
Amino Acid Profiling of Follicular Fluid in Assisted Reproduction Reveals Important Roles of Several Amino Acids in Patients with Insulin Resistance

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Demographic and Clinical Data of Patients
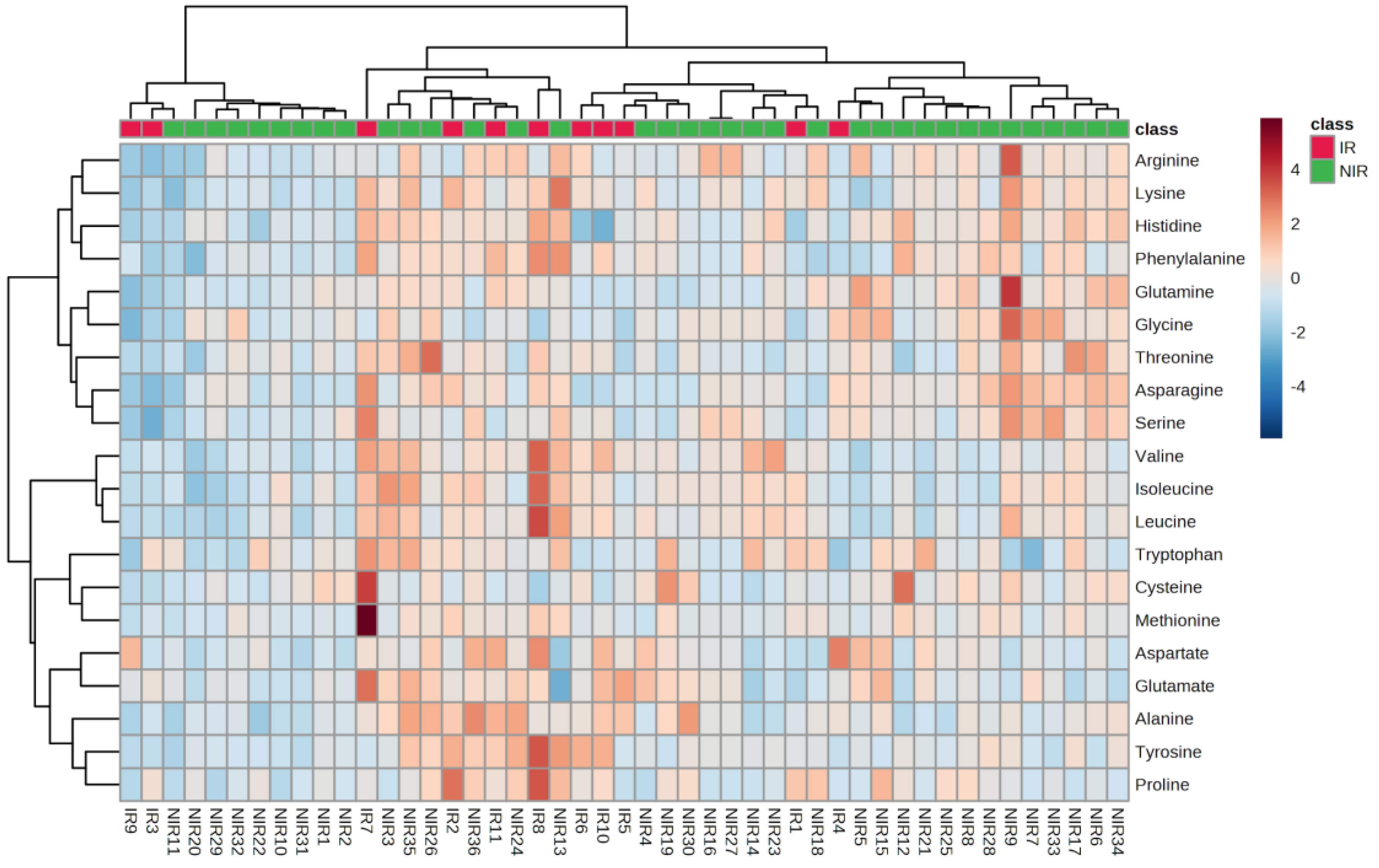
2.2. Amino Acid (AA) Analysis of the Follicular Fluid Samples
2.2.1. Comparison of Amino Acid Content According to Insulin Resistance and Non-Insulin Resistance Groups
2.2.2. Comparison of Amino Acid Contents Based on Body Mass Index (BMI)
2.2.3. Comparison of Amino Acid Contents Based on the Age of the Patients
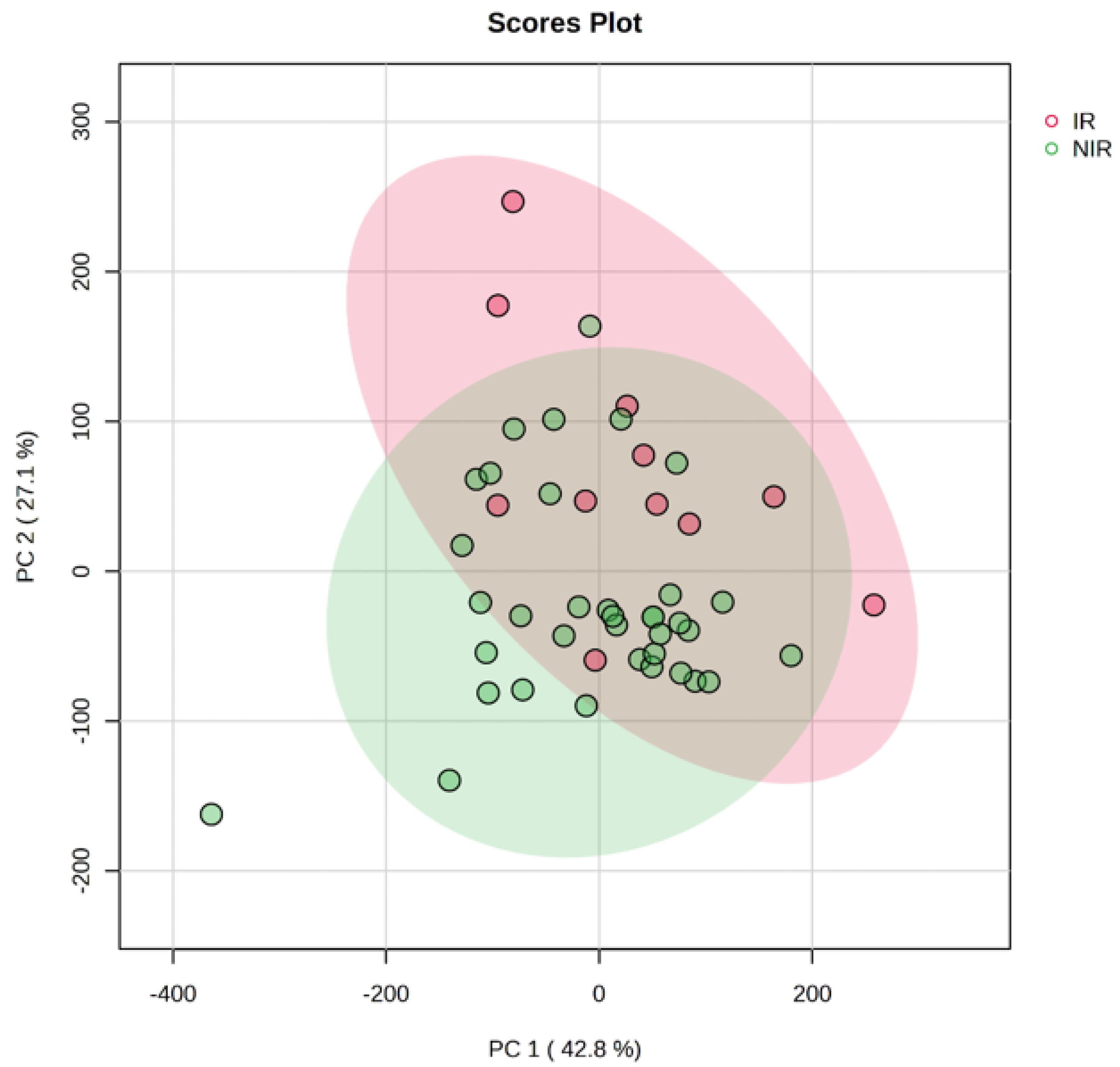
2.3. Multivariate PCA and PLS-DA Analysis
2.4. Potentially Important Metabolites—Biomarker Analysis
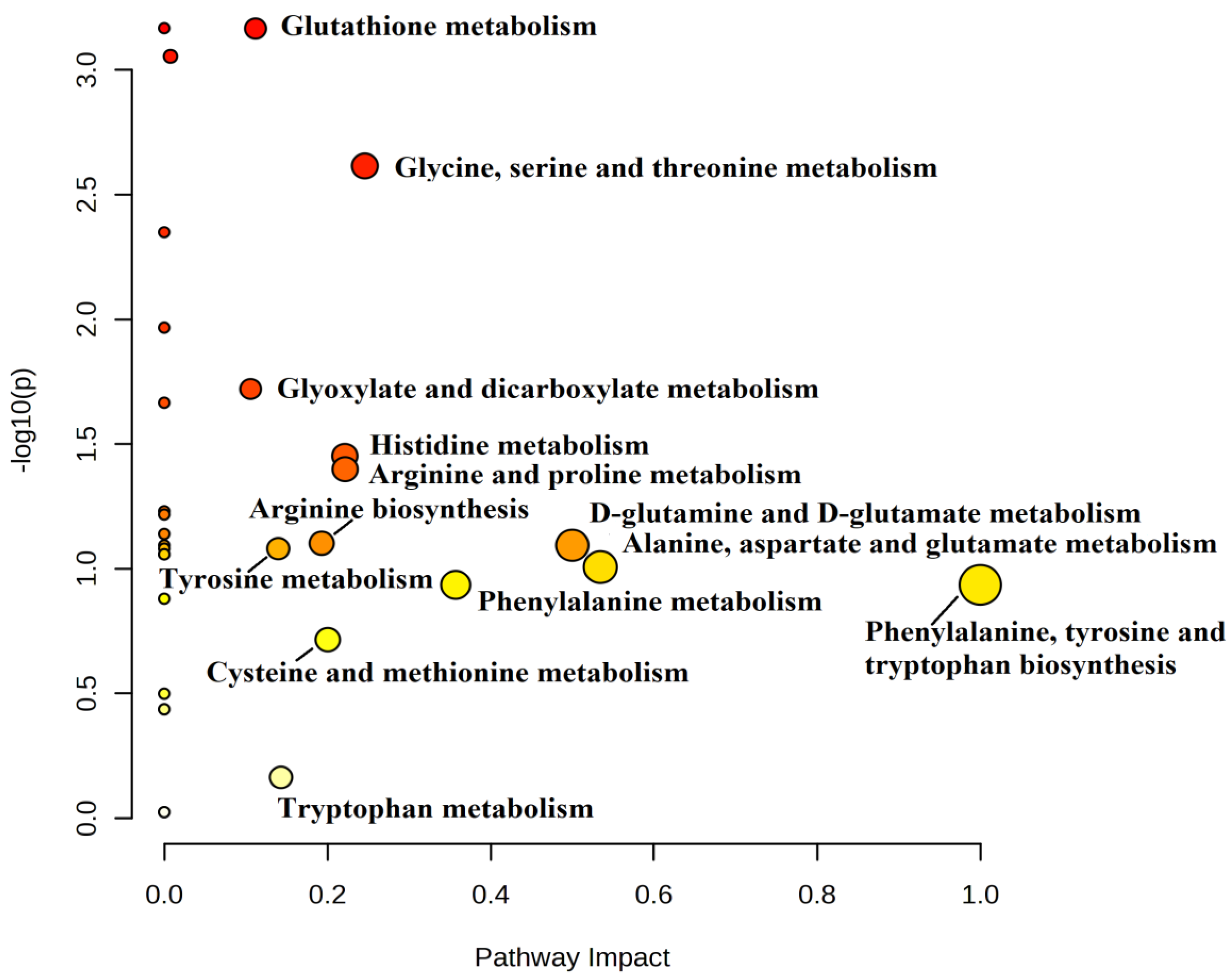
2.5. Metabolic Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. Patient Enrollment
4.2. Stimulation and Collection of Follicular Fluid
4.3. Sample Processing and Measurement
