


Dual Adeno-Associated Virus 9 with Codon-Optimized DYSF Gene Promotes In Vivo Muscle Regeneration and May Decrease Inflammatory Response in Limb Girdle Muscular Dystrophy Type R2

 , ,
, ,
Abstract
:1. Introduction
2. Results
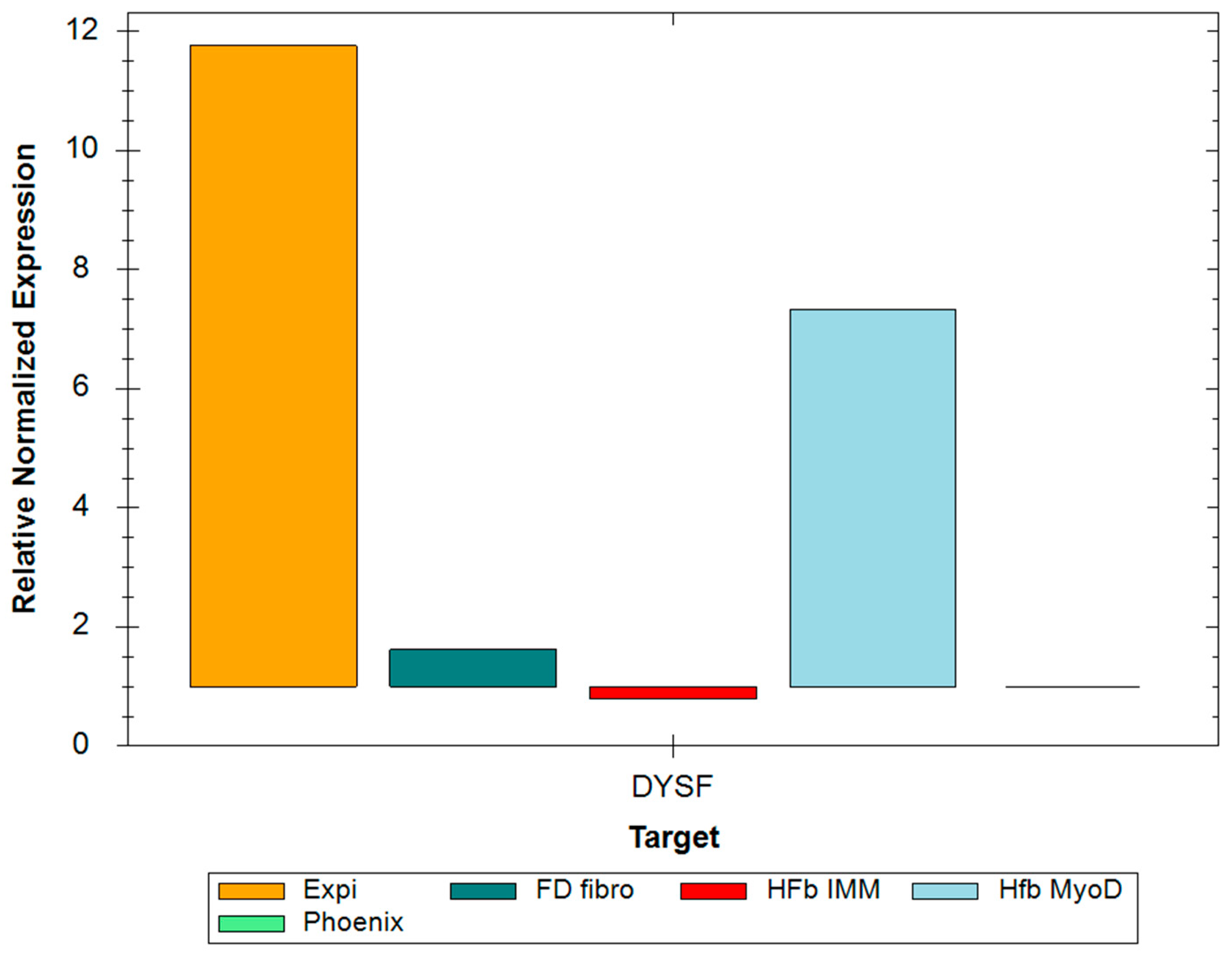
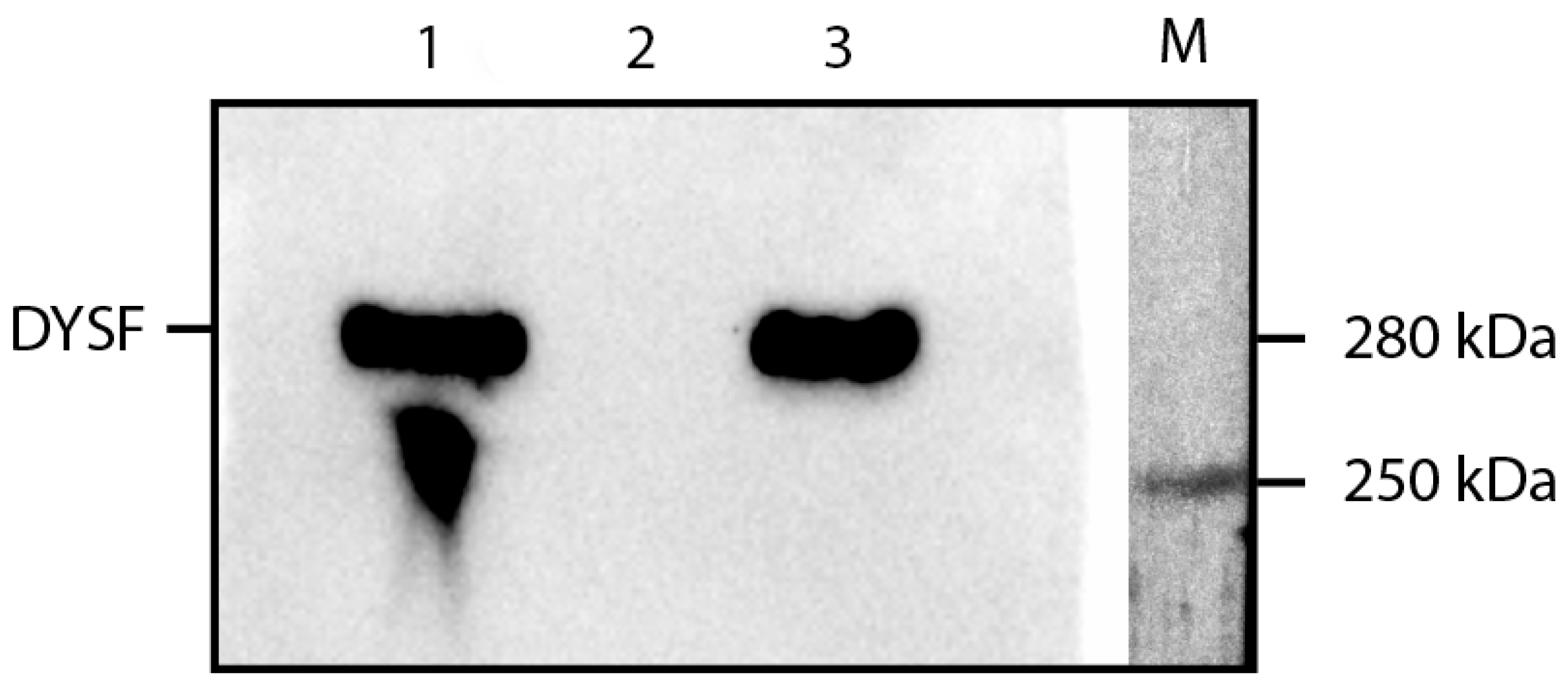
2.1. RT-PCR and Western Blot
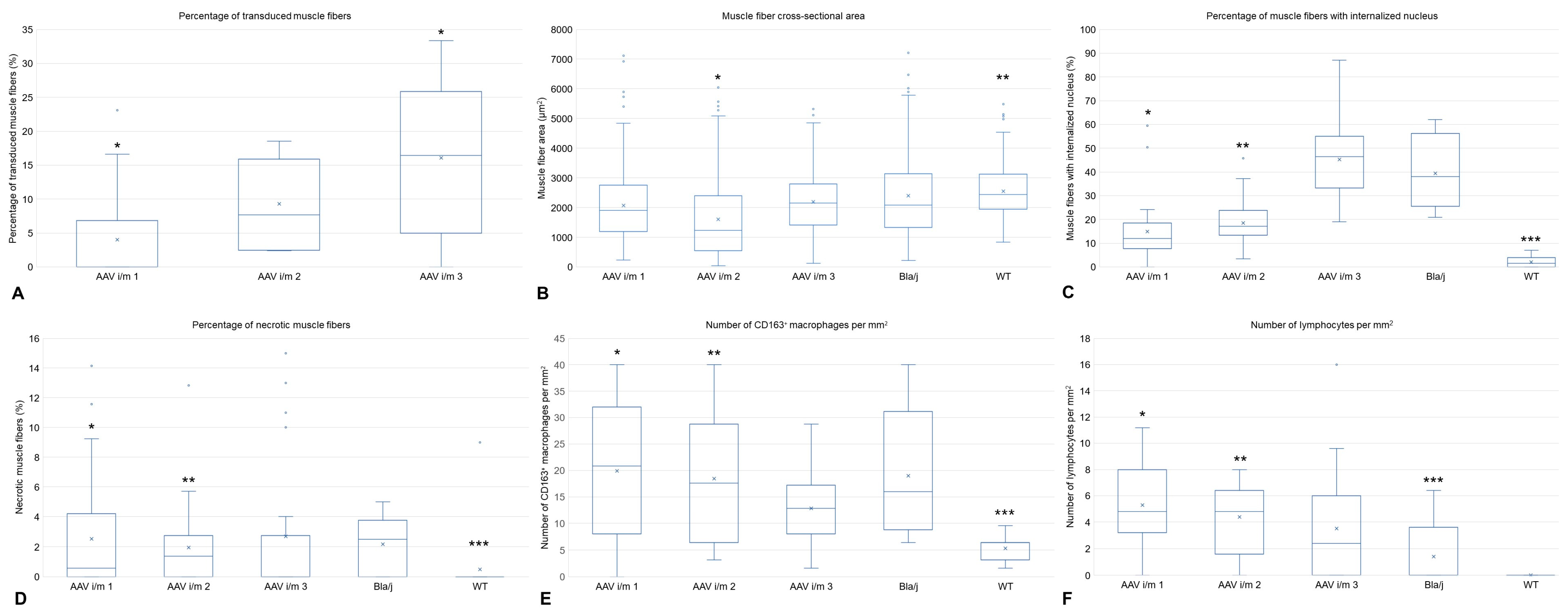
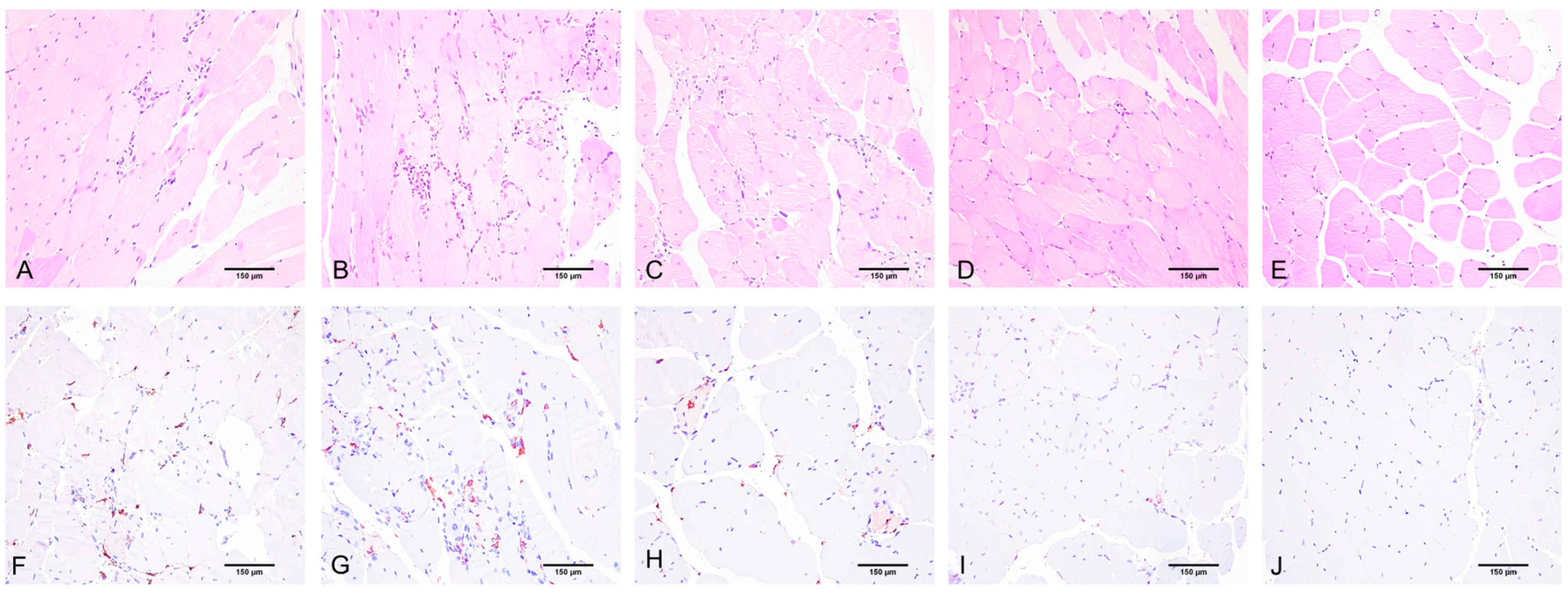
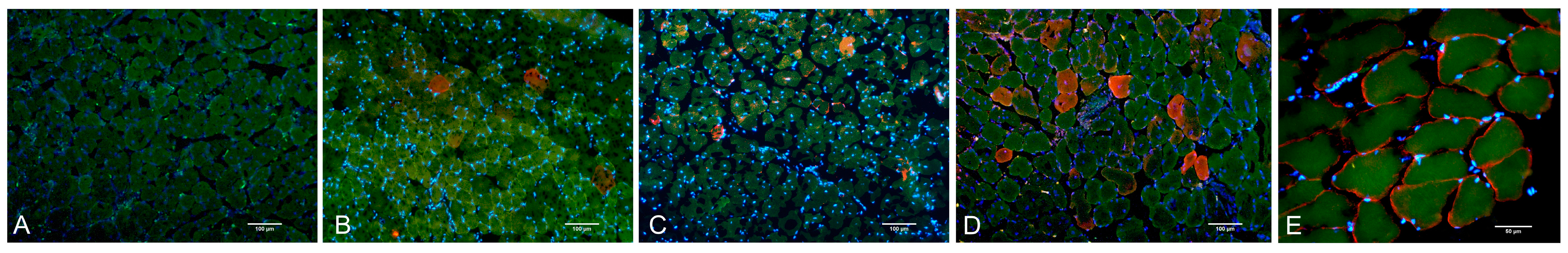
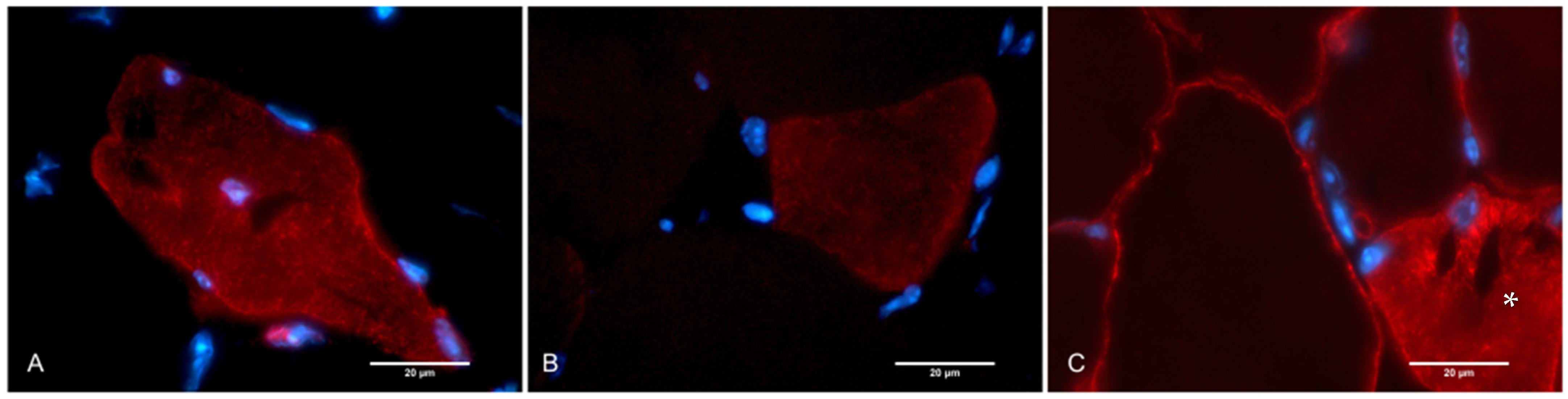
2.2. Histological Assay, Morphometry, and IHC
3. Discussion
4. Materials and Methods
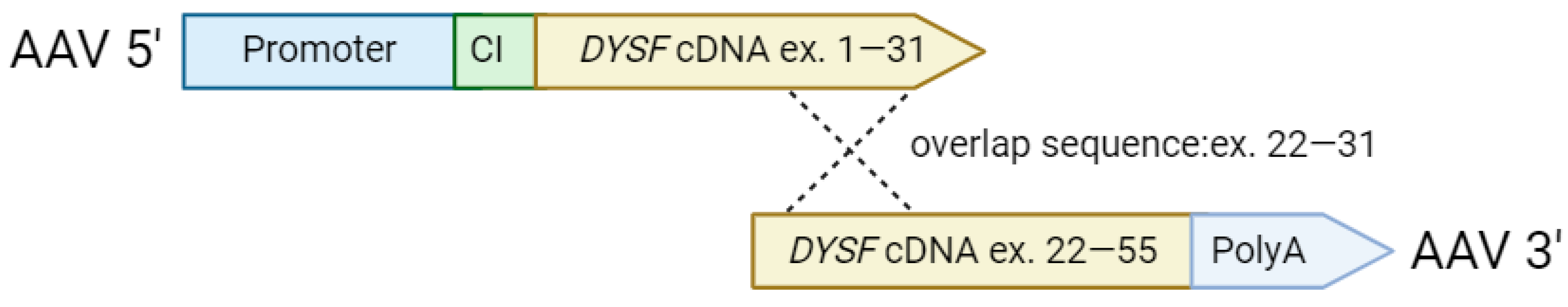
4.1. Construction and Production of Recombinant AAV
4.2. Construction and Production of Recombinant Lentivirus
4.3. Fibroblast Preparation
4.4. Myogenic Transdifferentiation
4.5. Animal Models
4.6. PCR
4.7. Western Blot
4.8. Histological Assay, Morphometry, and IHC
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Urtizberea, J.A.; Bassez, G.; Leturcq, F.; Nguyen, K.; Krahn, M.; Levy, N. Dysferlinopathies. Neurol. India 2008, 56, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Aoki, M.; Illa, I.; Wu, C.; Fardeau, M.; Angelini, C.; Serrano, C.; Urtizberea, J.A.; Hentati, F.; Hamida, M.B.; et al. Dysferlin, a novel skeletal muscle gene, is mutated in Miyoshi myopathy and limb girdle muscular dystrophy. Nat Genet. 