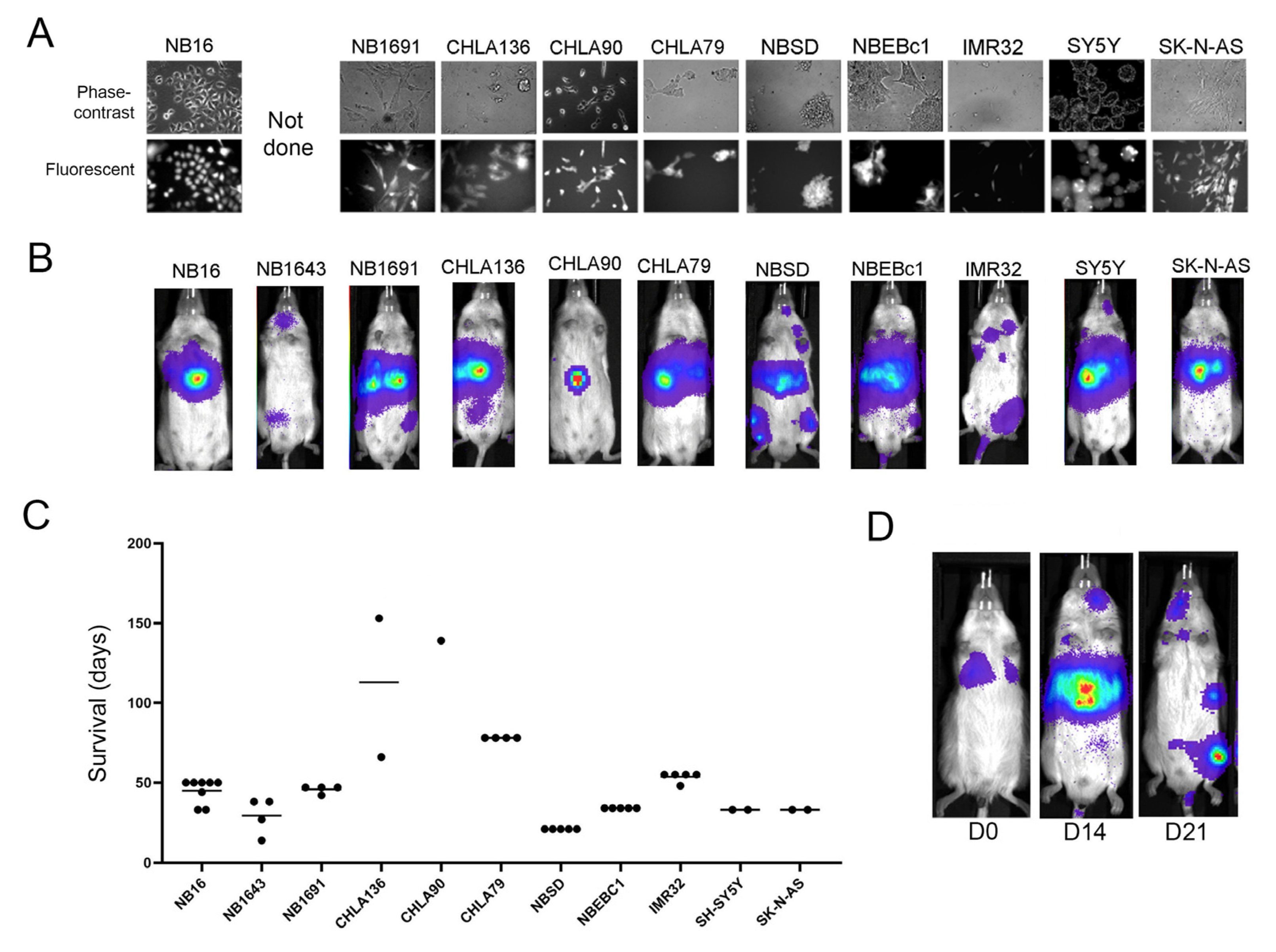
2. Results
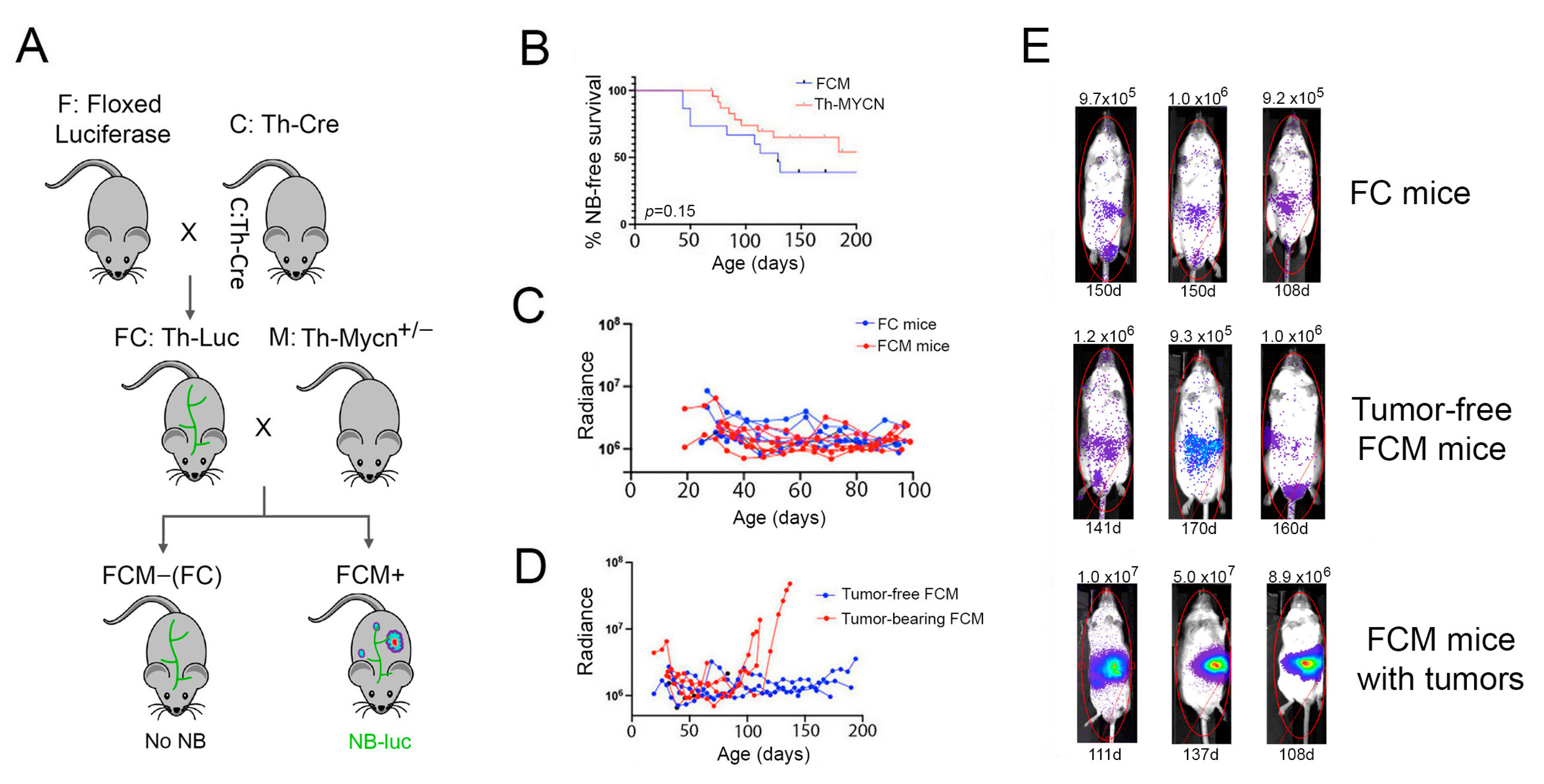
2.1. Bioluminescence-Based Monitoring of Neuroblastoma Progression in Th-MYCN Mice
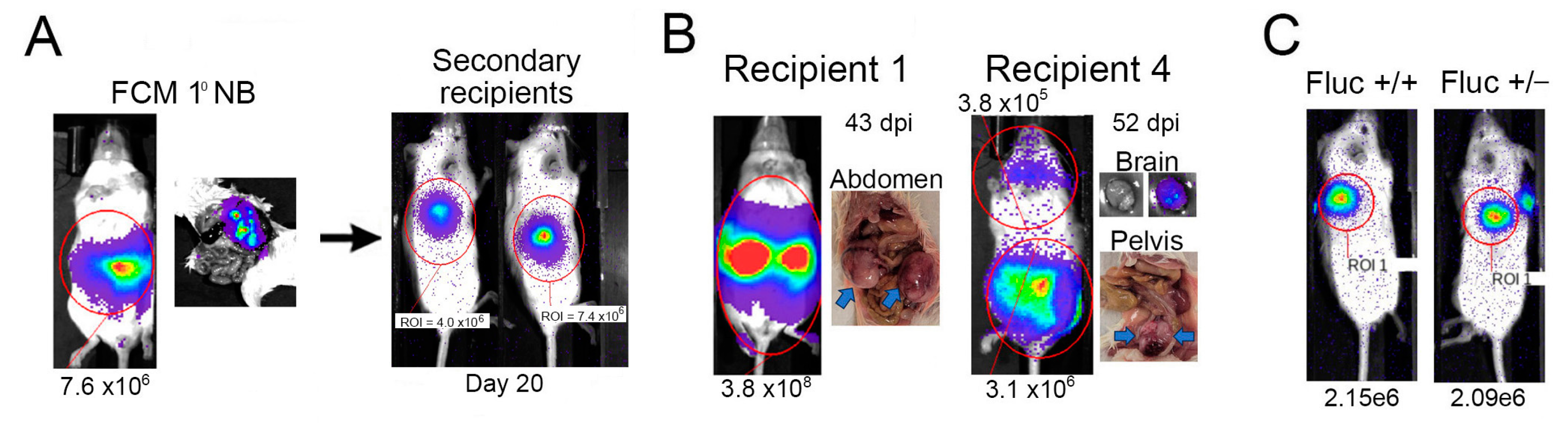
2.2. Tagged Th-MYCN-Derived Tumors Are Serially Transplantable
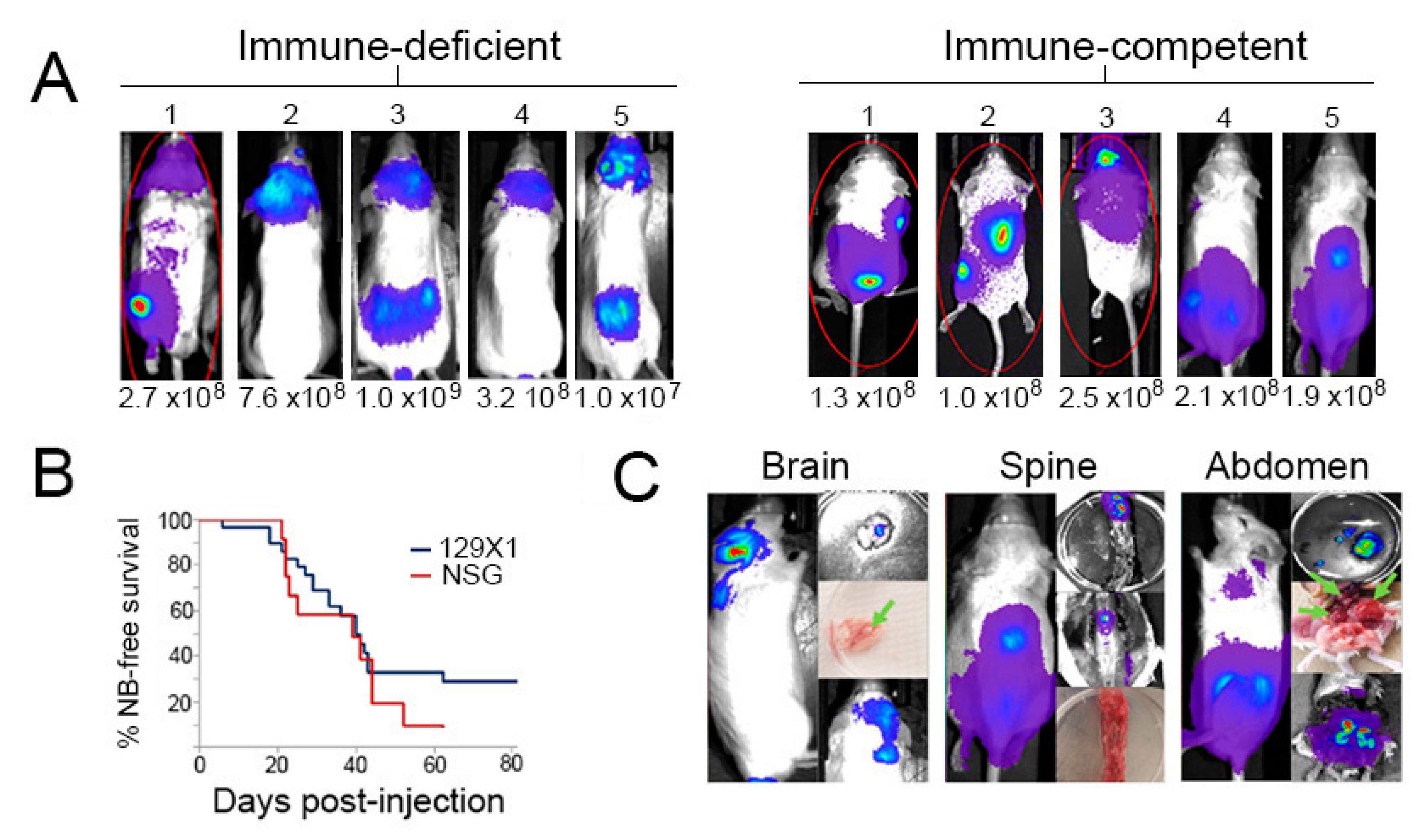
2.3. Transplanted Th-MYCN Tumor Cells Establish Broadly Disseminated Tumors
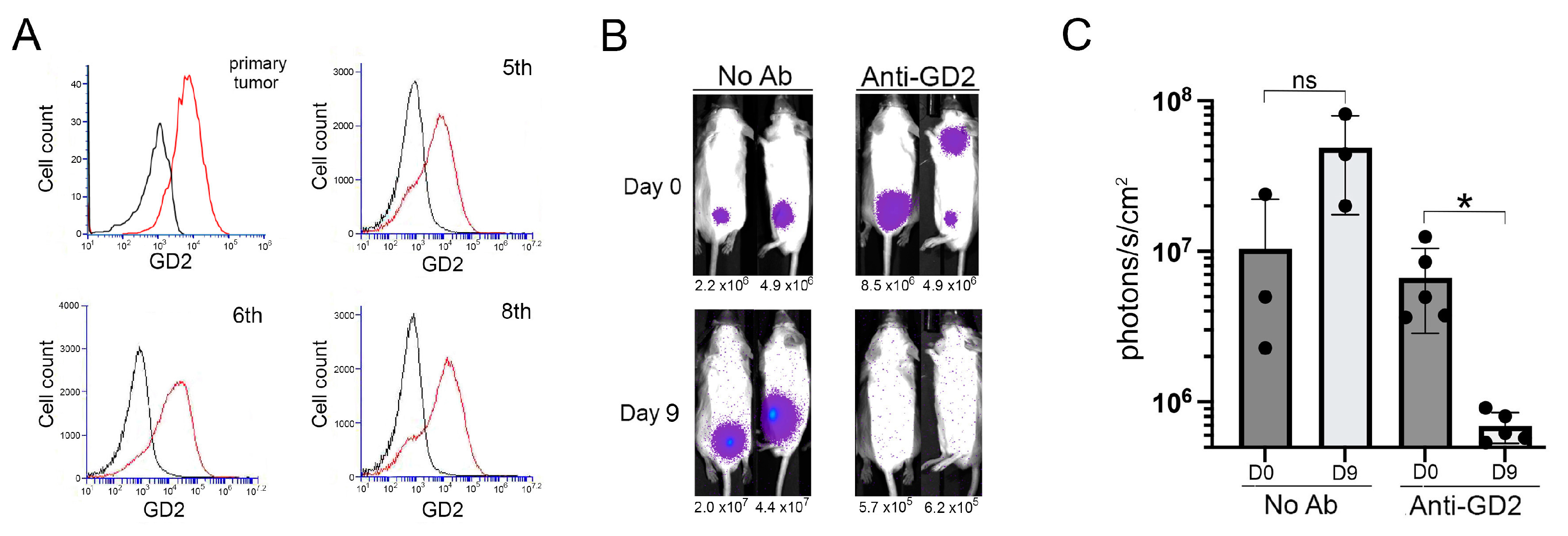
2.4. GD2-Targeted Depletion of Transplanted Th-MYCN Neuroblastoma
2.5. Localization and Growth in Human NB Cell Lines in Xenografts
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Mice
4.3. Isolation of Primary Th-MYCN-Derived NB Cells
4.4. Luciferase-Tagging Neuroblastoma Cells
4.5. Longitudinal Monitoring of NB Growth
4.6. Statistical Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, P.; Qi, F.; Bian, L.; Xu, Y.; Zhou, J.; Hu, J.; Ren, L.; Li, M.; Tang, W. Comparison of Incidence and Outcomes of Neuroblastoma in Children, Adolescents, and Adults in the United States: A Surveillance, Epidemiology, and End Results (SEER) Program Population Study. Med. Sci. Monit. 2020, 26, e927218-1–e927218-13. [Google Scholar] [CrossRef] [PubMed]
- Brodeur, G.M. Neuroblastoma: Biological insights into a clinical enigma. Nat. Rev. Cancer 2003, 3, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.S.; Naranjo, A.; Zhang, F.F.; Cohn, S.L.; London, W.B.; Gastier-Foster, J.M.; Ramirez, N.C.; Pfau, R.; Reshmi, S.; Wagner, E.; et al. Revised Neuroblastoma Risk Classification System: A Report From the Children’s Oncology Group. J. Clin. Oncol. 2021, 39, 3229–3241. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, L.; Erminio, G.; Makin, G.; Pearson, A.D.J.; Brock, P.; Valteau-Couanet, D.; Castel, V.; Pasquet, M.; Laureys, G.; Thomas, C.; et al. Topotecan-Vincristine-Doxorubicin in Stage 4 High-Risk Neuroblastoma Patients Failing to Achieve a Complete Metastatic Response to Rapid COJEC: A SIOPEN Study. Cancer Res. Treat. 2018, 50, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, V.; Foster, J. High-Risk Neuroblastoma Treatment Review. Children 2018, 5, 114. [Google Scholar] [CrossRef] [Green Version]
