
Drug Development from Natural Products Based on the Pathogenic Mechanism of Asthma

 and
and 
Abstract
:1. Introduction
1.1. Definition
1.2. Epidemiology
2. Pathogenesis of Asthma
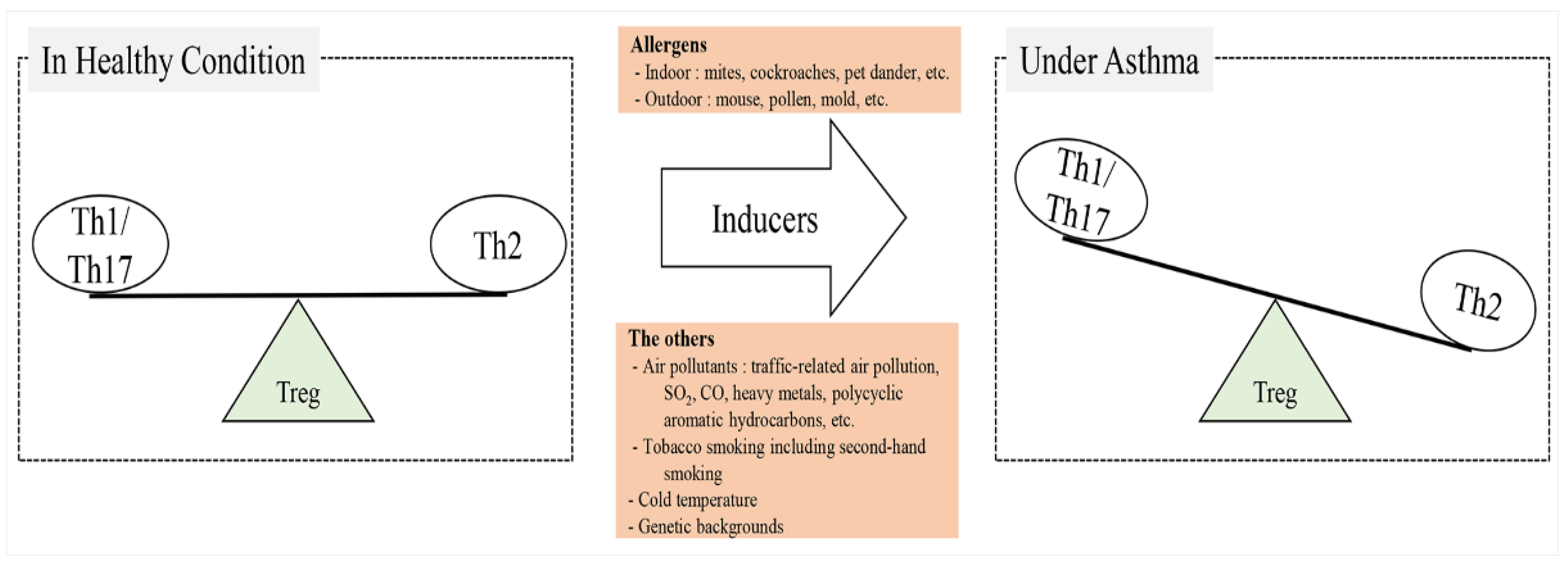
2.1. Asthma Inducers
2.2. Histopathological Changes in Asthma Occurrence
2.3. Mechanisms of Asthma Pathogenesis
2.3.1. Imbalance of Th1, Th17, and Th2
2.3.2. Inflammation in the Pulmonary System
2.3.3. Apoptosis/Cell Proliferation of Respiratory Epithelial Cells
3. Asthma Medications
3.1. Asthma Drug Classification
3.2. Adverse Effects of the Current Drugs

{kind=link}
{kind=link}
{kind=link}
| Classification | Drugs | Adverse Effects | Refs. | |
|---|---|---|---|---|
| Relievers (bronchodilators) | Anticholinergics | Atrovent, Tiotropium bromide, etc. | Dry mouth, constipation, cough, headache, nausea, etc. | [73] |
| β-adrenergic drugs | Salmeterol, Formoterol, etc. | Trembling, nervous tension, headaches, muscle cramps, heart attack, etc. | ||
| Methylxanthines | Theophylline, Aminophylline, etc. | Nausea/vomiting, diarrhea, palpitation, tachycardia, arrythmia, headaches, insomnia, etc. | ||
| Controllers | Anti-inflammatory drugs | Corticosteroids (Dexamethasone, Fluticasone, etc.) | Cataract/glaucoma, hypertension/hyperlipidemia, peptic ulcer/pancreatitis, myopathy/osteoporosis, dermal atrophy, immunological suppression, growth retardation, etc. | [74,75] |
| Leukotrienes modifiers (Montelukast, Zafirlukast, etc.) | Hyperactivity, excessive sleepiness, nyctophobia, nervousness, agitation, hallucination, sleep disorder, abdominal pain, rash, aphthous ulcer, appetite increase, headache, convulsion, etc. | [76] | ||
| Mast cell stabilizers (Cromolyn, Nedocromil, etc.) | Throat irritation, cough, anaphylaxis, headache, etc. | [77,78] | ||
| Immunomodulators | Glucocorticoids, 1,25-dihydroxy vitamin D3, etc. | Nausea, increment of serum aminotransferase, diarrhea, hair problem, etc. | [79] | |
4. Natural Drugs for Asthma Treatment
4.1. Immune Modulators
4.2. Anti-Inflammatory Effectors
4.3. Apoptosis Modulators
5. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Holgate, S.T. Pathogenesis of asthma. Clin. Exp. Allergy 2008, 38, 872–897. [Google Scholar] [CrossRef]
- 2022 GINA Main Report. Available online: https://ginasthma.org/wp-content/uploads/2022/07/GINA-Main-Report-2022-FINAL-22-07-01-WMS.pdf (accessed on 3 July 2023).
