


Roles and Preliminary Mechanism of Tobacco cis-Abienol in Inducing Tomato Resistance against Bacterial Wilt
 ,
, 
Abstract
:1. Introduction
2. Results
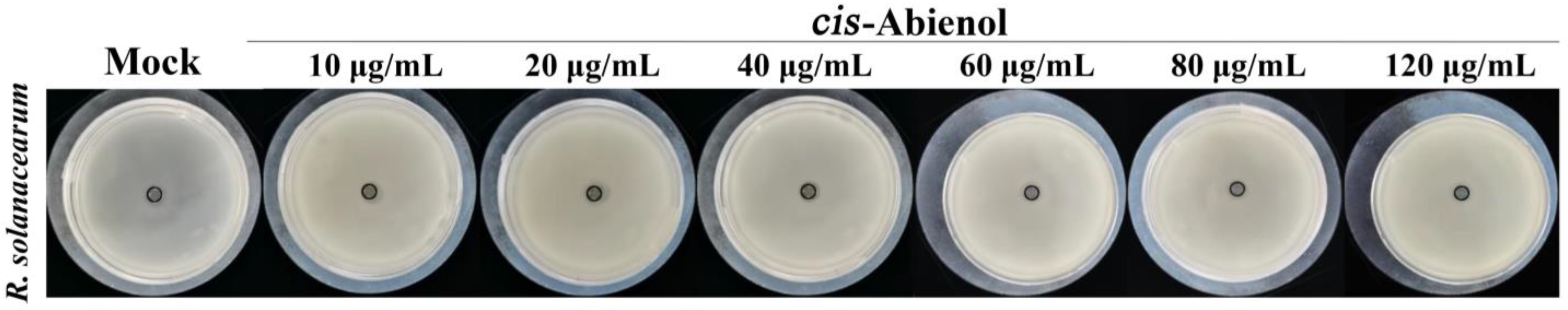
2.1. In Vitro Activity of cis-Abienol against R. solanacearum
2.2. Optimal Conditions for Tomato Seedlings to Acquire a cis-Abienol-Induced Resistance to Bacterial Wilt
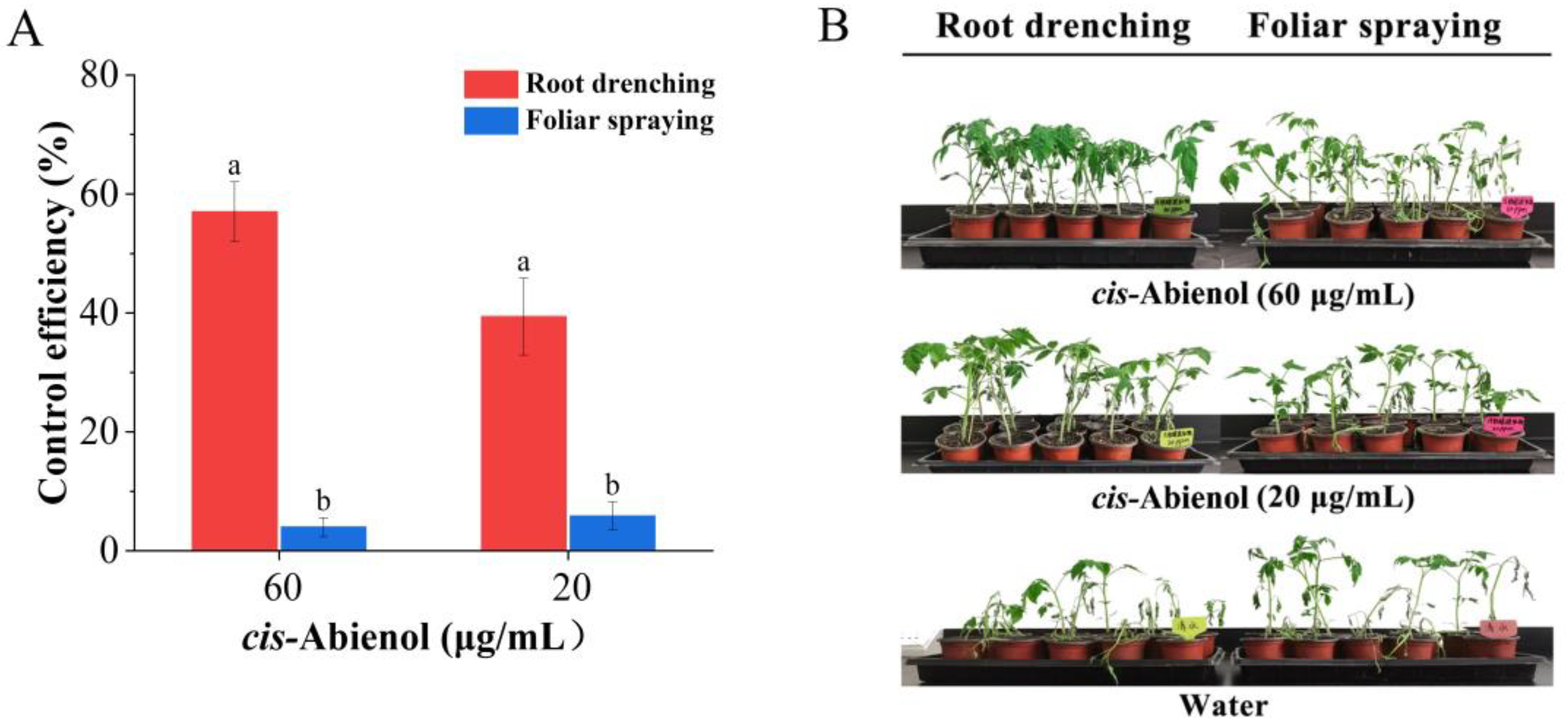
2.2.1. Determination of Optimal Organs for Resistance Induction
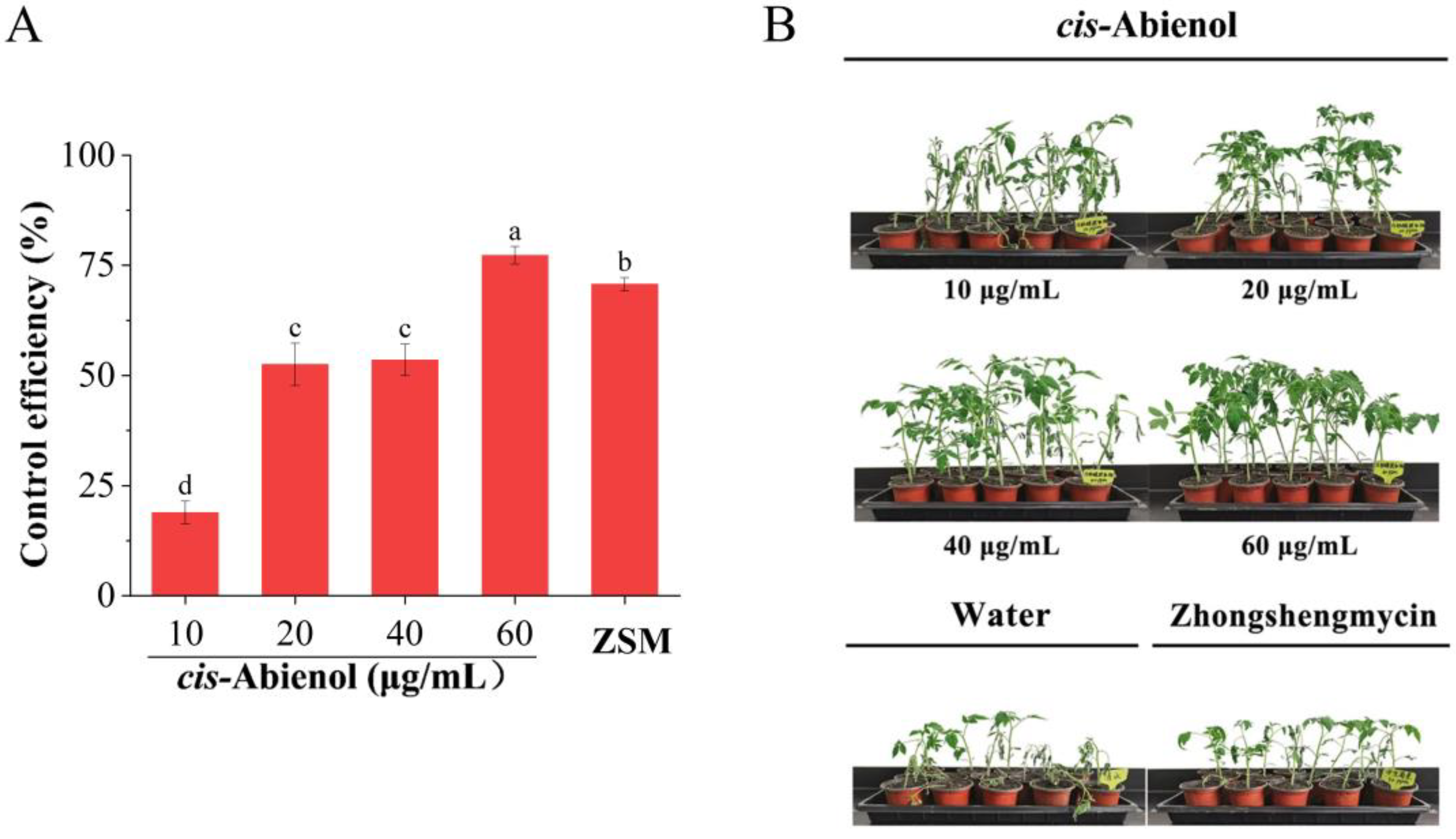
2.2.2. Determination of Optimal cis-Abienol Concentrations for Resistance Induction
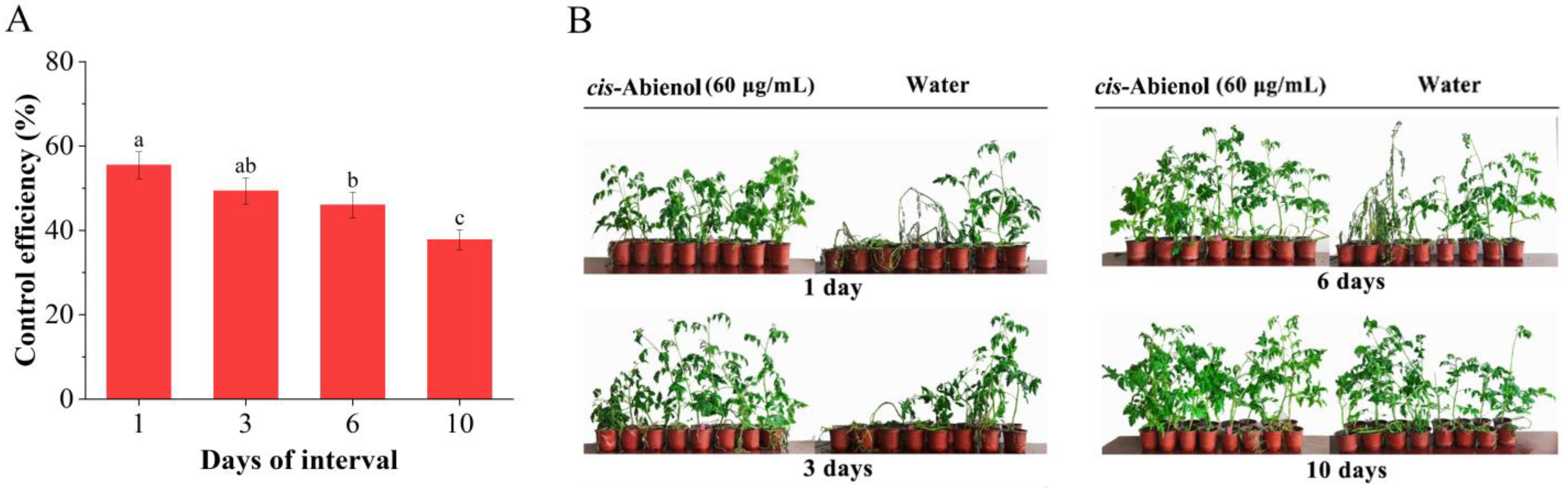
2.2.3. Determination of Optimal Intervals of cis-Abienol Application for Resistance Induction
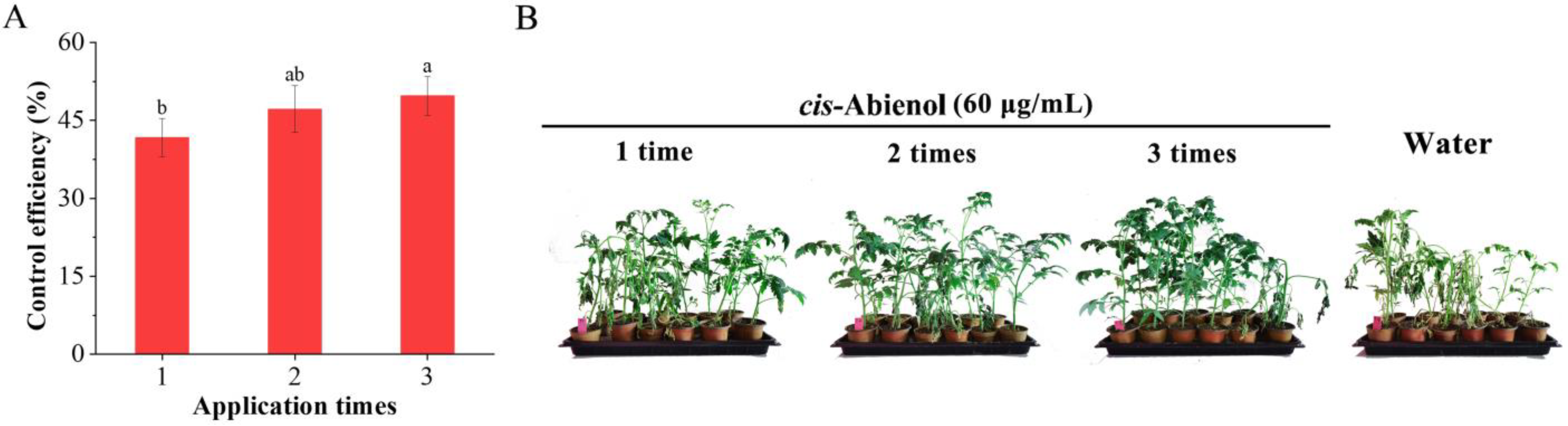
2.2.4. Determination of Suitable Number of cis-Abienol Applications for Resistance Induction
2.3. Effect of cis-Abienol Treatment on the Growth and Development of Tomato Seedlings
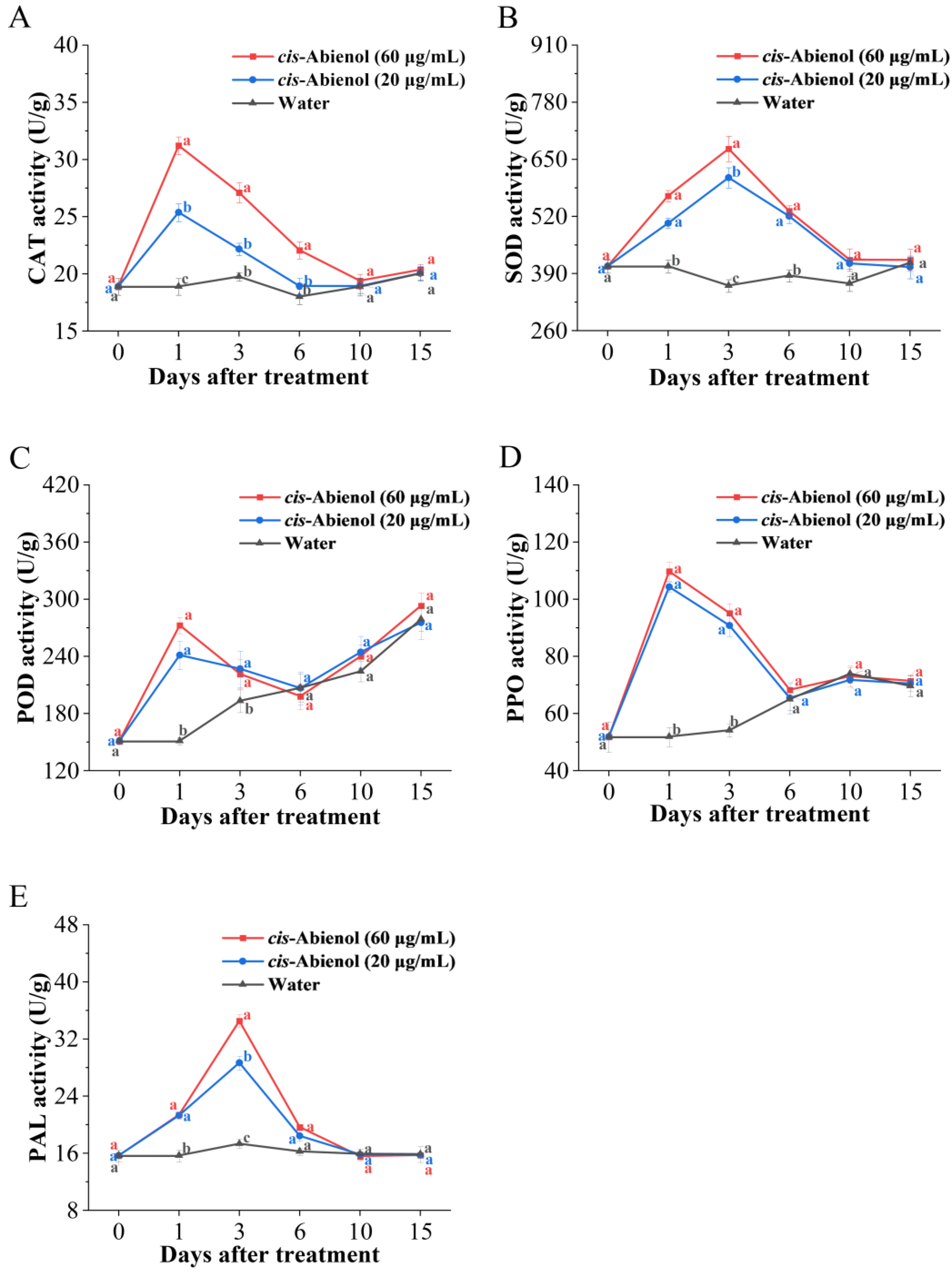
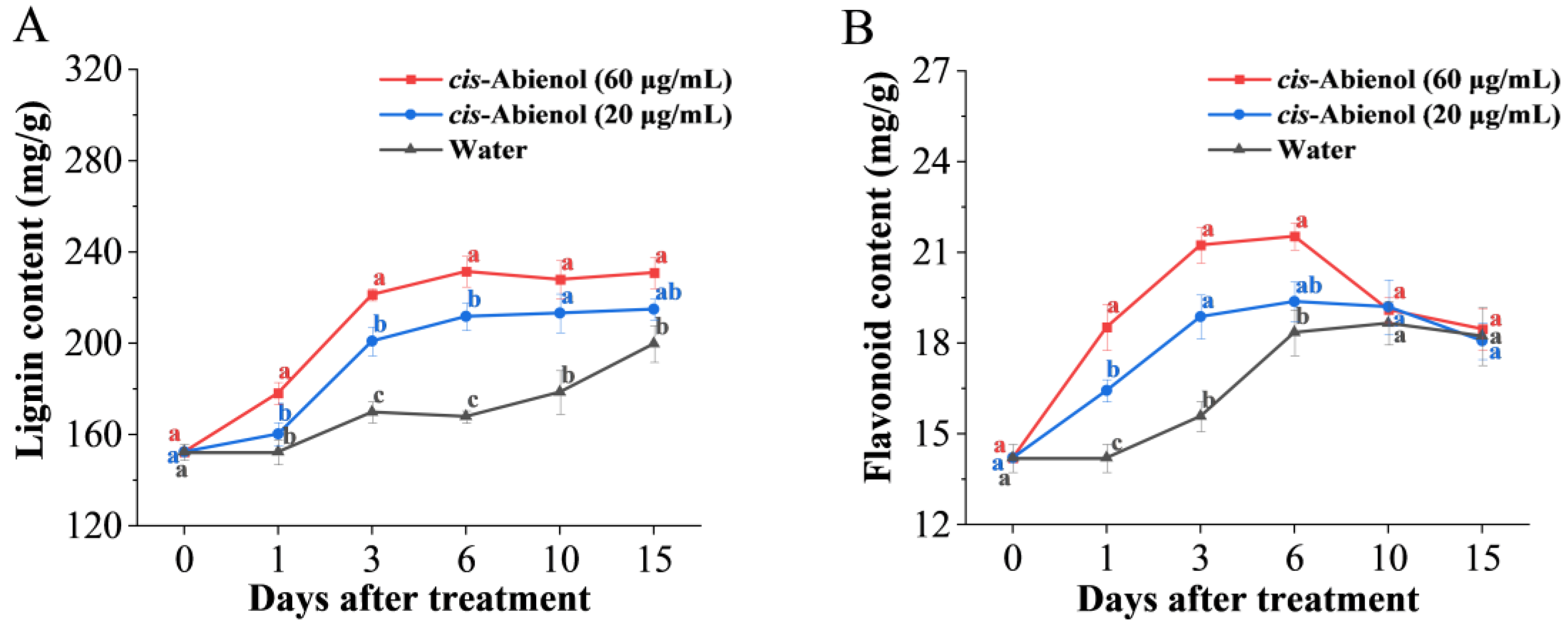
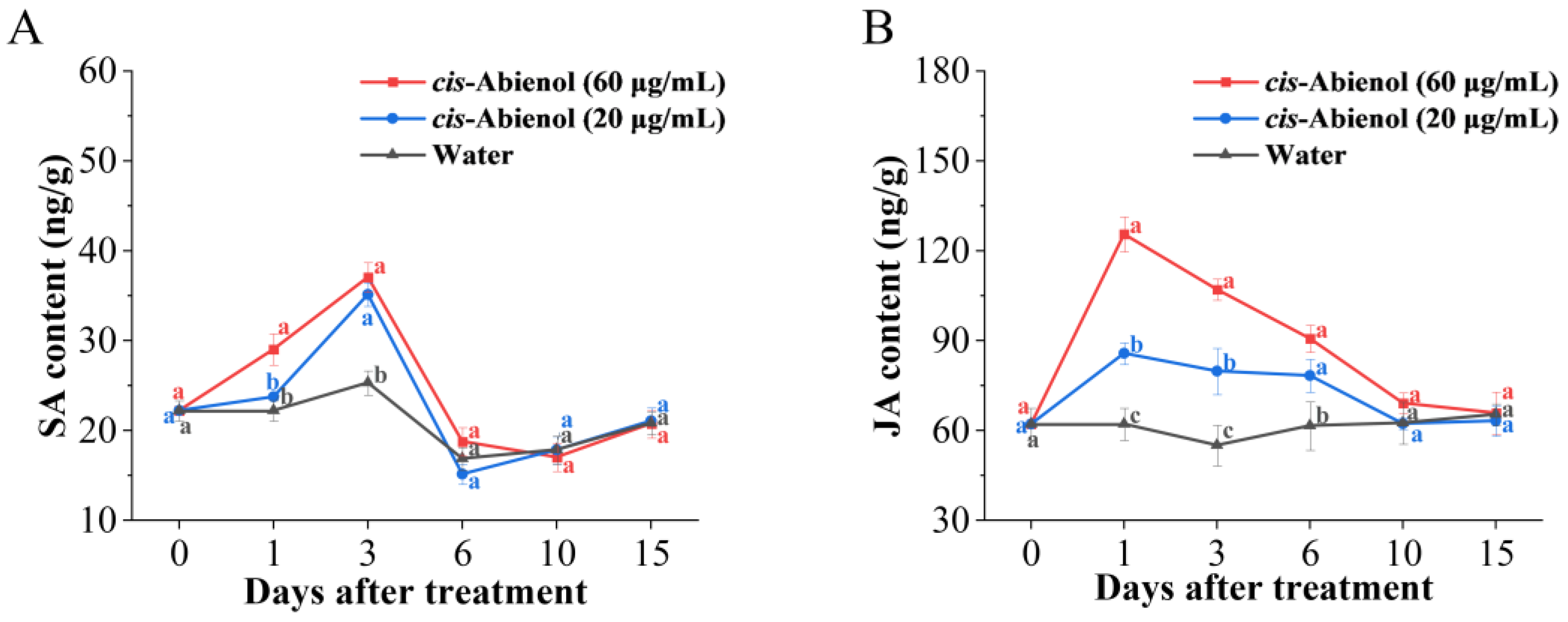
2.4. Effect of cis-Abienol Treatment on the Activities of Defence Enzymes, Phytoalexin, and Phytohormone Content in Tomato Roots
2.4.1. Effects on the Activities of Defence Enzymes
2.4.2. Effects on Phytoalexin Contents
2.4.3. Effect of cis-Abienol Treatment on Plant Hormone Content
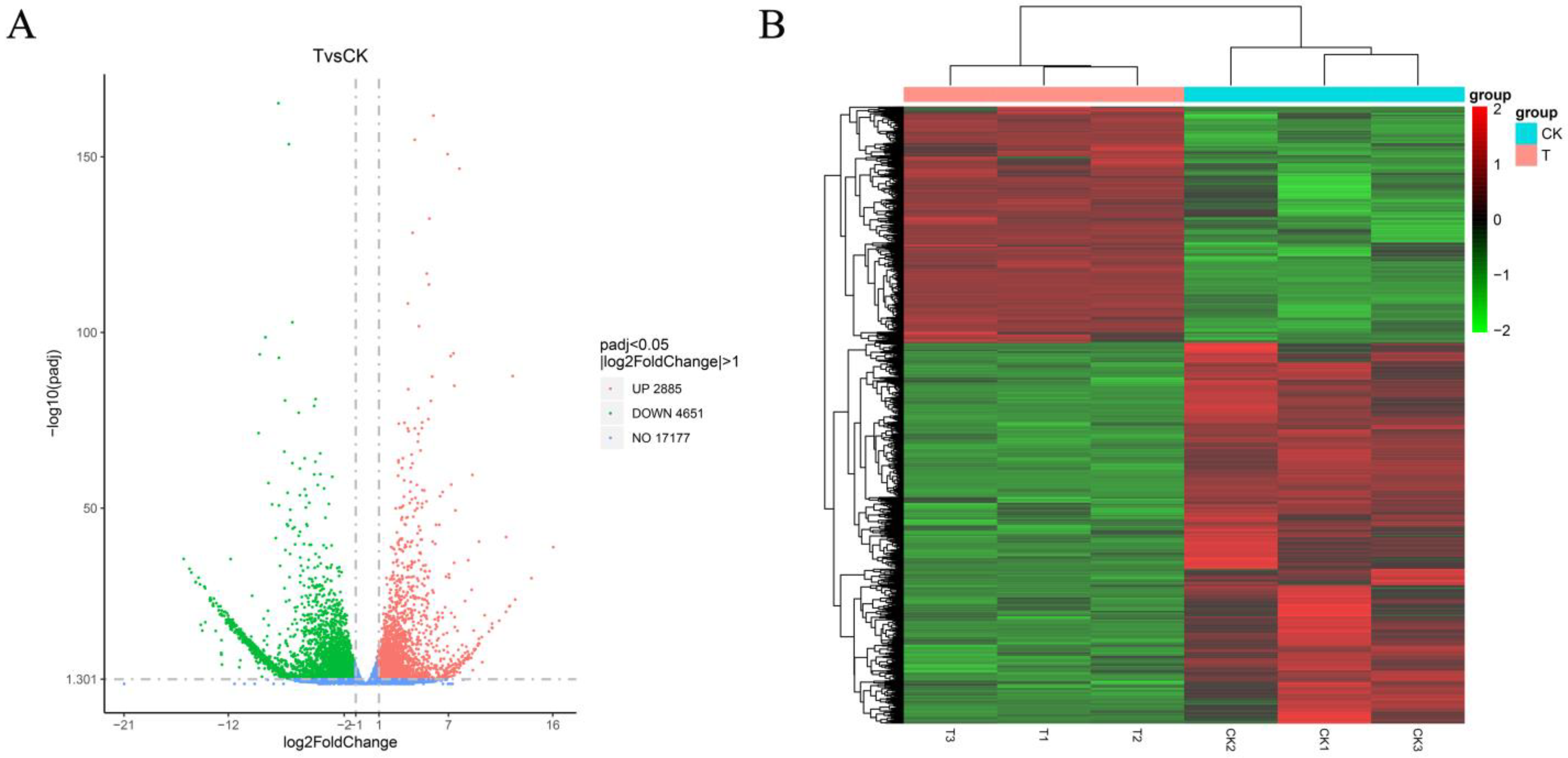
2.5. Analysis of Differentially Expressed Genes Induced by cis-Abienol in Tomato
2.5.1. Statistics of Differentially Expressed Genes
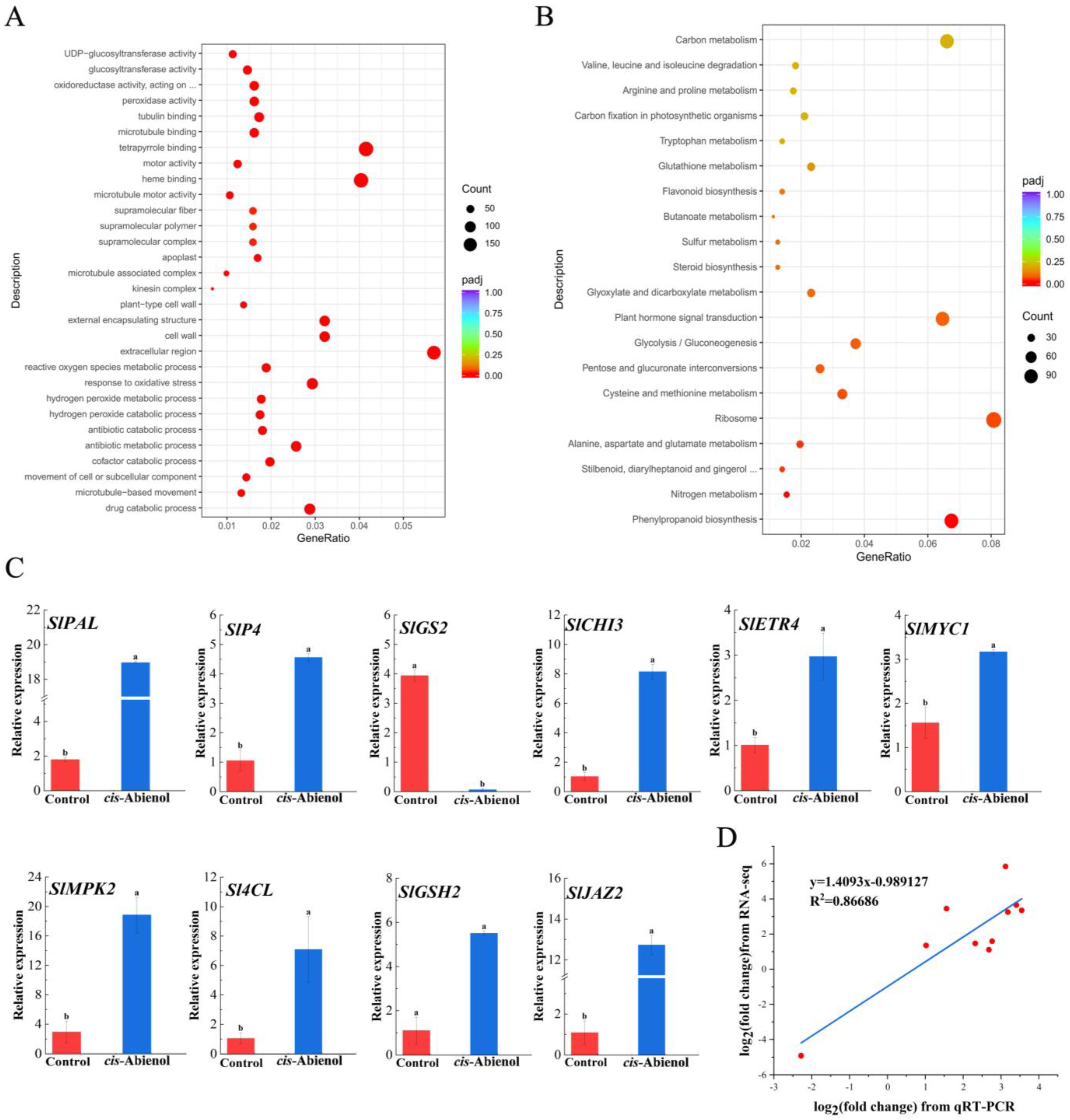
2.5.2. Enrichment of Gene Ontology and Genes of Different Metabolic Pathways
3. Discussion
3.1. Extracellular Signal Conversion Induced by cis-Abienol
3.2. Intracellular Signal Transduction Induced by cis-Abienol
3.3. Signal Transmission between Cells Induced by cis-Abienol
3.4. Production of Phytoalexins Induced by cis-Abienol
4. Materials and Methods
4.1. Collection of Seeds and Pathogens
4.2. Isolation and Identification of cis-Abienol
4.3. In Vitro Activity against R. solanacearum of cis-Abienol
4.4. Determination of the Suitable Conditions of Resistance to Bacterial Wilt Induced by cis-Abienol
4.4.1. Determination of the Suitable Organ for Induction