4.3.1. Reagents
4.3.2. Sample Preparation for the UHPLC Measurement
4.3.3. Derivatization
4.3.4. Parameters of the UHPLC Method
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simionescu, G.; Doroftei, B.; Maftei, R.; Obreja, B.-E.; Anton, E.; Grab, D.; Ilea, C.; Anton, C. The complex relationship between infertility and psychological distress (Review). Exp. Ther. Med. 2021, 21, 306. [Google Scholar] [CrossRef] [PubMed]
- Westerman, R.; Kuhnt, A.-K. Metabolic risk factors and fertility disorders: A narrative review of the female perspective. Reprod. Biomed. Soc. Online 2022, 14, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; He, D.; Luo, C.; Lin, X.; Wu, J.; Yin, X.; Jia, C.; Pan, Q.; Dong, X.; Zheng, F.; et al. Metabolic Syndrome and Its Components Are Associated With Altered Amino Acid Profile in Chinese Han Population. Front. Endocrinol. 2022, 12, 795044. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [Green Version]
- McCracken, E.; Monaghan, M.; Sreenivasan, S. Pathophysiology of the metabolic syndrome. Clin. Dermatol. 2018, 36, 14–20. [Google Scholar] [CrossRef]
- Sakumoto, T.; Tokunaga, Y.; Tanaka, H.; Nohara, M.; Motegi, E.; Shinkawa, T.; Nakaza, A.; Higashi, M. Insulin resistance/hyperinsulinemia and reproductive disorders in infertile women. Reprod. Med. Biol. 2010, 9, 185–190. [Google Scholar] [CrossRef]
- Lebowski, J.H.; Fergana, C.; Gawałek, M.; Skowronska, B.; Fichna, P.; Lehman, M.N. Insulin: Its role in the central control of reproduction. Physiol. Behav. 2014, 133, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Norman, R.J. Improving reproductive performance in overweight/obese women with effective weight management. Hum. Reprod. Update 2004, 10, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Dag, Z.O.; Dilbaz, B. Impact of obesity on infertility in women. J. Turk. Gynecol. Assoc. 2015, 16, 111–117. [Google Scholar] [CrossRef]
- Jungheim, E.S.; Travieso, J.L.; Carson, K.R.; Moley, K.H. Obesity and Reproductive Function. Obstet. Gynecol. Clin. North Am. 2012, 39, 479–493. [Google Scholar] [CrossRef] [Green Version]
- Maheshwari, A.; Stofberg, L.; Bhattacharya, S. Effect of overweight and obesity on assisted reproductive technology—A systematic review. Hum. Reprod. Updat. 2007, 13, 433–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ter, P.É.; Fedorcsa, F.F.; Storeng, R.; Dale, O.; Tanbo, T.; Åbyholm, T. Acta Obstetricia et Gynecologica Scandinavica Obesity is a risk factor for early pregnancy loss after IVF or ICSI. Acta Obs. Gynecol. Scand. 2020, 79, 43–48. [Google Scholar]
- Dokras, A.; Baredziak, L.; Blaine, J.; Syrop, C.; Vanvoorhis, B.J.; Sparks, A. Obstetric Outcomes after In Vitro Fertilization in Obese and Morbidly Obese Women Level of Evidence: II-2. 2006. Available online: http://journals.lww.com/greenjournal (accessed on 23 July 2023).