- Loachimescu, O.C.; Janocko, N.J.; Ciavatta, M.-M.; Howard, M.; Warnock, M.V. Obstructive lung disease and obstructive sleep apnea (OLDOSA) cohort study: 10-year assessment. J. Clin. Sleep Med. 2020, 16, 267–277. [Google Scholar] [CrossRef]
- WHO. Fact Sheet—Asthma. Available online: https://www.who.int/news-room/fact-sheets/detail/asthma (accessed on 3 July 2023).
- World Health Statistics 2021—Monitoring Health for the Sustainable Development Goals. Available online: https://apps.who.int/iris/bitstream/handle/10665/342703/9789240027053-eng.pdf?sequence=1&isAllowed=y (accessed on 3 July 2023).
- Ferrate, G.; Grutta, S.L. The burden of pediatric asthma. Front. Pediatr. 2018, 6, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, I.R.; Rosen, F.S. Allergy and allergic diseases. First of two parts. N. Eng. J. Med. 2001, 344, 30–37. [Google Scholar]
- Baos, S.; Calzada, D.; Cremades-Jimeno, L.; Sastre, J.; Picado, C.; Quiralte, J.; Florido, F.; Lahoz, C.; Cárdaba, B. Nonallergic asthma and its severity: Biomarkers for its discrimination in peripheral samples. Front. Immunol. 2018, 9, 1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platts-Mills, T.A.E.; Vervloet, D.; Thomas, W.R.; Aalberse, R.C.; Chapman, M.D. Indoor allergens and asthma: Report of the third international workshop. J. Allergy Clin. Immunol. 1997, 100 Pt 1, S1–S24. [Google Scholar]
- Baxi, S.N.; Phipatanakul, W. The role of allergen exposure and avoidance in asthma. Adolesc. Med. State. Art. Rev. 2010, 21, 57–71, viii–ix. [Google Scholar]
- Tiotiu, A.I.; Novakova, P.; Nedeva, D.; Chong-Neto, H.J.; Novakova, S.; Steiropoulos, P.; Kowal, K. Impact of air pollution on asthma outcomes. Int. J. Environ. Res. Public Health 2020, 17, 6212. [Google Scholar] [CrossRef] [PubMed]
- Bellou, V.; Gogali, A.; Kostikas, K. Asthma and tobacco smoking. J. Pers. Med. 2022, 12, 1231. [Google Scholar] [CrossRef]
- Hyrkas-Palmu, H.; Jaakkola, M.S.; Makikyro, E.M.S.; Jaakkola, J.J.K. Subtypes of asthma and cold weather-related respiratory symptoms. Int. J. Environ. Res. Public Health 2022, 19, 8790. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Zhang, Y.; Qiu, C. Gene polymorphisms in asthma: A narrative review. Ann. Transl. Med. 2022, 10, 711. [Google Scholar] [CrossRef] [PubMed]
- Omraninava, M.; Eslami, M.M.; Aslani, S.; Razi, B.; Imani, D.; Feyzinia, S. Interleukin 13 gene polymorphism and susceptibility to asthma: A meta-regression and meta-analysis. Eur. Ann. Allergy Clin. Immunol. 2022, 54, 150–167. [Google Scholar] [CrossRef]
- Huang, Q.P.; Xie, Z.F.; Huang, J. Assessment of the association between genetic polymorphisms in the CHI3L1 gene and asthma risk. Int. Arch. Allergy Immunol. 2022, 183, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Hough, K.P.; Curtiss, M.L.; Blain, T.J.; Liu, R.-M.; Trevor, J.; Deshane, J.S.; Thannickal, V.J. Airway remodeling in asthma. Front. Med. 2020, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Van den Bosch, W.B.; James, A.L.; Tiddens, H.A.W.M. Structure and function of small airways in asthma patients revised. Eur. Respir. Rev. 2021, 30, 200186. [Google Scholar] [CrossRef] [PubMed]
- Komlosi, Z.I.; van de Veen, W.; Kovacs, N.; Szucs, G.; Sokolowska, M.; O’Mahony, L.; Akdis, M.; Akdis, C.A. Cellular and molecular mechanisms of allergic asthma. Mol. Aspects Med. 2022, 85, 100995. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Kolls, J.K. Neutrophilic inflammation in asthma and association with disease severity. Trends Immunol. 2017, 38, 942–954. [Google Scholar] [CrossRef] [PubMed]
- Todd, I.; Spickett, G.; Faircolugh, L. Immunology Lecture Notes, 7th ed.; Wiley Blackwell: Pondicherry, India, 2015. [Google Scholar]
- Luo, W.; Hu, J.; Xu, W.; Dong, J. Distinct spatial and temporal roles for Th1, Th2, and Th17 cells in asthma. Front. Immunol. 2022, 13, 974066. [Google Scholar] [CrossRef]
- Boonpiyathad, T.; Sozener, Z.C.; Akdis, M.; Akdis, C.A. The role of Treg cell subsets in allergic disease. Asian Pac. J. Allergy 2020, 38, 139–149. [Google Scholar]