- Bayeva, N.; Coll, E.; Piskareva, O. Differentiating Neuroblastoma: A Systematic Review of the Retinoic Acid, Its Derivatives, and Synergistic Interactions. J. Pers. Med. 2021, 11, 211. [Google Scholar] [CrossRef]
- Chan, G.C.F.; Chan, C.M. Anti-GD2 Directed Immunotherapy for High-Risk and Metastatic Neuroblastoma. Biomolecules 2022, 12, 358. [Google Scholar] [CrossRef]
- DuBois, S.G.; Macy, M.E.; Henderson, T.O. High-Risk and Relapsed Neuroblastoma: Toward More Cures and Better Outcomes. In American Society of Clinical Oncology Educational Book; American Society of Clinical Oncology: Alexandria, VA, USA, 2022; Volume 42, pp. 1–13. [Google Scholar]
- Applebaum, M.A.; Desai, A.V.; Bender, J.L.G.; Cohn, S.L. Emerging and investigational therapies for neuroblastoma. Expert Opin. Orphan Drugs 2017, 5, 355. [Google Scholar] [CrossRef] [Green Version]
- DuBois, S.G.; Kalika, Y.; Lukens, J.N.; Brodeur, G.M.; Seeger, R.C.; Atkinson, J.B.; Haase, G.M.; Black, C.T.; Perez, C.; Shimada, H.; et al. Metastatic sites in stage IV and IVS neuroblastoma correlate with age, tumor biology, and survival. J. Pediatr. Hematol. Oncol. 1999, 21, 181–189. [Google Scholar] [CrossRef]
- Liu, S.; Yin, W.; Lin, Y.; Huang, S.; Xue, S.; Sun, G.; Wang, C. Metastasis pattern and prognosis in children with neuroblastoma. World J. Surg. Oncol. 2023, 21, 130. [Google Scholar] [CrossRef] [PubMed]
- DuBois, S.G.; London, W.B.; Zhang, Y.; Matthay, K.K.; Monclair, T.; Ambros, P.F.; Cohn, S.L.; Pearson, A.; Diller, L. Lung Metastases in Neuroblastoma at Initial Diagnosis: A Report from the International Neuroblastoma Risk Group (INRG) Project. Pediatr. Blood Cancer 2008, 51, 589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, K.; Kushner, B.H.; Modak, S.; Pandit-Taskar, N.; Tomlinson, U.; Wolden, S.L.; Zanzonico, P.; John, H.L.; Haque, S.; Souweidane, M.M.; et al. A curative approach to central nervous system metastases of neuroblastoma. J. Clin. Oncol. 2017, 35, 10545. [Google Scholar] [CrossRef]