- van Swieten, E.C.; van der Leeuw-Harmsen, L.; Badings, E.A.; van der Linden, P.J. Obesity and clomiphene challenge test as predictors of outcome of in vitro fertilization and intracytoplasmic sperm injection. Gynecol. Obstet. Investig. 2005, 59, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Metwally, M.; Cutting, R.; Tipton, A.; Skull, J.; Ledger, W.; Li, T. Effect of increased body mass index on oocyte and embryo quality in IVF patients. Reprod. Biomed. Online 2007, 15, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Chamayou, S. Oocyte quality in assisted reproduction techniques. Minerva Endocrinol. 2022, 47, 89–98. [Google Scholar] [CrossRef]
- Hurrle, S.; Hsu, W.H. The etiology of oxidative stress in insulin resistance. Biomed. J. 2017, 40, 257–262. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Várnagy, Á.; Kőszegi, T.; Györgyi, E.; Szegedi, S.; Sulyok, E.; Prémusz, V.; Bódis, J. Levels of total antioxidant capacity and 8-hydroxy-2′-deoxyguanosine of serum and follicular fluid in women undergoing in vitro fertilization: Focusing on endometriosis. Hum. Fertil. 2018, 23, 200–208. [Google Scholar] [CrossRef]
- Hood, R.B.; Liang, D.; Tan, Y.; Ford, J.; Souter, I.; Jones, D.P.; Hauser, R.; Gaskins, A.J. Characterizing the follicular fluid metabolome: Quantifying the correlation across follicles and differences with the serum metabolome. Fertil. Steril. 2022, 118, 970–979. [Google Scholar] [CrossRef]
- Arya, B.K.; Haq, A.U.; Chaudhury, K. Oocyte quality reflected by follicular fluid analysis in poly cystic ovary syndrome (PCOS): A hypothesis based on intermediates of energy metabolism. Med. Hypotheses 2012, 78, 475–478. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, Z.; Zhang, Y.; Li, J.; Ruan, X.; Wan, Q.; Yin, T.; Zou, Y.; Chen, S.; Zhang, Y. Nontargeted metabolomics analysis of follicular fluid in patients with endometriosis provides a new direction for the study of oocyte quality. Medcomm 2023, 4, e302. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, K.; Lunn, L.; Kwong, W.; Wonnacott, K.; Linforth, R.; Craigon, J. Amino acid and fatty acid composition of follicular fluid as predictors of in-vitro embryo development. Reprod. Biomed. Online 2008, 16, 859–868. [Google Scholar] [CrossRef]
- Józwik, M.; Józwik, M.; Teng, C.; Battaglia, F.C. Amino acid, ammonia and urea concentrations in human pre-ovulatory ovarian follicular fluid. Hum. Reprod. 2006, 21, 2776–2782. [Google Scholar] [CrossRef] [PubMed]
- Kirsipuu, T.; Laks, K.; Velthut-Meikas, A.; Levkov, L.; Salumets, A.; Palumaa, P. Comprehensive elucidation of amino acid profile in human follicular fluid and plasma of in vitrofertilization patients. Gynecol. Endocrinol. 2015, 31, 9–17. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kido, J.; Matsumoto, S.; Shimizu, K.; Nakamura, K. Associations among amino acid, lipid, and glucose metabolic profiles in childhood obesity. BMC Pediatr. 2019, 19, 273. [Google Scholar] [CrossRef]