- Irvin, C.; Zafar, I.; Good, J.; Rollins, D.; Christianson, C.; Gorska, M.M.; Martin, R.J.; Alam, R. Increased frequency of dual-positive Th2/Th17 cells in bronchoalveolar lavage fluid characterizes a population of patients with severe asthma. J. Allergy Clin. Immunol. 2014, 134, 1175–1186. [Google Scholar] [CrossRef]
- Barnes, P.J. Targeting cytokines to treat asthma and chronic obstructive pulmonary disease. Nat. Rev. Immunol. 2018, 18, 454–466. [Google Scholar] [CrossRef]
- Kouro, T.; Takatsu, K. IL-5- and eosinophil-mediated inflammation: From discovery to therapy. Int. Immunol. 2009, 21, 1303–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The cytokines of asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef] [PubMed]
- Rincon, M.; Irvin, C.G. Role of IL-6 in asthma and other inflammatory pulmonary diseases. Int. J. Biol. Sci. 2012, 8, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Berry, M.; Brightling, C.; Pavord, I.; Wardlaw, A.J. TNF-α in asthma. Curr. Opin. Pharmacol. 2007, 7, 279–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romagnani, S. T-cell subsets (Th1 versus Th2). Ann. Allergy Asthma Immunol. 2000, 85, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Lighvani, A.A.; Frucht, D.M.; Jankovic, D.; Yamane, H.; Aliberti, J.; Hissong, B.D.; Nguyen, B.V.; Gadina, M.; Sher, A.; Paul, W.E.; et al. T-bet is rapidly induced by interferon-γ in lymphoid and myeloid cells. Proc. Natl. Acad. Sci. USA 2001, 98, 15137–15142. [Google Scholar] [CrossRef]
- Mitchell, C.; Provost, K.; Niu, N.; Homer, R.; Cohn, L. Interferon-gamma acts on the airway epithelium to inhibit local and systemic pathology in allergic airway disease. J. Immunol. 2001, 187, 3815–3820. [Google Scholar] [CrossRef] [Green Version]
- Chung, F. Anti-inflammatory cytokines in asthma and allergy: Interleukin-10, interleukin-12, interferon-γ. Mediat. Inflamm. 2001, 10, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Michaeloudes, C.; Abubakar-Waziri, H.; Lakhdar, R.; Raby, K.; Dixey, P.; Adcock, I.M.; Mumby, S.; Bhavsar, P.K.; Chung, K.F. Molecular mechanisms of oxidative stress in asthma. Mol. Aspects Med. 2022, 85, 101026. [Google Scholar] [CrossRef]
- Gall, S.J.; Tsal, M.; Pilliponsky, A. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Zuo, L.; Lucas, K.; Fortuna, C.A.; Chuang, C.-C.; Best, T.M. Molecular regulation of Toll-like receptors in asthma and COPD. Front. Physiol. 2015, 6, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, T.B. Early innate immunity to bacterial infection in the lung is regulated systemically by the commensal microbiota via Nod-like receptor ligands. Infect. Immun. 2014, 82, 4596–4606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.R. Apoptosis and the airway epithelium. J. Allergy 2011, 2011, 948406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.R.; Leff, A.R. Chapter 23. Epithelium as a target. In Inflammatory Mechanisms in Asthma; Holgate, S.T., Busse, W.W., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 1998; pp. 497–536. [Google Scholar]
- Bucchieri, F.; Puddicombe, S.M.; Lordan, J.L.; Richter, A.; Buchanan, D.; Wison, S.J.; Ward, J.; Zummo, G.; Howarth, P.H.; Djukanović, R.; et al. Asthmatic bronchial epithelium is more susceptible to oxidant-induced apoptosis. Am. J. Respir. Cell Mol. Biol. 2002, 27, 179–185. [Google Scholar] [CrossRef]
- Pelaia, G.; Cuda, G.; Vatrella, A.; Fratto, D.; Grembiale, R.D.; Tagliaferri, P.; Maselli, R.; Costanzo, F.S.; Marsico, S.A. Effects of transforming growth factor-β and budesonide on mitogen-activated protein kinase activation and apoptosis in airway epithelial cells. Am. J. Respir. Cell Mol. Biol. 2003, 29, 12–18. [Google Scholar] [CrossRef]
- Druilhe, A.; Wallaert, B.; Tsicopoulos, A.; Silva, J.-R.L.E.; Tillie-Leblond, I.; Tonnel, A.-B.; Pretolani, M. Apoptosis, proliferation, and expression of bcl-2, Fas, and Fas ligand in bronchial biopsies from asthmatics. Am. J. Respir. Cell Mol. Biol. 1998, 19, 747–757. [Google Scholar] [CrossRef]
- Benayoun, L.; Letuve, S.; Druilhe, A.; Boczkowski, J.; Dombret, M.C.; Mechighel, P.; Megret, J.; Leseche, G.; Aubier, M.; Pretolani, M. Regulation of peroxisome proliferator-activated receptor gamma expression in human asthmatic airways: Relationship with proliferation, apoptosis, and airway remodeling. Am. J. Respir. Crit. Care Med. 2001, 164, 1487–1494. [Google Scholar] [CrossRef]