4.4.2. Determination of the Optimal Concentrations for Resistance Induction
4.4.3. Determination of Optimal Time Intervals for Resistance Induction
4.4.4. Determination of Optimal Times for Resistance Induction
4.5. Determination of Development Period and Agronomic Characters of Tomato Seedlings Induced by cis-Abienol
4.6. Determination of Main Defence Enzyme Activities and Phytoalexin and Phytohormone Content in Tomato Roots
4.6.1. Experimental Design
4.6.2. Determination Methods
4.7. Transcriptome Sequencing of Tomato Roots
4.7.1. Experimental Design
4.7.2. RNA Extraction and Transcriptome Sequencing
4.7.3. Analysis of Major Differential Gene Validation
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PPO | Polyphenol Oxidase |
| POD | Peroxidase |
| PAL | Phenylalanine Ammonia-Lyase |
| CAT | Catalase |
| SOD | Superoxide Dismutase |
| SA | Salicylic Acid |
| JA | Jasmonic Acid |
| MAPK | Mitogen-Activated Protein Kinase |
| BTH | Benzothiadiazole |
| ROS | Reactive Oxygen Species |
| ISR | Induced Systemic Resistance |
| SAR | Systemic Acquired Resistance |
| GO | Gene Ontology |
| KEGG | Genes and Genomes |
| NA | Nutrient Agar |
| HPLC | High-Performance Liquid Chromatography |
References
- Jiang, G.F.; Wei, Z.; Xu, J.; Chen, H.; Zhang, Y.; She, X.M.; Macho, A.P.; Ding, W.; Liao, B. Bacterial wilt in China: History, current status, and future perspectives. Front. Plant Sci. 2017, 8, 1549. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Yang, J.; Tan, Y.J.; Munir, S.; Liu, Q.; Zhang, J.H.; Ji, G.H.; Zhao, Z.X. Ralstonia solanacearum, a deadly pathogen: Revisiting the bacterial wilt biocontrol practices in tobacco and other Solanaceae. Rhizosphere. 2022, 21, 100479. [Google Scholar] [CrossRef]
- Kland, M.J. Chapter 8 teratogenicity of pesticides and other environmental pollutants. In Studies in Environmental Science, Teratogens: Chemicals Which Cause Birth Defects; Meyers, V.K., Ed.; Elsevier: Amsterdam, The Netherlands, 1988; pp. 315–463. [Google Scholar]
- Shen, G.; Zhang, S.; Liu, X.; Jiang, Q.; Ding, W. Soil acidification amendments change the rhizosphere bacterial community of tobacco in a bacterial wilt affected field. Appl. Microbiol. Biotechnol. 2018, 102, 9781–9791. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.; Luo, K.; Jiang, H.D.; Deng, Y.N.; Bai, L.Y.; Zhou, X.M. Combined use of Bacillus subtilis strain B-001 and bactericide for the control of tomato bacterial wilt. Pest Manag. Sci. 2017, 73, 1253–1257. [Google Scholar] [CrossRef]