1998, 20, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Rawat, R.; Cohen, T.V.; Ampong, B.; Francia, D.; Henriques-Pons, A.; Hoffman, E.P.; Nagaraju, K. Inflammasome Up-Regulation and Activation in Dysferlin-Deficient Skeletal Muscle. Am. J. Pathol. 2010, 176, 2891–2900. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, I.; Brengman, J.M.; Engel, A.G. Analysis of cytokine expression in muscle in inflammatory myopathies, Duchenne dystrophy, and non-weak controls. J. Neuroimmunol. 1995, 63, 9–16. [Google Scholar] [CrossRef]
- Lundberg, I.; Ulfgren, A.-K.; Nyberg, P.; Andersson, U.; Klareskog, L. Cytokine production in muscle tissue of patients with idiopathic inflammatory myopathies. Arthritis Rheum. 1997, 40, 865–874. [Google Scholar] [CrossRef]
- Hofhuis, J.; Bersch, K.; Büssenschütt, R.; Drzymalski, M.; Liebetanz, D.; Nikolaev, V.O.; Wagner, S.; Maier, L.S.; Gärtner, J.; Klinge, L.; et al. Dysferlin mediates membrane tubulation and links T-tubule biogenesis to muscular dystrophy. J. Cell Sci. 2017, 130, 841–852. [Google Scholar] [CrossRef]