- Penberthy, K.K.; Juncadella, I.J.; Ravichandran, K.S. Apoptosis and engulfment by bronchial epithelial cells. Implications for allergic airway inflammation. Ann. Am. Thorac. Soc. 2014, 11 (Suppl. S5), S259–S262. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Igney, F.H.; Krammer, P.H. Death and anti-death: Tumour resistance to apoptosis. Nat. Rev. Cancer 2002, 2, 277–288. [Google Scholar] [CrossRef]
- Saelens, X.; Festjens, N.; Walle, L.V.; van Grup, M.; van Loo, G.; Vandenabeele, P. Toxic proteins released from mitochondria in cell death. Oncogene 2004, 23, 2861–2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Chicheportiche, Y.; Bourdon, P.R.; Xu, H.; Hsu, Y.M.; Scott, H.; Hession, C.; Garcia, I.; Browning, J.L. TWEAK, a new secreted ligand in the tumor necrosis factor family that weakly induces apoptosis. J. Biol. Chem. 1997, 272, 32401–32410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef] [Green Version]
- Peter, M.E.; Kramer, P.H. Mechanisms of CD95 (APO-1/Fas)-mediated apoptosis. Curr. Opin. Immunol. 1998, 10, 545–551. [Google Scholar] [CrossRef]
- Suliman, A.; Datta, R.; Srivastava, R.K. Intracellular mechanisms of TRAIL:apoptosis through mitochondrial-dependent and –independent pathways. Oncogene 2001, 20, 2122–2133. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Moscardo, F.; Blesa, D.; Mestre, C.; Siebert, R.; Balasas, T.; Benito, A.; Rosenwald, A.; Climent, J.; Martinez, J.I.; Schilhabel, M.; et al. Characterization of 8p21.3 chromosomal deletions in B-cell lymphoma: TRAIL-R1and TRAIL-R2 as candidate dosage-dependent tumor suppressor genes. Blood 2005, 106, 3214–3222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapani, J.A.; Smyth, M.J. Functional significance of the perforin/granzyme cell death pathway. Nat. Rev. Immunol. 2002, 2, 735–747. [Google Scholar] [CrossRef]
- Pardo, J.; Bosque, A.; Brehm, R.; Wallich, R.; Naval, J.; Mullbacher, A.; Anel, A.; Simon, M.M. Apoptotic pathways are selectively activated by granzymne A and/or granzyme B in CTL-mediated target cell lysis. J. Cell. Biol. 2004, 167, 457–468. [Google Scholar] [CrossRef]
- Slee, E.A.; Adrain, C.; Martin, S.J. Executioner caspase-3, -6, and -7 perform distinct, non-redundant roles during the demolition phase of apoptosis. J. Biol. Chem. 2001, 276, 73020–77326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.W.; Kim, H.; Kim, K.H. Nuclear factor-κB regulates cyclooxygenase-2 expression and cell proliferation in human gastric cancer cells. Lab. Investig. 2001, 81, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuliga, M. NF-kappaB signaling in chronic inflammatory airway disease. Biomolecules 2015, 5, 1266–1283. [Google Scholar] [CrossRef] [Green Version]
- Nagai, H. Recent research and developmental strategy of anti-asthma drugs. Pharmacol. Ther. 2012, 133, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Sears, M.R.; Radner, F. Safety of budesonide/formoterol maintenance and reliever therapy in asthma trials. Respir. Med. 2009, 103, 1960–1968. [Google Scholar] [CrossRef] [Green Version]
- Matera, M.G.; Page, C.P.; Calzetta, L.; Rogliani, P.; Cazzola, M. Pharmacology and therapeutics of bronchodilators revisited. Pharmacol. Rev. 2020, 72, 218–252. [Google Scholar] [CrossRef]
- Cazzolar, M.; Page, C.P.; Rogliani, P.; Matera, M.G. β2-agonist therapy in lung disease. Am. J. Respir. Crit. Care Med. 2013, 187, 690–696. [Google Scholar] [CrossRef]
- Barnes, P.J. Theophylline. Am. J. Respir. Crit. Care Med. 2013, 188, 901–906. [Google Scholar] [CrossRef]
- Horita, N.; Miyazawa, N.; Kojima, R.; Inoue, M.; Ishigatsubo, Y.; Kaneko, T. Chronic use of theophylline and mortality in chronic obstructive pulmonary disease: A meta-analysis. Arch. Bronchneumol. 2016, 52, 233–238. [Google Scholar] [CrossRef]
- Debeuf, N.; Lambrecht, B.N. Eicosanoid control over antigen presenting cells in asthma. Front. Immunol. 2018, 9, 2006. [Google Scholar] [CrossRef] [Green Version]
- Hirata, F.; Stracke, M.L.; Schiffmann, E. Regulation of prostaglandin formation by glucocorticoids and their second messenger, lipocortins. J. Steroid Biochem. 1987, 27, 1053–1056. [Google Scholar] [CrossRef]