- Janvier, C.; Villeneuve, F.; Alabouvette, C.; Edel-Hermann, V.; Mateille, T.; Steinberg, C. Soil health through soil disease suppression: Which strategy from descriptors to indicators? Soil Biol. Biochem. 2007, 39, 1–23. [Google Scholar] [CrossRef]
- Kesel, J.D.; Conrath, U.; Flors, V.; Luna, E.; Mageroy, M.H.; Mauch–Mani, B.; Pastor, V.; Pozo, M.J.; Pieterse, C.M.J.; Ton, J.; et al. The induced resistance lexicon: Do’s and don’ts. Trends Plant Sci. 2021, 26, 685–691. [Google Scholar] [CrossRef]
- Abo-Elyousr, K.A.M.; Ibrahim, Y.E.; Balabel, N.M. Induction of disease defensive enzymes in response QWSE to treatment with acibenzolar-S-methyl (ASM) and pseudomonas fluorescens Pf2 and inoculation with Ralstonia solanacearum race 3, biovar 2 (phylotype II): Induction of disease defensive enzymes. J. Phytopathol. 2012, 160, 382–389. [Google Scholar] [CrossRef]
- Hong, J.C.; Timur Momol, M.; Ji, P.; Olson, S.M.; Colee, J.; Jones, J.B. Management of bacterial wilt in tomatoes with thymol and acibenzolar-S-methyl. Crop Prot. 2011, 30, 1340–1345. [Google Scholar] [CrossRef]
- Kurabachew, H.; Wydra, K. Induction of systemic resistance and defense-related enzymes after elicitation of resistance by rhizobacteria and silicon application against Ralstonia solanacearum, in tomato (Solanum lycopersicum). Crop Prot. 2014, 57, 1–7. [Google Scholar] [CrossRef]
- MacIntyre, A.M.; Meline, V.; Gorman, Z.; Augustine, S.P.; Dye, C.J.; Hamilton, C.D.; Iyer-Pascuzzi, A.S.; Kolomiets, M.V.; McCulloh, K.A.; Allen, C. Trehalose increases tomato drought tolerance, induces defenses, and increases resistance to bacterial wilt disease. PLoS ONE 2022, 17, e0266254. [Google Scholar] [CrossRef]
- Fu, H.Z.; Marian, M.; Enomoto, T.; Suga, H.; Shimizu, M. Potential use of L-arabinose for the control of tomato bacterial wilt. Microbes Environ. 2020, 35, ME20106. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, J.; Zhang, P.; Xie, S.N.; Yuan, X.L.; Hou, X.D.; Yan, N.; Fang, Y.D.; Du, Y.M. In vitro and in vivo antifungal activity and preliminary mechanism of cembratrien-diols against Botrytis cinerea. Ind. Crop. Prod. 2022, 154, 112745. [Google Scholar] [CrossRef]