- Owei, I.; Umekwe, N.; Stentz, F.; Wan, J.; Dagogo-Jack, S. Amino acid signature predictive of incident prediabetes: A case-control study nested within the longitudinal pathobiology of prediabetes in a biracial cohort. Metabolism 2019, 98, 76–83. [Google Scholar] [CrossRef]
- Cetica, P.; Pintos, L.; Dalvit, G.; Beconi, M. Involvement of enzymes of amino acid metabolism and tricarboxylic acid cycle in bovine oocyte maturation in vitro. Reproduction 2003, 126, 753–763. [Google Scholar] [CrossRef]
- Uhde, K.; Van Tol, H.T.A.; Stout, T.A.E.; Roelen, B.A.J. Metabolomic profiles of bovine cumulus cells and cumulus-oocyte-complex-conditioned medium during maturation in vitro. Sci. Rep. 2018, 8, 9477. [Google Scholar] [CrossRef] [Green Version]
- Razak, M.A.; Begum, P.S.; Viswanath, B.; Rajagopal, S. Multifarious Beneficial Effect of Nonessential Amino Acid, Glycine: A Review. Oxidative Med. Cell. Longev. 2017, 2017, 1716701. [Google Scholar] [CrossRef] [Green Version]
- Seli, E.; Babayev, E.; Collins, S.C.; Nemeth, G.; Horvath, T.L. Minireview: Metabolism of female reproduction: Regulatory mechanisms and clinical implications. Mol. Endocrinol. 2014, 28, 790–804. [Google Scholar] [CrossRef] [Green Version]
- Yan-Do, R.; MacDonald, P.E. Impaired “Glycine”-mia in type 2 diabetes and potential mechanisms contributing to glucose homeostasis. Endocrinology 2017, 158, 1064–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, A.; Bassot, A.; Bulteau, A.-L.; Pirola, L.; Morio, B. Glycine metabolism and its alterations in obesity and metabolic diseases. Nutrients 2019, 11, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Guo, Q.; Wang, Y.-M.; Li, Z.-Y.; Kang, J.-D.; Yin, X.-J.; Zheng, X. Glycine treatment enhances developmental potential of porcine oocytes and early embryos by inhibiting apoptosis1. J. Anim. Sci. 2018, 96, 2427–2437. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Foote, R. Effects of amino acids on the development of in-vitro matured/in-vitro fertilization bovine embryos in a simple protein-free medium. 1995. Available online: https://academic.oup.com/humrep/article-abstract/10/11/2985/674301 (accessed on 26 July 2023).
- Rasmussen, B.F.; Ennis, M.A.; Dyer, R.A.; Lim, K.; Elango, R. Glycine, a Dispensable Amino Acid, Is Conditionally Indispensable in Late Stages of Human Pregnancy. J. Nutr. 2021, 151, 361–369. [Google Scholar] [CrossRef]
- Otasevic, V.; Korac, B. Amino Acids: Metabolism. In Encyclopedia of Food and Health; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 149–155. [Google Scholar] [CrossRef]
- Jeong, B.-S.; Yang, X. Cysteine, glutathione, and percoll treatments improve porcine oocyte maturation and fertilization in vitro. Mol. Reprod. Dev. 2001, 59, 330–335. [Google Scholar] [CrossRef]
- Jain, S.K.; Micinski, D.; Huning, L.; Kahlon, G.; Bass, P.F.; Levine, S.N. Vitamin D and L-cysteine levels correlate positively with GSH and negatively with insulin resistance levels in the blood of type 2 diabetic patients. Eur. J. Clin. Nutr. 2014, 68, 1148–1153. [Google Scholar] [CrossRef] [Green Version]