- Dahlen, S.E. Treatment of asthma with antileukotrienes: First line or last resort therapy? Eur. J. Pharmacol. 2006, 533, 40–56. [Google Scholar] [CrossRef]
- Holgate, S.T.; Peters-Golden, M.; Panettieri, R.A.; Henderson, W.R. Roles of cysteinyl leukotrienes in airway inflammation, smooth muscle function, and remodeling. J. Allergy Clin. Immunol. 2003, 111 (Suppl. S1), S18–S36. [Google Scholar] [CrossRef] [PubMed]
- Finn, D.F.; Walsh, J.J. Twenty-first century mast cell stabilizers. Br. J. Pharmacol. 2013, 170, 23–37. [Google Scholar] [CrossRef] [Green Version]
- Oka, T.; Kalesnikoff, J.; Starkl, P.; Tsai, M.; Galli, S.J. Evidence questioning cromolyn’s effectiveness and selectivity as a ‘mast cell stabilizer’ in mice. Lab. Investig. 2012, 92, 1472–1482. [Google Scholar] [CrossRef] [Green Version]
- Chandel, A.; Goyal, A.K.; Ghosh, G.; Rath, G. Recent advances in aerosolized drug delivery. Biomed. Pharmacother. 2019, 112, 108601. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Gelfand, A.; Mathew, M.S.; Atem, F.D.; Srikanth, N.; Delclos, G.L. Adverse drug events related to common asthma medications in US hospitalized children, 2000–2016. Drugs Real World Outcomes 2022, 9, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Side Effects—Bronchodilators. Available online: https://www.nhs.uk/conditions/bronchodilators/side-effects/ (accessed on 3 July 2023).
- Ciriaco, M.; Ventrice, P.; Russo, G.; Scicchitano, M.; Mazzitello, G.; Scicchitano, F.; Russo, E. Corticosteroid-related central nervous system side effects. J. Pharmacol. Pharmther. 2013, 4 (Suppl. S1), S94–S98. [Google Scholar] [CrossRef] [Green Version]
- Wise, J. Corticosteroids for asthma may suppress growth in children in first year of treatment, researchers say. BMJ 2014, 349, g4623. [Google Scholar] [CrossRef]
- Erdem, S.B.; Nacaroglu, H.T.; Karkiner, C.S.U.; Gunay, I.; Can, D. Side effects of leukotriene receptor antagonists in asthmatic children. Iran J. Pediatr. 2015, 25, e3313. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, G.G.; Konig, P. Cromolyn sodium: A review. Pharmacotherapy 1985, 5, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Bielory, L.; Lien, K.W.; Bigelsen, S. Efficacy and tolerability of newer antihistamines in the treatment of allergic conjunctivitis. Drugs 2005, 65, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.J.; Segars, L.W. Methotrexate for the treatment of chronic corticosteroid-dependent asthma. Clin. Pharm. 1993, 12, 762–767. [Google Scholar] [PubMed]
- Yao, J.; Jiang, M.; Zhang, Y.; Liu, X.; Du, Q.; Feng, G. Chrysin alleviates allergic inflammation and airway remodeling in a murine model of chronic asthma. Int. Immunopharmacol. 2016, 32, 24–31. [Google Scholar] [CrossRef]
- Seo, J.-W.; Cho, S.-C.; Park, S.-J.; Lee, E.-J.; Lee, J.-H.; Han, S.-S.; Pyo, B.S.; Park, D.-H.; Kim, B.-H. 1′-acetoxychavicol acetate isolated from Alpinia galanga ameliorates ovalbumin-induced asthma in mice. PLoS ONE 2013, 8, e56447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, M.-A.; Seo, J.-H.; Seo, J.-W.; Jo, G.H.; Jung, S.K.; Yu, R.; Park, D.-H.; Park, S.-J. Bacillus subtilis KCTC 11782BP-produced alginate oligosaccharide effectively suppresses asthma via T-helper cell type 2-related cytokines. PLoS ONE 2015, 10, e0117524. [Google Scholar] [CrossRef]
- Oliveira, T.T.; Campos, K.M.; Cerqueira-Lima, A.T.; Cana Brasil Carneiro, T.; da Silva Velozo, E.; Ribeiro Melo, I.C.A.; Figueiredo, E.A.; de Jesus Oliveira, E.; de Vasconcelos, D.F.S.A.; Pontes-de-Carvalho, L.C.; et al. Potential therapeutic effect of Allium cepa L. and quercetin in a murine model of Blomia tropicalis induced asthma. Daru 2015, 23, 18. [Google Scholar] [CrossRef]
- Bok, S.H.; Seo, J.H.; Bae, C.S.; Kang, B.; Cho, S.; Park, D.H. Allium hookeri root extract regulates asthmatic changes through immunological modulation of Th1/Th2-related factors in an ovalbumin-induced asthma mouse model. Mol. Med. Rep. 2019, 20, 3215–3223. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.C.; Hsiao, H.B.; Lin, W.C. A standardized aqueous extract of Anoectochilus formosanus modulated airway hyperresponsiveness in an OVA-inhaled murine model. Phytomedicine 2010, 17, 557–562. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, Y.; Li, C.; Jiang, Y.; He, L.; Guo, W.; Guo, M.; Gong, W. Immunoregulation effect of crude extract of C. elegans on allergic asthma. Int. J. Clin. Exp. Med. 2014, 7, 886–892. [Google Scholar]
