
HRM Accuracy and Limitations as a Species Typing Tool for Leishmania Parasites

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
2.1. HRM HSP70 Efficiently Identifies the Main Leishmania Species with High Reproducibility
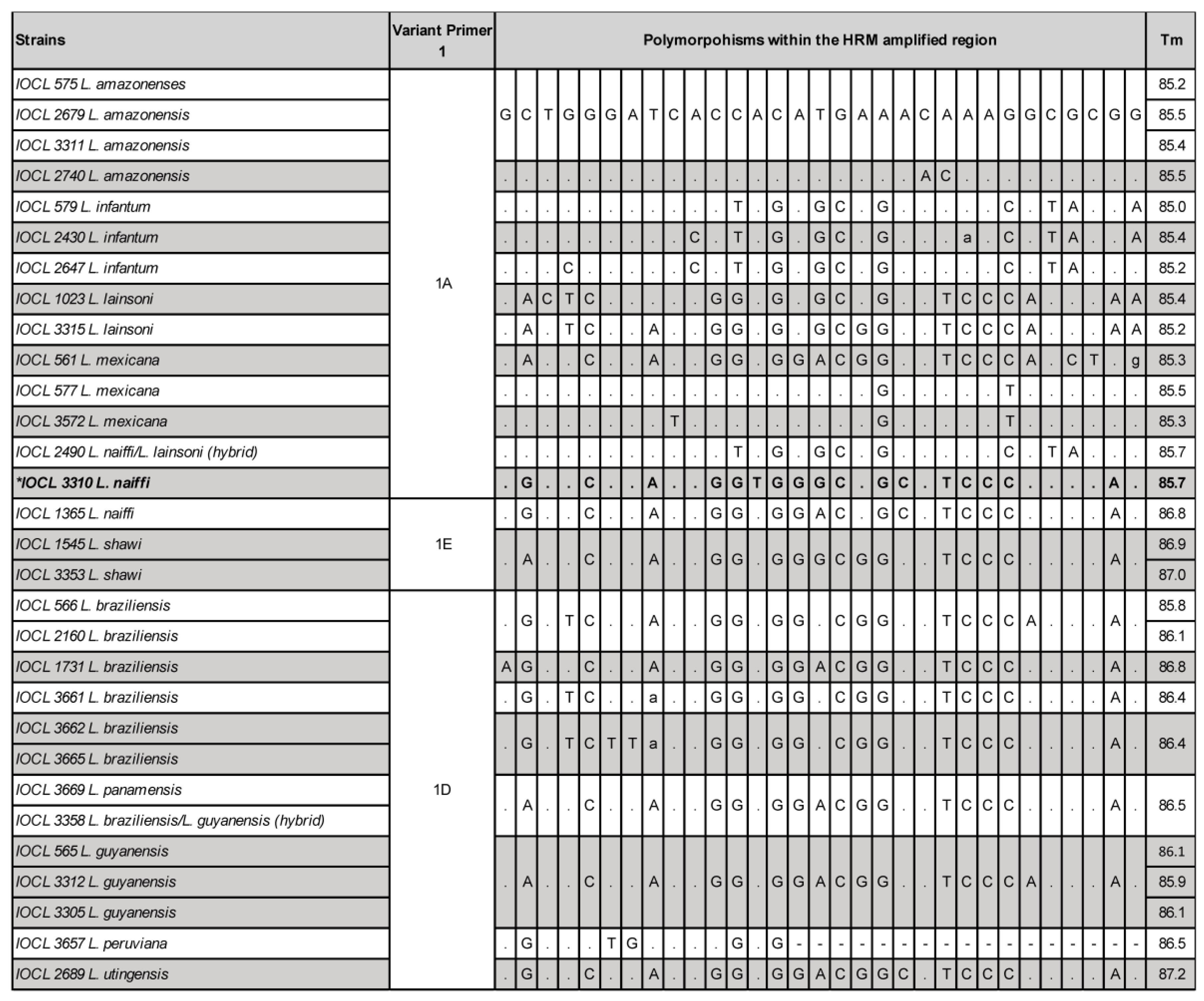
2.2. HRM Discordant Results Cannot Be Directly and Exclusively Associated with Polymorphisms and/or Tms
2.3. HRM HSP70 Presents High Positivity and Similar Sensitivity and Specificity to the Previously Proposed HSP70 qPCR Protocol for Parasite Load Determination
2.4. HRM HSP70 as a Species Typing Tool for Clinical Samples and the Effect of Parasite Load on Its Accuracy
3. Discussion
4. Materials and Methods
4.1. Strains and DNA Samples
4.2. Species Typing
4.3. HRM Reactions and Reproducibility Assay
4.4. Clinical Samples
4.5. Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M.; WHO Leishmaniasis Control Team. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Pan American Health Organization. Manual of Procedures for Leishmaniases Surveillance and Control in the Americas [Internet]. 2019. Available online: https://iris.paho.org/handle/10665.2/51838 (accessed on 3 March 2023).