- Brosnan, J.T.; Brosnan, M.E. Glutamate: A truly functional amino acid. Amino Acids 2013, 45, 413–418. [Google Scholar] [CrossRef]
- Balikcioglu, P.G.; Trub, C.J.; Balikcioglu, M.; Ilkayeva, O.; White, P.J.; Muehlbauer, M.; Bain, J.R.; Armstrong, S.; Freemark, M. Branched-chain α-keto acids and glutamate/glutamine: Biomarkers of insulin resistance in childhood obesity. Endocrinol. Diabetes Metab. 2023, 6, e388. [Google Scholar] [CrossRef]
- Sekhon, L.H.; Gupta, S.; Kim, Y.; Agarwal, A. Female Infertility and Antioxidants. Curr. Women s Heal. Rev. 2010, 6, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Anchordoquy, J.P.; Lizarraga, R.M.; Anchordoquy, J.M.; Nikoloff, N.; Rosa, D.E.; Fabra, M.C.; Peral-García, P.; Furnus, C.C. Effect of cysteine, glutamate and glycine supplementation to in vitro fertilization medium during bovine early embryo development. Reprod. Biol. 2019, 19, 349–355. [Google Scholar] [CrossRef]
- Mukherjee, A.; Malik, H.; Saha, A.P.; Dubey, A.; Singhal, D.K.; Boateng, S.; Saugandhika, S.; Kumar, S.; De, S.; Guha, S.K.; et al. Resveratrol treatment during goat oocytes maturation enhances developmental competence of parthenogenetic and hand-made cloned blastocysts by modulating intracellular glutathione level and embryonic gene expression. J. Assist. Reprod. Genet. 2014, 31, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Lawson, G.W.; Nakamura, B.N.; Ortiz, L.; Hur, J.A.; Kavanagh, T.J.; Luderer, U. Glutathione-deficient mice have increased sensitivity to transplacental Benzo[a]pyrene-Induced premature ovarian failure and ovarian tumorigenesis. Cancer Res. 2013, 73, 908–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IR [Mean ± SD] (n = 11) | NIR [Mean ±SD] (n = 36) | |
|---|---|---|
| Age | 34.45 ±6.78 | 35.58 ± 4.69 |
| BMI | 32.95 ± 5.55 | 23.58 ± 4.54 |
| Number of oocytes retrieved | 12.27 ± 9.83 | 10.19 ± 6.81 |
| Number of fertilized oocytes | 3.45 ± 3.88 | 4.42 ± 3.72 |
| Number of IVF cycles | 1.9 ± 0.94 | 2.08 ± 0.77 |
| Cause of infertility | ||
| Male factor | 4 (36.36%) | 12 (33.3%) |
| Female factor | 3 (27.27%) | 15 (42.6%) |
| Combined male–female | 3 (27.27%) | 6 (16.6%) |
| Unexplained | 1 (9%) | 3 (8.3%) |
| Significance (p) Values | |||
|---|---|---|---|
| IR/NIR | BMI | Age | |
| Aspartate | 0.02 | 0.029 | 0.297 |
| Glutamate | 0.106 | 0.008 | 0.322 |
| Asparagine | 0.42 | 0.234 | 0.813 |
| Serine | 0.061 | 0.12 | 0.182 |
| Glutamine | 0.13 | 0.234 | 0.813 |
| Histidine | 0.72 | 0.307 | 0.747 |
| Glycine | <0.001 | 0.009 | 0.228 |
| Threonine | 0.951 | 0.593 | 0.966 |
| Arginine | 0.072 | 0.264 | 0.132 |
| Alanine | 0.19 | 0.068 | 0.312 |
| Tyrosine | 0.647 | 0.274 | 0.074 |
| Cysteine | 0.037 | 0.459 | 0.54 |
| Valine | 0.111 | 0.068 | 0.245 |
| Methionine | 0.594 | 0.481 | 0.813 |
| Tryptophan | 0.719 | 1 | 0.636 |
| Phenylalanine | 0.683 | 0.576 | 0.78 |
| Isoleucine | 0.351 | 0.166 | 0.401 |
| Leucine | 0.29 | 0.174 | 0.389 |
| Lysine | 0.931 | 0.395 | 0.254 |
| Proline | 0.227 | 0.369 | 0.88 |