- Mastronuzzi, A.; Colafati, G.S.; Carai, A.; D’egidio, M.; Fabozzi, F.; Del Bufalo, F.; Villani, M.F.; Del Baldo, G.; Vennarini, S.; Canino, C.; et al. Central Nervous System Metastasis in Neuroblastoma: From Three Decades Clinical Experience to New Considerations in the Immunotherapy Era. Cancers 2022, 14, 6249. [Google Scholar] [CrossRef] [PubMed]
- Esiashvili, N.; Anderson, C.; Katzenstein, H.M. Neuroblastoma. Curr. Probl. Cancer 2009, 33, 333–360. [Google Scholar] [CrossRef]
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Prim. 2016, 2, 16078. [Google Scholar] [CrossRef]
- Kamili, A.; Atkinson, C.; Trahair, T.N.; Fletcher, J.I. Mouse models of high-risk neuroblastoma. Cancer Metastasis Rev. 2020, 39, 261–274. [Google Scholar] [CrossRef]
- Ornell, K.J.; Coburn, J.M. Developing preclinical models of neuroblastoma: Driving therapeutic testing. BMC Biomed. Eng. 2019, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Braekeveldt, N.; Bexell, D. Patient-derived xenografts as preclinical neuroblastoma models. Cell Tissue Res. 2018, 372, 233. [Google Scholar] [CrossRef]
- Kroesen, M.; Brok, I.C.; Reijnen, D.; van Hout-Kuijer, M.A.; Zeelenberg, I.S.; Brok, M.H.D.; Hoogerbrugge, P.M.; Adema, G.J. Intra-adrenal murine TH-MYCN neuroblastoma tumors grow more aggressive and exhibit a distinct tumor microenvironment relative to their subcutaneous equivalents. Cancer Immunol. Immunother. 2015, 64, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Weiss, W.; Aldape, K.; Mohapatra, G.; Feuerstein, B.; Bishop, J. Targeted expression of MYCN causes neuroblastoma in transgenic mice. EMBO J. 1997, 16, 2985–2995. [Google Scholar] [CrossRef]