- Zorin, V.L.; Zorina, A.I.; Eremin, I.I.; Deev, R.V.; Kopnin, P.B.; Volozhin, G.A.; Pulin, A.A. Gingiva as a source of stromal cells with high differentiating and reparative potential. Genes Cells 2017, 12, 37–51. [Google Scholar] [CrossRef]
- Pryadkina, M.; Lostal, W.; Bourg, N.; Charton, K.; Roudaut, C.; Hirsch, M.L.; Richard, I. A comparison of AAV strategies distinguishes overlapping vectors for efficient systemic delivery of the 6.2 kb Dysferlin coding sequence. Mol. Ther. Methods Clin. Dev. 2015, 2, 15009. [Google Scholar] [CrossRef]
- Llanga, T.; Nagy, N.; Conatser, L.; Dial, C.; Sutton, R.B.; Hirsch, M.L. Structure-Based Designed Nano-Dysferlin Significantly Improves Dysferlinopathy in BLA/J Mice. Mol. Ther. 2017, 25, 2150–2162. [Google Scholar] [CrossRef]
- Grose, W.E.; Clark, K.R.; Griffin, D.; Malik, V.; Shontz, K.M.; Montgomery, C.L.; Lewis, S.; Brown, R.H., Jr.; Janssen, P.M.; Mendell, J.R.; et al. Homologous recombination mediates functional recovery of dysferlin deficiency following AAV5 gene transfer. PLoS ONE 2012, 7, e39233. [Google Scholar] [CrossRef]
- Lostal, W.; Bartoli, M.; Bourg, N.; Roudaut, C.; Bentaïb, A.; Miyake, K.; Guerchet, N.; Fougerousse, F.; McNeil, P.; Richard, I. Efficient recovery of dysferlin deficiency by dual adeno-associated vector-mediated gene transfer. Hum. Mol. Genet. 2010, 19, 1897–1907. [Google Scholar] [CrossRef] [PubMed]