- Lee, S.-Y.; Bae, C.-S.; Seo, N.-S.; Na, C.-S.; Yoo, H.Y.; Oh, D.-S.; Bae, M.-S.; Kwon, M.-S.; Cho, S.-S.; Park, D.-H. Camellia japonica oil suppressed asthma occurrence via GATA-3 & IL-4 pathway and its effective and major component is oleic acid. Phytomedicine 2019, 57, 84–94. [Google Scholar] [PubMed]
- Bui, T.T.; Piao, C.H.; Kim, S.M.; Song, C.H.; Shin, H.S.; Lee, C.-H.; Chai, O.H. Citrus tachibana leaves ethanol extract alleviates airway inflammation by the modulation of Th1/Th2 imbalance via inhibiting NF-κB signaling and histamine secretion in a mouse model of allergic asthma. J. Med. Food 2017, 20, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Chong, L.; Zhang, W.; Nie, Y.; Yu, G.; Liu, L.; Lin, L.; Wen, S.; Zhu, L.; Li, C. Protective effect of curcumin on acute airway inflammation of allergic asthma in mice through h Notch1-GATA3 signaling pathway. Inflammation 2014, 37, 1476–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.H.; Bang, M.A.; Kim, G.; Cho, S.S.; Park, D.H. Erythronium japonicum attenuates histopathological lung abnormalities in a mouse model of ovalbumin-induced asthma. Int. J. Mol. Med. 2016, 37, 1221–1228. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-Y.; Kim, M.-H.; Kim, S.-H.; Ahn, T.; Kim, S.-W.; Kwak, Y.-S.; Cho, I.-H.; Nah, S.-Y.; Cho, S.-S.; Park, K.M.; et al. Korea red ginseng affects ovalbumin-induced asthma by modulating IL-12, IL-4, and IL-6 levels and the NF-κB/COX-2 and PGE2 pathways. J. Ginseng Res. 2021, 45, 482–489. [Google Scholar] [CrossRef]
- Lee, S.Y.; Bae, C.S.; Seo, J.H.; Cho, S.S.; Bae, M.S.; Oh, D.S.; Park, D.H. Mycoleptodonoides aitchisonii suppresses asthma via Th2 and Th1 cell regulation in an ovalbumin-induced asthma mouse model. Mol. Med. Rep. 2018, 17, 11–20. [Google Scholar]
- Lee, S.Y.; Bae, C.S.; Choi, Y.-h.; Seo, N.S.; Na, C.S.; Yoo, J.C.; Cho, S.S.; Park, D.H. Opuntia humifusa modulates morphological changes characteristic of asthma via IL-4 and IL-13 in an asthma murine model. Food Nutr. Res. 2017, 61, 1393307. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.W.; Lee, S.U.; Lee, S.; Song, H.-H.; Son, T.H.; Kim, Y.-U.; Yuk, H.J.; Ro, H.; Lee, C.-K.; Hong, S.-T.; et al. 3-methoxy-catalposide inhibits inflammatory effects in lipopolysaccharide-stimulated RAW 264.7 macrophages. Cytokines 2017, 91, 57–64. [Google Scholar] [CrossRef]
- Kang, H.J.; Jeong, J.S.; Park, N.J.; Go, G.B.; Kim, S.O.; Park, C.; Kim, B.W.; Hong, S.Y.; Choi, Y.H. An ethanol extract of Aster yomena (Kitam.) Honda inhibits lipopolysaccharide-induced inflammatory response in murine RAW 264.7 macrophages. Biosci. Trends 2017, 11, 85–94. [Google Scholar] [CrossRef]
- Bok, S.H.; Han, K.M.; Boo, H.O.; Cho, S.S.; Park, D.H. Codonopsis laceolata water extract ameliorates asthma severity by inducing Th2 cells’and pulmononary epithelial cells’ apoptosis via NF-κB/COX-2 pathway. Processes 2022, 10, 1249. [Google Scholar] [CrossRef]
- Rosa, S.I.G.; Rios-Santos, F.; Balogun, S.O.; de Almeida, D.A.T.; Damazo, A.S.; da Cruz, T.C.D.; Pavan, E.; Barbosa, R.d.S.; Alvim, T.d.C.; Soares, I.M.; et al. Hydroethanolic extract from Echinodorus scaber Rataj leaves inhibits inflammation in ovalbumin-induced allergic asthma. J. Ethnopharmacol. 2017, 203, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Shipton, F.N.; Khoo, T.J.; Hossan, M.S.; Wiart, C. Activity of Pericampylus glaucus and periglaucine A in vitro against nasopharangeal carcinoma and anti-inflammatory activity. J. Ethnopharmacol. 2017, 198, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Shin, I.S.; Shin, N.R.; Jeon, C.M.; Hong, J.M.; Kwon, O.K.; Kim, J.C.; Oh, S.P.; Hahn, K.W.; Ahn, K.S. Inhibitory effects of Pycnogeneol® (French maritime pine bark extract) on airway inflammation in ovalbumin-induced allergic asthma. Food Chem. Toxicol. 2013, 62, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Lee, S.Y.; Kim, M.; Park, S.; Park, J.; Kwon, Y.; Park, D.H. Saururus chinensis-controlled allergic pulmonary disease through NF-κB/COX-2 and PGE2 pathways. PeerJ 2020, 8, e10043. [Google Scholar] [CrossRef]