- Global Report on Neglected Tropical Diseases 2023: Executive Summary [Internet]. Available online: https://www.who.int/publications-detail-redirect/9789240069015 (accessed on 19 September 2023).
- Bailey, M.S.; Lockwood, D.N.J. Cutaneous leishmaniasis. Clin. Dermatol. 2007, 25, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Cantanhêde, L.M.; Mattos, C.B.; de Souza Ronconi, C.; Filgueira, C.P.B.; da Silva Júnior, C.F.; Limeira, C.; de Jesus Silva, H.P.; Ferreira, G.E.M.; Porrozzi, R.; de Godoi Mattos Ferreira, R.; et al. First report of Leishmania (Viannia) lindenbergi causing tegumentary leishmaniasis in the Brazilian western Amazon region. Parasite 2019, 26, 30. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R. The Neotropical Leishmania Species: A Brief Historical Review of Their Discovery, Ecology and Taxonomy. Revista Pan-Amazônica de Saúde [Internet]. 2010. Available online: http://scielo.iec.pa.gov.br/scielo.php?script=sci_arttext&pid=S2176-62232010000200002&lng=en&nrm=iso&tlng=en (accessed on 23 January 2020).
- de Almeida, J.V.; de Souza, C.F.; Fuzari, A.A.; Joya, C.A.; Valdivia, H.O.; Bartholomeu, D.C.; Brazil, R.P. Diagnosis and identification of Leishmania species in patients with cutaneous leishmaniasis in the state of Roraima, Brazil’s Amazon Region. Parasites Vectors 2021, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Cupolillo, E.; Momen, H.; Grimaldi, G. Genetic diversity in natural populations of New World Leishmania. Mem. Inst. Oswaldo. Cruz. 1998, 93, 663–668. [Google Scholar] [CrossRef]
- Brito, M.E.F.; Andrade, M.S.; Mendonça, M.G.; Silva, C.J.; Almeida, E.L.; Lima, B.S.; Félix, S.M.; Abath, F.G.C.; Da Graça, G.C.; Porrozzi, R.; et al. Species diversity of Leishmania (Viannia) parasites circulating in an endemic area for cutaneous leishmaniasis located in the Atlantic rainforest region of northeastern Brazil. Trop. Med. Int. Health 2009, 14, 1278–1286. [Google Scholar] [CrossRef]
- Romero, G.A.S.; Ishikawa, E.; Cupolillo, E.; Toaldo, C.B.; Guerra, M.V.F.; Paes, M.G.; Macêdo, V.O.; Shaw, J.J. The rarity of infection with Leishmania (Viannia) braziliensis among patients from the Manaus region of Amazonas state, Brazil, who have cutaneous leishmaniasis. Ann. Trop. Med. Parasitol. 2002, 96, 131–136. [Google Scholar] [CrossRef]
- da Silva, L.A.; de Sousa, C.D.S.; da Graça, G.C.; Porrozzi, R.; Cupolillo, E. Sequence analysis and PCR-RFLP profiling of the hsp70 gene as a valuable tool for identifying Leishmania species associated with human leishmaniasis in Brazil. Infect. Genet. Evol. 2010, 10, 77–83. [Google Scholar] [CrossRef]
- da Graça, G.C.; Volpini, A.C.; Romero, G.A.S.; de Oliveira Neto, M.P.; Hueb, M.; Porrozzi, R.; Boité, M.C.; Cupolillo, E. Development and validation of PCR-based assays for diagnosis of American cutaneous leishmaniasis and identificatio nof the parasite species. Mem. Inst. Oswaldo. Cruz. 2012, 107, 664–674. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Kubista, M.; Andrade, J.M.; Bengtsson, M.; Forootan, A.; Jonák, J.; Lind, K.; Sindelka, R.; Sjöback, R.; Sjögreen, B.; Strömbom, L.; et al. The real-time polymerase chain reaction. Mol. Asp. Med. 2006, 27, 95–125. [Google Scholar] [CrossRef] [PubMed]