- Rasmuson, A.; Segerström, L.; Nethander, M.; Finnman, J.; Elfman, L.H.M.; Javanmardi, N.; Nilsson, S.; Johnsen, J.I.; Martinsson, T.; Kogner, P. Tumor Development, Growth Characteristics and Spectrum of Genetic Aberrations in the TH-MYCN Mouse Model of Neuroblastoma. PLoS ONE 2012, 7, e51297. [Google Scholar] [CrossRef] [PubMed]
- Hackett, C.S.; Hodgson, J.G.; Law, M.E.; Fridlyand, J.; Osoegawa, K.; De Jong, P.J.; Nowak, N.J.; Pinkel, D.; Albertson, D.G.; Jain, A.; et al. Genome-wide array CGH analysis of murine neuroblastoma reveals distinct genomic aberrations which parallel those in human tumors. Cancer Res. 2003, 63, 5266–5273. [Google Scholar] [PubMed]
- Weiss, W.; Godfrey, T.; Francisco, C.; Bishop, J. Genome-wide screen for allelic imbalance in a mouse model for neuroblastoma. Cancer Res. 2000, 60, 2483–2487. [Google Scholar] [PubMed]
- Teitz, T.; Inoue, M.; Valentine, M.B.; Zhu, K.; Rehg, J.E.; Zhao, W.; Finkelstein, D.; Wang, Y.-D.; Johnson, M.D.; Calabrese, C.; et al. Th-MYCN mice with caspase-8 deficiency develop advanced neuroblastoma with bone marrow metastasis. Cancer Res. 2013, 73, 4086–4097. [Google Scholar] [CrossRef] [Green Version]
- Teitz, T.; Stanke, J.J.; Federico, S.; Bradley, C.L.; Brennan, R.; Zhang, J.; Johnson, M.D.; Sedlacik, J.; Inoue, M.; Zhang, Z.M.; et al. Preclinical Models for Neuroblastoma: Establishing a Baseline for Treatment. PLoS ONE 2011, 6, e19133. [Google Scholar] [CrossRef] [PubMed]
- Dodd, K.W.; Burns, T.C.; Weisner, S.M.; Kudishevich, E.; Schomberg, D.T.; Jung, B.-W.; Kim, J.-E.; Ohlfest, J.R.; Low, W.C. Transgenic Mice Expressing Luciferase Under a 4.5 kb Tyrosine Hydroxylase Promoter. Cureus 2011, 3, e34. [Google Scholar] [CrossRef] [Green Version]