- Rigamonti, E.; Zordan, P.; Sciorati, C.; Rovere-Querini, P.; Brunelli, S. Macrophage plasticity in skeletal muscle repair. BioMed Res. Int. 2014, 2014, 560629. [Google Scholar] [CrossRef] [PubMed]
- Cohen, T.V.; Cohen, J.E.; Partridge, T.A. Myogenesis in dysferlin-deficient myoblasts is inhibited by an intrinsic inflammatory response. Neuromuscul. Disord. 2012, 22, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G.; Welc, S.S.; Wehling-Henricks, M. Immunobiology of Inherited Muscular Dystrophies. Compr. Physiol. 2018, 8, 1313–1356. [Google Scholar] [CrossRef]
- Klinge, L.; Harris, J.; Sewry, C.; Charlton, R.; Anderson, L.; Laval, S.; Chiu, Y.H.; Hornsey, M.; Straub, V.; Barresi, R.; et al. Dysferlin associates with the developing T-tubule system in rodent and human skeletal muscle. Muscle Nerve 2010, 41, 166–173. [Google Scholar] [CrossRef]
- Ampong, B.N.; Imamura, M.; Matsumiya, T.; Yoshida, M.; Takeda, S.I. Intracellular localization of dysferlin and its association with the dihydropyridine receptor. Acta Myol. 2005, 24, 134–144. [Google Scholar]