- Yao, J.; Zhang, Y.S.; Feng, G.Z.; Du, Q. Chrysin inhibits human airway smooth muscle cell proliferation through the extracellular signal-regulated kinase 1/2 signaling pathway. Mol. Med. Rep. 2015, 12, 7693–7698. [Google Scholar] [CrossRef]
- Zheng, M.; Zhang, Q.; Joe, Y.; Lee, B.H.; Ryu, D.G.; Kwon, K.B.; Ryter, S.W.; Chung, H.T. Curcumin induces apoptotic cell death of activated human CD4+ T cells via increasing endoplasmic reticulum stress and mitochondrial dysfunction. Int. Immunopharmacol. 2013, 15, 517–523. [Google Scholar] [CrossRef]
- Orru, H.; Ebi, K.L.; Forsberg, B. The interplay of climate change and air pollution on health. Curr. Envir. Health Rep. 2017, 4, 504–513. [Google Scholar] [CrossRef]
- Guryanova, S.V.; Gigani, O.B.; Gudima, G.O.; Kataeva, A.M.; Kolesnikova, N.V. Dual effect of low-molecular-weight bioregulators of bacterial origin in experimental model of asthma. Life 2022, 12, 192. [Google Scholar] [CrossRef]

 , inducer;
, inducer;  , neutrophil;
, neutrophil;  , eosinophil;
, eosinophil;  , basophil;
, basophil;  , Goblet cell.
, inducer; , neutrophil; , eosinophil; , basophil; , Goblet cell.
, Goblet cell.
, inducer; , neutrophil; , eosinophil; , basophil; , Goblet cell. , apoptotic ligand;
, apoptotic ligand;  , apoptotic receptor;
, apoptotic receptor;  , perforin;
, perforin;  , granzyme.
, apoptotic ligand; , apoptotic receptor; , perforin; , granzyme.
, granzyme.
, apoptotic ligand; , apoptotic receptor; , perforin; , granzyme.
| Stage | Acute | Chronic | |
|---|---|---|---|
| Phase | Early | Late | |
| Onset Time after Allergens Contact | Several minutes | Onset time: 2~6 h Peak time: 6~9 h | A few months to years |
| Physiological Symptoms | Vasodilation, vascular permeability, bronchoconstriction, mucous hypersecretion, etc. | Wheeze, shortness of breath, cough | Wheeze, shortness of breath, cough, mucus hypersecretion, sleep apnea to death |
| Causes of the Symptoms | Cytokine and chemokine release after the cross linkage of IgE–mast cell/basophil–allergens | Rise of Th2-related cytokines and increment of WBCs | Alteration of the extracellular matrix and of the constitutive cells of the affected organ |
| Classification | Source | Applied Type/ Active Substance | Study | Dose (Max) & Route (mg/kg) | Mode of Action | Refs. | |
|---|---|---|---|---|---|---|---|
| Immune modulators | Chrysin | Propolis, passionflower, etc. | Isolated compound/ chrysin | Animal | 100, P.O. | Decreasing the levels of IgE, IL-4, and IL-13 | [80] |
| 1′-acetoxychavicol acetate | Alpinia galanga | Isolated compound/ 1′-acetoxychavicol acetate | Animal | 50, P.O. | Decreasing the levels of Th1 cell-related cytokines such as IL-12 and IFN-γ and of Th2-related cytokines such as IL-4, IL-5, and IL-13 | [81] | |
| Alginate oligosaccharide | Fermented alginate | Fermented form | Animal | 400, P.O. | Inhibiting the releases of IL-15 and IL-13 | [82] | |
| Allium cepa L. and quercetin | Allium cepa L. and quercetin | Methanol extract and isolated compound/quercetin | Cell & Animal | 1000 + 15 μg/mL, 1000 + 30 mg/kg, P.O. | Decreasing the levels of IL-4 and IL-13 | [83] | |
| Allium hookeri | Allium hookeri | Ethanol extract | Animal | 300, P.O. | Inhibiting the releases of IL-4, IL-5, and IL-13 | [84] | |
| Anoectochilus formosanus HAYATA | Anoectochilus formosanus HAYATA | Water extract | Animal | 1000, P.O. | Decreasing the levels of IgE, IL-4, and IL-5 | [85] | |
| Caenorhabditis elegans | Caenorhabditis elegans | Crude extract | Animal | 50 μg/head | Decreasing the levels of IgE, IL-5, and IL-13 but increasing the levels of IFN-γ | [86] | |
| Camellia japonica | Camellia japonica | Extract oil and isolated compound/oleic acid | Animal | 500, P.O. | Decreasing the levels of IL-4 via GATA-3 inactivation | [87] | |
| Citrus tachibana | Citrus tachibana | Ethanol extract narirutin, hesperidin | Animal | 400, P.O. | Decreasing the levels of IL-4 and IL-5 but increasing the levels of IL-12 and IFN-γ | [88] | |