| Amino Acid | AUC | p Value |
|---|---|---|
| Glycine | 0.848 | <0.001 |
| Aspartate | 0.732 | 0.004 |
| Cysteine | 0.708 | 0.365 |
| Serine | 0.689 | 0.090 |
| Histidine | 0.681 | 0.029 |
| Arginine | 0.681 | 0.071 |
| Glutamate | 0.664 | 0.058 |
| Valine | 0.661 | 0.060 |
| Glutamine | 0.65 | 0.087 |
| Alanine | 0.633 | 0.316 |
| Proline | 0.623 | 0.046 |
| Leucine | 0.608 | 0.190 |
| Isoleucine | 0.595 | 0.246 |
| Asparagine | 0.582 | 0.425 |
| Methionine | 0.555 | 0.082 |
| Tyrosine | 0.547 | 0.083 |
| Phenylalanine | 0.542 | 0.383 |
| Tryptophan | 0.537 | 0.685 |
| Lysine | 0.510 | 0.945 |
| Threonine | 0.507 | 0.613 |
| Pathway Name | Impact | p Value |
|---|---|---|
| Phenylalanine, tyrosine, and tryptophan biosynthesis | 1 | 0.116 |
| Alanine, aspartate, and glutamate metabolism | 0.534 | 0.098 |
| D-Glutamine and D-glutamate metabolism | 0.500 | 0.081 |
| Phenylalanine metabolism | 0.357 | 0.116 |
| Glycine, serine, and threonine metabolism | 0.246 | 0.002 |
| Arginine and proline metabolism | 0.222 | 0.039 |
| Histidine metabolism | 0.221 | 0.035 |
| Cysteine and methionine metabolism | 0.200 | 0.192 |
| Arginine biosynthesis | 0.193 | 0.079 |
| Tryptophan metabolism | 0.143 | 0.685 |
| Tyrosine metabolism | 0.140 | 0.083 |
| Glutathione metabolism | 0.112 | 0.0006 |
| Glyoxylate and dicarboxylate metabolism | 0.106 | 0.019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurdi, C.; Lelovics, V.; Hesszenberger, D.; Lajtai, A.; Lakatos, Á.; Herczeg, R.; Gödöny, K.; Mauchart, P.; Várnagy, Á.; Kovács, G.L.; et al. Amino Acid Profiling of Follicular Fluid in Assisted Reproduction Reveals Important Roles of Several Amino Acids in Patients with Insulin Resistance. Int. J. Mol. Sci. 2023, 24, 12458. https://doi.org/10.3390/ijms241512458
Kurdi C, Lelovics V, Hesszenberger D, Lajtai A, Lakatos Á, Herczeg R, Gödöny K, Mauchart P, Várnagy Á, Kovács GL, et al. Amino Acid Profiling of Follicular Fluid in Assisted Reproduction Reveals Important Roles of Several Amino Acids in Patients with Insulin Resistance. International Journal of Molecular Sciences. 2023; 24(15):12458. https://doi.org/10.3390/ijms241512458
Chicago/Turabian StyleKurdi, Csilla, Vanessza Lelovics, Dávid Hesszenberger, Anikó Lajtai, Ágnes Lakatos, Róbert Herczeg, Krisztina Gödöny, Péter Mauchart, Ákos Várnagy, Gábor L. Kovács, and et al. 2023. "Amino Acid Profiling of Follicular Fluid in Assisted Reproduction Reveals Important Roles of Several Amino Acids in Patients with Insulin Resistance" International Journal of Molecular Sciences 24, no. 15: 12458. https://doi.org/10.3390/ijms241512458
APA StyleKurdi, C., Lelovics, V., Hesszenberger, D., Lajtai, A., Lakatos, Á., Herczeg, R., Gödöny, K., Mauchart, P., Várnagy, Á., Kovács, G. L., & Kőszegi, T. (2023). Amino Acid Profiling of Follicular Fluid in Assisted Reproduction Reveals Important Roles of Several Amino Acids in Patients with Insulin Resistance. International Journal of Molecular Sciences, 24(15), 12458. https://doi.org/10.3390/ijms241512458