- Barefield, D.Y.; Sell, J.J.; Tahtah, I.; Kearns, S.D.; McNally, E.M.; Demonbreun, A.R. Loss of dysferlin or myoferlin results in differential defects in excitation-contraction coupling in mouse skeletal muscle. Sci. Rep. 2021, 11, 15865. [Google Scholar] [CrossRef]
- Kerr, J.P.; Ziman, A.P.; Mueller, A.L.; Muriel, J.M.; Kleinhans-Welte, E.; Gumerson, J.D.; Vogel, S.S.; Ward, C.W.; Roche, J.A.; Bloch, R.J. Dysferlin stabilizes stress-induced Ca2+ signaling in the transverse tubule membrane. Proc. Natl. Acad. Sci. USA 2013, 110, 20831–20836. [Google Scholar] [CrossRef]
- Bansal, D.; Campbell, K.P. Dysferlin and the plasma membrane repair in muscular dystrophy. Trends Cell Biol. 2004, 14, 206–213. [Google Scholar] [CrossRef]
- de Luna, N.; Gallardo, E.; Illa, I. In Vivo and In Vitro Dysferlin Expression in Human Muscle Satellite Cells. J. Neuropathol. Exp. Neurol. 2004, 63, 1104–1113. [Google Scholar] [CrossRef]
- Piccolo, F.; Moore, S.A.; Ford, G.C.; Campbell, K.P. Intracellular accumulation and reduced sarcolemmal expression of dysferlin in limb–girdle muscular dystrophies. Ann. Neurol. 2000, 48, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Starostina, I.G.; Solovyeva, V.V.; Shevchenko, K.G.; Deev, R.V.; Isaev, A.A.; Rizvanov, A.A. Formation of the recombinant adenovirus encoding codon-optimized dysferlin gene and analysis of the recombinant protein expression in cell culture in vitro. Genes Cells 2012, 7, 25–28. [Google Scholar]