- Jassbi, A.R.; Zare, S.; Asadollahi, M.; Schuman, M.C. Ecological roles and biological activities of specialized metabolites from the genus Nicotiana. Chem. Rev. 2017, 117, 12227–12280. [Google Scholar] [CrossRef] [PubMed]
- Leffingwell, J.C. Chapter 8: Leaf chemistry. basic chemical constituents of tobacco leaf and differences among tobacco types. In Tabacco: Production, Chemistry, and Technology; Layten Davis, D., Nielsen, M.T., Eds.; Blackwell Science: Oxford, UK, 1999; pp. 265–284. [Google Scholar]
- Seo, S.; Gomi, K.; Kaku, H.; Abe, H.; Seto, H.; Nakatsu, S.; Neya, M.; Kobayashi, M.; Nakaho, K.; Ichinose, Y.; et al. Identification of natural diterpenes that inhibit bacterial wilt disease in tobacco, tomato and Arabidopsis. Plant Cell Physiol. 2012, 53, 1432–1444. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, T.; Mizukubo, T.; Abe, H.; Seo, S. Sclareol induces plant resistance to root-knot nematode partially through ethylene-dependent enhancement of lignin accumulation. Mol. Plant Microbe Interact. 2015, 28, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.M.; Xu, X.Y.; Chen, Y.M.; Chen, Z.P.; Wan, S.Q. Several inducers induced the resistance of tobacco to bacterial wilt and synergistic effect mixed with streptomycin. Agrochemicals 2012, 51, 608–610. [Google Scholar]
- Jiang, H.L.; Li, P.; Li, N.J.; Cui, W.W.; Zhang, Y.; Geng, L.N.; Ding, W. Inhibition effects of induced resistance on tobacco resistance to Ralstonia solanacearum. Chin. Agric. Sci. Bull. 2014, 30, 286–290. [Google Scholar]
- Basson, A.E.; Dubery, I.A. Identification of a cytochrome P450 cDNA (CYP98A5) from Phaseolus vulgaris, inducible by 3,5-dichlorosalicylic acid and 2,6-dichloro isonicotinic acid. J. Plant Physiol. 2007, 164, 421–428. [Google Scholar] [CrossRef]
- Jiang, N.H.; Fan, X.Y.; Lin, W.P.; Wang, G.P.; Cai, K.Z. Transcriptome analysis reveals new insights into the bacterial wilt resistance mechanism mediated by silicon in tomato. Int. J. Mol. Sci. 2019, 20, 761. [Google Scholar] [CrossRef] [Green Version]
- Gnonlonfin, G.B.; Sanni, A.; Brimer, L. Review scopoletin–a coumarin phytoalexin with medicinal properties. Crit. Rev. Plant Sci. 2012, 31, 47–56. [Google Scholar] [CrossRef]