- Hansford, L.M.; Thomas, W.D.; Keating, J.M.; Burkhart, C.A.; Peaston, A.E.; Norris, M.D.; Haber, M.; Armati, P.J.; Weiss, W.A.; Marshall, G.M. Mechanisms of embryonal tumor initiation: Distinct roles for MycN expression and MYCN amplification. Proc. Natl. Acad. Sci. USA 2004, 101, 12664–12669. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Cuadrado, L.; Tracey, N.; Ma, R.; Qian, B.; Brunton, V.G. Mouse models of metastasis: Progress and prospects. Dis. Model. Mech. 2017, 10, 1061. [Google Scholar] [CrossRef] [Green Version]
- Barrett, D.M.; Seif, A.E.; Carpenito, C.; Teachey, D.T.; Fish, J.D.; June, C.H.; Grupp, S.A.; Reid, G.S.D. Noninvasive bioluminescent imaging of primary patient acute lymphoblastic leukemia: A strategy for preclinical modeling. Blood 2011, 118, e112–e117. [Google Scholar] [CrossRef] [Green Version]
- Voeller, J.; Erbe, A.K.; Slowinski, J.; Rasmussen, K.; Carlson, P.M.; Hoefges, A.; VandenHeuvel, S.; Stuckwisch, A.; Wang, X.; Gillies, S.D.; et al. Combined innate and adaptive immunotherapy overcomes resistance of immunologically cold syngeneic murine neuroblastoma to checkpoint inhibition. J. Immunother. Cancer 2019, 7, 344. [Google Scholar] [CrossRef]
- McNerney, K.; Karageorgos, S.; Ferry, G.; Wolpaw, A.; Burudpakdee, C.; Khurana, P.; Toland, C.; Vemu, R.; Vu, A.; Hogarty; et al. TH-MYCN tumors, but not tumor-derived cell lines, are adrenergic lineage, GD2+, and responsive to anti-GD2 antibody therapy. Oncoimmunology 2022, 11, 2075204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yeger, H.; Das, B.; Irwinz, M.S.; Baruchel, S. Tissue Microenvironment Modulates CXCR4 Expression and Tumor Metastasis in Neuroblastoma. Neoplasia 2007, 9, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, E.R.; Lanati, S.; Wareham, C.; Easton, A.; Dunn, S.N.; Inzhelevskaya, T.; Sadler, F.M.; James, S.; Ashton-Key, M.; Cragg, M.S.; et al. Immune characterization of pre-clinical murine models of neuroblastoma. Sci. Rep. 2020, 10, 16695. [Google Scholar] [CrossRef]