- Mavlikeev, M.O.; Kiyasov, A.P.; Deev, R.V. Histological technique in the pathomorphological laboratory: Educational and methodological manual (Gistologicheskaya Tekhnika v Patomorfologicheskoj Laboratorii: Uchebno-Metodicheskoe Posobie); Prakticheskaya Medicina: Moscow, Russia, 2023; p. 123. [Google Scholar]
- Chernova, O.N.; Chekmareva, I.A.; Mavlikeev, M.O.; Yakovlev, I.A.; Kiyasov, A.P.; Deev, R.V. Structural and ultrastructural changes in the skeletal muscles of dysferlin-deficient mice during postnatal ontogenesis. Ultrastruct. Pathol. 2022, 46, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Sondergaard, P.C.; Griffin, D.A.; Pozsgai, E.R.; Johnson, R.W.; Grose, W.E.; Heller, K.N.; Shontz, K.M.; Montgomery, C.L.; Liu, J.; Clark, K.R.; et al. AAV.Dysferlin Overlap Vectors Restore Function in Dysferlinopathy Animal Models. Ann. Clin. Transl. Neurol. 2015, 2, 256–270. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| hGAPDH_F | TCGGAGTCAACGGATTTGGT |
| hGAPDH_R | TTCCCGTTCTCAGCCTTGAC |
| DYSF_F | CGGTGGGCCATCATTCTGTTT |