| Curcumin | Curcuma longa | Isolated compound/diferuloylmethane | Animal | 2000, P.O. | Inactivating Th2 cell transcription factor, GATA-3 | [89] | |
| Erythronium japonicum | Erythronium japonicum | Ethanol extract chlorogenic acid | Animal | 600, P.O. | Decreasing the levels of IL-12p35, IFN-γ, IL-4, IL-5, and IL-13 | [90] | |
| Korean red ginseng | Panax ginseng | Water extract Active substances: Rb1, Rg1 | Animal | 50, P.O. | Decreasing the levels of IL-12, IL-4, and IL-6 | [91] | |
| Mycoleptodonoides aitchisonii | Mycoleptodonoides aitchisonii | Water extract niacin, oleic acid, linoleic acid | Animal | 1000, P.O. | Decreasing the levels of IL-4, IL-5, and IL-13 | [92] | |
| Opuntia humifusa | Opuntia humifusa | Ethanol extract rutin, quercetin | Animal | 500, P.O. | Decreasing the levels of IL-12, IFN-γ, IL-4, IL-13, IL-6, and TNF-α | [93] | |
| Anti-inflammatory effectors | Chrysin | Propolis, passionflower, etc. | Isolated compound/chrysin | Animal | 100, P.O. | Blocking inflammatory cell infiltration | [80] |
| 3-methoxy-catalposide | Psueolysimachion rotundum var. subintegrum | Isolated compound/3-methoxy-catalposide | Cell | 20 μM | Decreasing the expression of COX-2 and iNOS and downregulating the levels of IL-1β, IL-6, and TNF-α | [94] | |
| Aster yomensa (Kitam.) Honda | Aster yomensa (Kitam.) Honda | Ethanol extract | Cell | 300 ng/mL | Decreasing the levels of NO and IL-1β | [95] | |
| Codonopsis laceolata | Codonopsis laceolata | Water extract Active substance: lobetyolin | Animal | 300, P.O. | Blocking the NF-κB/COX-2 and PGE2 pathway | [96] | |
| Echinodorus scaber Rataj | Echinodorus scaber Rataj | Ethanol extract | Animal | 30, P.O. | Suppressing inflammatory cells’ migration | [97] | |
| Korean red ginseng | Panax ginseng | Water extract Active substances: Rb1, Rg1 | Animal | 50, P.O. | Blocking the NF-κB/COX-2 and PGE2 pathway | [91] | |
| Pericampylus glaucus | Pericampylus glaucus | Hexane, chloroform or ethanol extract | Cell | 250 μg/mL | Blocking the synthesis of COX-1 and COX-2 | [98] | |
| Pinus maritime | Pinus maritime | Water extract | Cell | 100 pg/mL | Enhancing the HO-1 antioxidative system and decreasing the levels of IL-1β and IL-6 | [99] | |
| Saururus chinenesis | Saururus chinenesis | Water extract rutin, quercitrin, quercetin | Animal | 300, P.O. | Blocking the NF-κB/COX-2 and PGE2 pathway | [100] | |
| Apoptosis modulators | Chrysin | Propolis, passionflower, etc. | Isolated compound/chrysin | Animal/Cell | 100, P.O./40 μM | Inducing the apoptosis of airway smooth muscle cells | [80,101] |
| Codonopsis laceolata | Codonopsis laceolata | Water extract Active substance: lobetyolin | Animal | 300, P.O. | Inducing infiltrated inflammatory cells’ apoptosis and pulmonary epithelial cells’ death | [96] | |
| Curcumin | Curcuma longa | Isolated compound/diferuloylmethane | Cell | 20 μM | Decreasing the expression of the anti-apoptotic protein Bcl-2 and inducing endoplasmic reticulum stress | [102] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-H.; Bae, C.-S.; Bok, S.-H.; Choi, H.-S.; Ahn, T.; Cho, S.-S.; Park, D.-H. Drug Development from Natural Products Based on the Pathogenic Mechanism of Asthma. Int. J. Mol. Sci. 2023, 24, 12469. https://doi.org/10.3390/ijms241512469
Kim M-H, Bae C-S, Bok S-H, Choi H-S, Ahn T, Cho S-S, Park D-H. Drug Development from Natural Products Based on the Pathogenic Mechanism of Asthma. International Journal of Molecular Sciences. 2023; 24(15):12469. https://doi.org/10.3390/ijms241512469
Chicago/Turabian StyleKim, Min-Hee, Chun-Sik Bae, So-Hyeon Bok, Hyo-Seung Choi, Taeho Ahn, Seung-Sik Cho, and Dae-Hun Park. 2023. "Drug Development from Natural Products Based on the Pathogenic Mechanism of Asthma" International Journal of Molecular Sciences 24, no. 15: 12469. https://doi.org/10.3390/ijms241512469
APA StyleKim, M. -H., Bae, C. -S., Bok, S. -H., Choi, H. -S., Ahn, T., Cho, S. -S., & Park, D. -H. (2023). Drug Development from Natural Products Based on the Pathogenic Mechanism of Asthma. International Journal of Molecular Sciences, 24(15), 12469. https://doi.org/10.3390/ijms241512469