- Engler, S.; Thiel, C.; Förster, K.; David, K.; Bredehorst, R.; Juhl, H. A Novel Metastatic Animal Model Reflecting the Clinical Appearance of Human Neuroblastoma: Growth Arrest of Orthotopic Tumors by Natural, Cytotoxic Human Immunoglobulin M Antibodies 1. Cancer Res. 2001, 61, 2968–2973. [Google Scholar]
- Seong, B.K.A.; Fathers, K.E.; Hallett, R.M.; Yung, C.K.; Stein, L.D.; Mouaaz, S.; Kee, L.; Hawkins, C.E.; Irwin, M.S.; Kaplan, D.R. A metastatic mouse model identifies genes that regulate neuroblastoma metastasis. Cancer Res. 2017, 77, 696–706. [Google Scholar] [CrossRef] [Green Version]
- Jung, J. Human Tumor Xenograft Models for Preclinical Assessment of Anticancer Drug Development. Toxicol. Res. 2014, 30, 1. [Google Scholar] [CrossRef]
- Romiani, A.; Spetz, J.; Shubbar, E.; Lind, D.E.; Hallberg, B.; Palmer, R.H.; Forssell-Aronsson, E. Neuroblastoma xenograft models demonstrate the therapeutic potential of 177Lu-octreotate. BMC Cancer 2021, 21, 1–14. [Google Scholar] [CrossRef]
- Aaltonen, K.; Radke, K.; Adamska, A.; Seger, A.; Mañas, A.; Bexell, D. Patient-derived models: Advanced tools for precision medicine in neuroblastoma. Front. Oncol. 2023, 12, 1085270. [Google Scholar] [CrossRef]
- Sun, M.; Cao, Y.; Okada, R.; Reyes-González, J.M.; Stack, H.G.; Qin, H.; Li, N.; Seibert, C.; Kelly, M.C.; Ruppin, E.; et al. Preclinical optimization of a GPC2-targeting CAR T-cell therapy for neuroblastoma. J. Immunother. Cancer 2023, 11, 5881. [Google Scholar] [CrossRef]
- Heitzeneder, S.; Bosse, K.R.; Zhu, Z.; Zhelev, D.; Majzner, R.G.; Radosevich, M.T.; Dhingra, S.; Sotillo, E.; Buongervino, S.; Pascual-Pasto, G.; et al. GPC2-CAR T cells tuned for low antigen density mediate potent activity against neuroblastoma without toxicity. Cancer Cell 2022, 40, 53–69.e9. [Google Scholar] [CrossRef] [PubMed]
- Harenza, J.L.; Diamond, M.A.; Adams, R.N.; Song, M.M.; Davidson, H.L.; Hart, L.S.; Dent, M.H.; Fortina, P.; Reynolds, C.P.; Maris, J.M. Transcriptomic profiling of 39 commonly-used neuroblastoma cell lines. Sci. Data 2017, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Keshelava, N.; Seeger, R.C.; Reynolds, C.P. Drug resistance in human neuroblastoma cell lines correlates with clinical therapy. Eur. J. Cancer 1997, 33, 2002–2006. [Google Scholar] [CrossRef] [PubMed]