- Chong, J.; Baltz, R.; Schmitt, C.; Beffa, R.; Fritig, B.; Saindrenan, P. Downregulation of a pathogen-responsive tobacco UDP-Glc: Phenylpropanoid glucosyltransferase reduces scopoletin glucoside accumulation, enhances oxidative stress, and weakens virus resistance. Plant Cell 2002, 14, 1093–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.L.; Li, M.H.; Yu, S.S.; Cui, Y.H.; Zhang, Y.B.; Wang, L. Cotton resistance induced by 5% amino-oligosaccharin to fusarium wilt and verticillium wilt. China Cott. 2016, 43, 15–18+21. [Google Scholar]
- Walters, D.R.; Ratsep, J.; Havis, N.D. Controlling crop diseases using induced resistance: Challenges for the future. J. Exp. Bot. 2013, 64, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, A.; Brian, E. Double jeopardy: Both overexpression and suppression of a redox-activated plant mitogen-activated protein kinase render tobacco plants ozone sensitive. Plant Cell 2022, 14, 2059–2069. [Google Scholar]
- Wang, L.; Pan, T.W.; Gao, X.H.; An, J.; Ning, C.C.; Li, S.C.; Cai, K.Z. Silica nanoparticles activate defense responses by reducing reactive oxygen species under Ralstonia solanacearum infection in tomato plants. NanoImpact 2022, 28, 100418. [Google Scholar] [CrossRef]
- Zheng, J.R.; Li, Y.Z. Study on BTH-induced resistance of tomato to tomato spot wilt Orthotospovirus (TSWV). Acta Agric. Nucl. Sin. 2022, 36, 489–496. [Google Scholar]
- Zhou, J.M.; Zhang, Y. Plant immunity: Danger perception and signaling. Cell 2022, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Lin, Y.M.; Chao, T.C.; Wang, J.F.; Liu, A.C.; Ho, F.I.; Cheng, C.P. Virus-induced gene silencing reveals the involvement of ethylene-, salicylic acid- and mitogen-activated protein kinase-related defense pathways in the resistance of tomato to bacterial wilt. Physiol. Plantarum. 2009, 136, 324–335. [Google Scholar] [CrossRef]
- Frank, L.; Wenig, M.; Ghirardo, A.; van der Krol, A.; Vlot, A.C.; Schnitzler, J.P.; Rosenkranz, M. Isoprene and β-caryophyllene confer plant resistance via different plant internal signaling pathways. Plant Cell Environ. 2021, 44, 1151–1164. [Google Scholar] [CrossRef]