| DYSF_R | TCAGGAGAATGGCTTCACCAGTTTCAT |
| Component | Buffer Contents |
|---|---|
| Tris-HCl, pH 6.8 | 200 mM |
| β-mercaptoethanol | 400 mM |
| SDS | 4% |
| bromophenol blue | 0.01% |
| Glycerol | 40% |
| Separating Gel | Stacking Gel | |
|---|---|---|
| Volume | 4 mL | 2 mL |
| % Acrylamide (AA) | 10% | 6% |
| Water | 1.7 mL | 1.4 mL |
| AA:bis-AA (30%) | 1.3 mL | 0.33 mL |
| Buffer | 1.5 Tris (pH 8.8) 1 mL | 1.0 Tris (pH 6.8), 0.25 mL |
| SDS 10% | 40 μL | 20 μL |
| PSA 10% | 40 μL | 20 μL |
| TEMED | 6 μL | 4 μL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yakovlev, I.A.; Emelin, A.M.; Slesarenko, Y.S.; Limaev, I.S.; Vetrova, I.A.; Belikova, L.D.; Grafskaia, E.N.; Bobrovsky, P.A.; Pokrovsky, M.V.; Kuzubova, E.V.; et al. Dual Adeno-Associated Virus 9 with Codon-Optimized DYSF Gene Promotes In Vivo Muscle Regeneration and May Decrease Inflammatory Response in Limb Girdle Muscular Dystrophy Type R2. Int. J. Mol. Sci. 2023, 24, 13551. https://doi.org/10.3390/ijms241713551
Yakovlev IA, Emelin AM, Slesarenko YS, Limaev IS, Vetrova IA, Belikova LD, Grafskaia EN, Bobrovsky PA, Pokrovsky MV, Kuzubova EV, et al. Dual Adeno-Associated Virus 9 with Codon-Optimized DYSF Gene Promotes In Vivo Muscle Regeneration and May Decrease Inflammatory Response in Limb Girdle Muscular Dystrophy Type R2. International Journal of Molecular Sciences. 2023; 24(17):13551. https://doi.org/10.3390/ijms241713551