- Krytska, K.; Ryles, H.T.; Sano, R.; Raman, P.; Infarinato, N.R.; Hansel, T.D.; Makena, M.R.; Song, M.M.; Reynolds, C.P.; Mossé, Y.P. Crizotinib synergizes with chemotherapy in preclinical models of neuroblastoma. Clin. Cancer Res. 2016, 22, 948–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos Cogo, S.; Gradowski Farias da Costa do Nascimento, T.; de Almeida Brehm Pinhatti, F.; de Franca Junior, N.; Santos Rodrigues, B.; Cavalli, L.R.; Elifio-Esposito, S. An overview of neuroblastoma cell lineage phenotypes and in vitro models. Exp. Biol. Med. 2020, 245, 1637. [Google Scholar] [CrossRef] [PubMed]
- Dorel, M.; Klinger, B.; Mari, T.; Toedling, J.; Blanc, E.; Messerschmidt, C.; Nadler-Holly, M.; Ziehm, M.; Sieber, A.; Hertwig, F.; et al. Neuroblastoma signalling models unveil combination therapies targeting feedback-mediated resistance. PLOS Comput. Biol. 2021, 17, e1009515. [Google Scholar] [CrossRef]
- Lorentzian, A.; Biegel, J.A.; Ostrow, D.G.; Rolf, N.; Liu, C.-C.; Rassekh, S.R.; Deyell, R.J.; Triche, T.; Schultz, K.R.; Rozmus, J.; et al. Tumor Variant Identification That Accounts for the Unique Molecular Landscape of Pediatric Malignancies. JNCI Cancer Spectr. 2018, 2, pky079. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immune Status | Injected | Engrafted | Survival (Days) | Paraspinal | BM | Cranial | Other |
|---|---|---|---|---|---|---|---|
| Competent | 15 | 11 | 41 ± 28 | 5 | 4 | 9 | 3 |
| Deficient | 9 | 8 | 43 ± 24 | 2 | 4 | 7 | 1 |
| MYCN | ALK | TP53 | Other SNV | |
|---|---|---|---|---|
| CHLA90 | F1245V | TP53 E286K | CIC | |
| NB1643 | Amplified | R1275Q | WT | |
| NBSD | Amplified | F1174L | TP53 C176F | ARID1A, DICER |
| SK-N-AS | WT | CNV 0.5 | NRAS | |