- Yu, F.; Li, J.; Li, Y.Y.; Li, X.H.; Zheng, L.; Peng, W.X.; Huang, J. Study on the effect of hesperetin on the induction of tobacco resistance to bacterial wilt. Chin. Tob. Sci. 2022, 43, 55–61. [Google Scholar]
- Zhao, P.; Liu, L.; Cao, J.; Wang, Z.; Zhao, Y.; Zhong, N. Transcriptome analysis of tryptophan-induced resistance against potato common scab. Int. J. Mol. Sci. 2022, 23, 8420. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.J.; Du, Y.M.; Wang, A.H.; Liu, Y.H.; Wang, X.J. Determination of cis-abienol in tobacco with ultra-high performance liquid chromatography. Tobacco Sci. Tech. 2020, 53, 64–68+75. [Google Scholar]
- Pastrana-Bonilla, E.; Akoh, C.C.; Sellappan, S.; Krewer, G. Phenolic content and antioxidant capacity of muscadine grapes. J. Agric. Food Chem. 2003, 51, 5497–5503. [Google Scholar] [CrossRef]
- Zhong, T.X.; Li, Y.H.; Li, F.; Peng, X.Q.; Ke, S.W.; Chen, S.; Xie, X.M. Isolation of 4-coumarate CoA Ligase gene from Pennisetum purpureum cv. Huanan and lignin content analysis of transgenic tobacco plants. Acta Bot. Bor-Occid. Sin. 2015, 35, 2355–2364. [Google Scholar]
- Debouba, M.; Maâroufi-Dghimi, H.; Suzuki, A.; Ghorbel, M.H.; Gouia, H. Changes in growth and activity of enzymes involved in nitrate reduction and ammonium assimilation in tomato seedlings in response to NaCl stress. Ann. Bot. 2007, 99, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cis-Abienol (μg/mL) | Sowing to Initial Time of Flowering (d) | Plant Height (cm) | Aboveground Fresh Weight (g) | Underground Fresh Weight (g) |
|---|---|---|---|---|
| 60 | 47–49 | 97.25 ± 7.17 a | 135.68 ± 10.18 a | 30.26 ± 3.10 a |
| 20 | 49–51 | 101.37 ± 4.57 a | 130.42 ± 9.53 a | 29.43 ± 2.49 a |
| Water | 50–51 | 94.82 ± 6.62 a | 127.47 ± 9.42 a | 27.46 ± 3.64 a |
| Gene ID | Name | Description | Log2FC |
|---|---|---|---|
| MAPK cascades | |||
| Solyc08g014425.1 | MPK2 | Mitogen-activated protein kinase 2 | 1.11 |
| Solyc06g005170.3 | MPK3 | Mitogen-activated protein kinase 3 | 1.03 |
| Solyc11g005720.1 | Protein MKS1 | 1.91 | |
| Solyc08g005050.3 | MYC1 | Transcription factor MYC1 | 1.35 |
| Solyc02g082920.3 | CHI3 | Acidic 26 kDa endochitinase precursor | 3.25 |
| Solyc10g055810.2 | CHI9 | Basic 30 kDa endochitinase precursor | 1.92 |
| Solyc01g058720.3 | Calcium-binding protein CP1 | 2.30 | |
| Solyc03g096670.3 | PP2C-2 | Protein phosphatase 2C AHG3 homolog | 2.58 |