Chicago/Turabian StyleYakovlev, Ivan A., Aleksei M. Emelin, Yana S. Slesarenko, Igor S. Limaev, Iuliia A. Vetrova, Liliya D. Belikova, Ekaterina N. Grafskaia, Pavel A. Bobrovsky, Mikhail V. Pokrovsky, Elena V. Kuzubova, and et al. 2023. "Dual Adeno-Associated Virus 9 with Codon-Optimized DYSF Gene Promotes In Vivo Muscle Regeneration and May Decrease Inflammatory Response in Limb Girdle Muscular Dystrophy Type R2" International Journal of Molecular Sciences 24, no. 17: 13551. https://doi.org/10.3390/ijms241713551
APA StyleYakovlev, I. A., Emelin, A. M., Slesarenko, Y. S., Limaev, I. S., Vetrova, I. A., Belikova, L. D., Grafskaia, E. N., Bobrovsky, P. A., Pokrovsky, M. V., Kuzubova, E. V., Pokrovsky, V. M., Lebedev, P. A., Bardakov, S. N., Isaev, A. A., & Deev, R. V. (2023). Dual Adeno-Associated Virus 9 with Codon-Optimized DYSF Gene Promotes In Vivo Muscle Regeneration and May Decrease Inflammatory Response in Limb Girdle Muscular Dystrophy Type R2. International Journal of Molecular Sciences, 24(17), 13551. https://doi.org/10.3390/ijms241713551