| NB16 | WT | TP53 R248W | PIK3CA, NF1, NRAS | |
| SY5Y | F1174L | WT | SMARCA4 | |
| CHLA79 | Low-amplified (4.9) | WT | WT | |
| CHLA136 | Amplified | WT | WT | |
| NB1691 | Amplified | WT | WT | |
| NBEBc1 | Low-amplified (3.8) | WT | WT | KRAS |
| IMR32 | Amplified | WT | WT |
| Cell Line | Liver | BM | Cranial | Other |
|---|---|---|---|---|
| CHLA90 | 1/6 | |||
| NB1691 | 4/4 | 2/4 | ||
| NBSD | 5/5 | 7/7 | 6/7 | |
| SK-N-AS | 3/3 | |||
| NB16 | 9/9 | 1/9 kidney | ||
| SY5Y | 5/5 | |||
| CHLA79 | 4/4 | 2/4 | ||
| CHLA136 | 2/4 | 1/4 abdominal | ||
| NB1643 | 2/4 | 1/4 | 3/4 abdominal | |
| NBEBc1 | 5/5 | 3/5 | ||
| IMR32 | 5/5 | 5/5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahavi, S.M.; Aletaha, M.; Farrokhi, A.; Lorentzian, A.; Lange, P.F.; Maxwell, C.A.; Lim, C.J.; Reid, G.S.D. Adaptation of the Th-MYCN Mouse Model of Neuroblastoma for Evaluation of Disseminated Disease. Int. J. Mol. Sci. 2023, 24, 12071. https://doi.org/10.3390/ijms241512071
Rahavi SM, Aletaha M, Farrokhi A, Lorentzian A, Lange PF, Maxwell CA, Lim CJ, Reid GSD. Adaptation of the Th-MYCN Mouse Model of Neuroblastoma for Evaluation of Disseminated Disease. International Journal of Molecular Sciences. 2023; 24(15):12071. https://doi.org/10.3390/ijms241512071
Chicago/Turabian StyleRahavi, Seyed M., Maryam Aletaha, Ali Farrokhi, Amanda Lorentzian, Philipp F. Lange, Christopher A. Maxwell, Chinten James Lim, and Gregor S. D. Reid. 2023. "Adaptation of the Th-MYCN Mouse Model of Neuroblastoma for Evaluation of Disseminated Disease" International Journal of Molecular Sciences 24, no. 15: 12071. https://doi.org/10.3390/ijms241512071
APA StyleRahavi, S. M., Aletaha, M., Farrokhi, A., Lorentzian, A., Lange, P. F., Maxwell, C. A., Lim, C. J., & Reid, G. S. D. (2023). Adaptation of the Th-MYCN Mouse Model of Neuroblastoma for Evaluation of Disseminated Disease. International Journal of Molecular Sciences, 24(15), 12071. https://doi.org/10.3390/ijms241512071