| Plant hormone metabolism | |||
| Solyc12g007230.2 | IAA8 | Auxin-regulated IAA8 | 1.25 |
| Solyc10g055260.2 | LAX5 protein | 2.96 | |
| Solyc12g009220.2 | JAZ2 | Jasmonate ZIM-domain protein 2 | 3.35 |
| Solyc03g122190.3 | JAZ3 | Jasmonate ZIM-domain protein 3 | 4.56 |
| Solyc07g043580.3 | Transcription factor PIF4 | 3.86 | |
| Solyc01g102300.3 | Transcription factor PIF3 isoform X1 | 2.07 | |
| Solyc05g052980.3 | Protein phosphatase 2C 37 | 1.23 | |
| Solyc06g053710.3 | ETR4 | Ethylene receptor ETR4 precursor | 3.45 |
| Solyc09g089930.2 | EREB | Ethylene-responsive element binding protein | 3.70 |
| Solyc03g093130.3 | Xyloglucan endotransglucosylase-hydrolase XTH3 precursor | −1.69 | |
| Solyc00g174330.3 | P4 | Pathogenesis-related leaf protein 4 precursor | 5.85 |
| Solyc05g050280.3 | Jasmonic acid-amido synthetase JAR1 | 1.60 | |
| Solyc11g011260.1 | GAI | DELLA protein GAI | −1.20 |
| Solyc00g174330.3 | P4 | Pathogenesis-related leaf protein 4 precursor | 5.85 |
| Solyc01g106630.2 | PR1 protein precursor | 1.52 | |
| Phenylpropanoid biosynthesis | |||
| Solyc09g007910.3 | Phenylalanine ammonia-lyase | −1.83 | |
| Solyc10g086180.2 | Phenylalanine ammonia-lyase | 3.13 | |
| Solyc03g036470.2 | Low-quality protein: phenylalanine ammonia-lyase-like | 3.13 | |
| Solyc05g056170.3 | PAL | Phenylalanine ammonia-lyase | 3.65 |
| Solyc12g042460.2 | 4CL | 4-coumarate-CoA ligase | 1.59 |
| Solyc05g010320.3 | CHI1 | Chalcone-flavonone isomerase | 2.27 |
| Solyc12g055820.2 | Probable cinnamyl alcohol dehydrogenase 1 | 1.32 | |
| Solyc10g078540.2 | Glutamate dehydrogenase | 1.85 | |
| Solyc01g098610.3 | GSH2 | Glutathione synthetase, chloroplastic | 1.47 |
| Solyc01g080280.3 | Glutamine synthetase | −4.92 | |
| Solyc03g098240.3 | Glutamate decarboxylase | 3.01 | |
| Solyc01g080280.3 | GS2 | Glutamine synthetase | −4.92 |
| Solyc01g098610.3 | Glutathione synthetase, chloroplastic | 1.46 | |
| Secondary metabolism | |||
| Solyc05g047530.3 | Cytochrome P450 CYP73A100 | 4.54 | |
| Solyc01g105590.2 | AT3 | AT3 protein | −7.47 |
| Solyc10g078220.2 | Cytochrome P450 98A3 | 4.70 | |
| Solyc01g008110.3 | CYP51 | Sterol C14-demetylase | 1.06 |
| Solyc10g078230.2 | Cytochrome P450 98A2-like | 3.02 | |
| Solyc02g093270.3 | Caffeoyl-CoA O-methyltransferase-like | 2.51 | |
| Solyc08g074620.2 | Polyphenol oxidase E, chloroplastic | 9.80 | |
| Solyc07g032740.3 | Aspartate aminotransferase, cytoplasmic | 3.08 | |
| Solyc01g110290.3 | SQS1 | Squalene synthase | 1.63 |
| Solyc01g101210.3 | TPS33 | Viridiflorene synthase | 2.48 |
| Solyc12g006510.2 | SlTTS1 | Beta-amyrin synthase | 4.45 |
| Disease Grade | Standard |
|---|---|
| 0 | No visible disease in any plant part. |
| 1 | Chlorotic spots were occasionally observed on the stem or the leaves below 1/2 of the diseased side wilted. |
| 3 | Black stripes were observed on the stem but at a height shorter than 1/2 of the stem height or 1/2 to 2/3 of the leaves on the diseased side wilted. |
| 5 | Black stripes on the stem exceed 1/2 of the stem height but do not reach the top of the stem, or more than 2/3 of the leaves on the diseased side wilted. |
| 7 | Black stripes on the stem reach the top of the stem, or all leaves of the infected plant were completely wilted. |
| 9 | The plant almost died. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Gui, Z.; Yan, N.; Wang, Q.; Zhang, Z.; Zhang, H.; Sun, F.; Han, X.; Du, Y. Roles and Preliminary Mechanism of Tobacco cis-Abienol in Inducing Tomato Resistance against Bacterial Wilt. Int. J. Mol. Sci. 2023, 24, 12226. https://doi.org/10.3390/ijms241512226
Sun Y, Gui Z, Yan N, Wang Q, Zhang Z, Zhang H, Sun F, Han X, Du Y. Roles and Preliminary Mechanism of Tobacco cis-Abienol in Inducing Tomato Resistance against Bacterial Wilt. International Journal of Molecular Sciences. 2023; 24(15):12226. https://doi.org/10.3390/ijms241512226
Chicago/Turabian StyleSun, Yuqing, Zuqing Gui, Ning Yan, Qian Wang, Zhongfeng Zhang, Hongbo Zhang, Feifei Sun, Xiao Han, and Yongmei Du. 2023. "Roles and Preliminary Mechanism of Tobacco cis-Abienol in Inducing Tomato Resistance against Bacterial Wilt" International Journal of Molecular Sciences 24, no. 15: 12226. https://doi.org/10.3390/ijms241512226
APA StyleSun, Y., Gui, Z., Yan, N., Wang, Q., Zhang, Z., Zhang, H., Sun, F., Han, X., & Du, Y. (2023). Roles and Preliminary Mechanism of Tobacco cis-Abienol in Inducing Tomato Resistance against Bacterial Wilt. International Journal of Molecular Sciences, 24(15), 12226. https://doi.org/10.3390/ijms241512226