- Talmi-Frank, D.; Nasereddin, A.; Schnur, L.F.; Schönian, G.; Töz, S.Ö.; Jaffe, C.L.; Baneth, G. Detection and Identification of Old World Leishmania by High Resolution Melt Analysis. PLoS Neglected Trop. Dis. 2010, 4, e581. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, M.; Galluzzi, L.; Migliazzo, A.; Magnani, M. Detection and Characterization of Leishmania (Leishmania) and Leishmania (Viannia) by SYBR Green-Based Real-Time PCR and High Resolution Melt Analysis Targeting Kinetoplast Minicircle DNA. PLoS ONE 2014, 9, e88845. [Google Scholar] [CrossRef] [PubMed]
- Hernández, C.; Alvarez, C.; González, C.; Ayala, M.S.; León, C.M.; Ramírez, J.D. Identification of Six New World Leishmania species through the implementation of a High-Resolution Melting (HRM) genotyping assay. Parasites Vectors 2014, 7, 501. [Google Scholar] [CrossRef] [PubMed]
- Reed, G.H.; Kent, J.O.; Wittwer, C.T. High-resolution DNA melting analysis for simple and efficient molecular diagnostics. Pharmacogenomics 2007, 8, 597–608. [Google Scholar] [CrossRef]
- Zampieri, R.A.; Laranjeira-Silva, M.F.; Muxel, S.M.; de Lima, A.C.S.; Shaw, J.J.; Floeter-Winter, L.M. High Resolution Melting Analysis Targeting hsp70 as a Fast and Efficient Method for the Discrimination of Leishmania Species. PLoS Neglected Trop. Dis. 2016, 10, e0004485. [Google Scholar] [CrossRef]
- Filgueira, C.P.B.; Moreira, O.C.; Cantanhêde, L.M.; de Farias, H.M.T.; Porrozzi, R.; Britto, C.; Boité, M.C.; Cupolillo, E. Comparison and clinical validation of qPCR assays targeting Leishmania 18S rDNA and HSP70 genes in patients with American Tegumentary Leishmaniasis. PLoS Neglected Trop. Dis. 2020, 14, e0008750. [Google Scholar] [CrossRef]
- Moreira, O.C.; Yadon, Z.E.; Cupolillo, E. The applicability of real-time PCR in the diagnostic of cutaneous leishmaniasis and parasite quantification for clinical management: Current status and perspectives. Acta Trop. 2018, 184, 29–37. [Google Scholar] [CrossRef]
- Słomka, M.; Sobalska-Kwapis, M.; Wachulec, M.; Bartosz, G.; Strapagiel, D. High Resolution Melting (HRM) for High-Throughput Genotyping—Limitations and Caveats in Practical Case Studies. Int. J. Mol. Sci. 2017, 18, 2316. [Google Scholar] [CrossRef]
- Cortes, S.; Esteves, C.; Maurício, I.; Maia, C.; Cristovão, J.M.; Miles, M.; Campino, L. In vitro and in vivo behaviour of sympatric Leishmania (V.) braziliensis, L. (V.) peruviana and their hybrids. Parasitology 2012, 139, 191–199. [Google Scholar] [CrossRef]
- Kato, H.; Seki, C.; Kubo, M.; Gonzales-Cornejo, L.; Caceres, A.G. Natural infections of Pintomyia verrucarum and Pintomyia maranonensis by Leishmania (Viannia) peruviana in the Eastern Andes of northern Peru. PLoS Neglected Trop. Dis. 2021, 15, e0009352. [Google Scholar] [CrossRef]
- León, C.M.; Muñoz, M.; Hernández, C.; Ayala, M.S.; Flórez, C.; Teherán, A.; Cubides, J.R.; Ramírez, J.D. Analytical Performance of Four Polymerase Chain Reaction (PCR) and Real Time PCR (qPCR) Assays for the Detection of Six Leishmania Species DNA in Colombia. Front. Microbiol. 2017, 8, 1907. [Google Scholar] [CrossRef]
- Rosales-Chilama, M.; Oviedo, M.Y.; Quintero, Y.K.; Fernández, O.L.; Gómez, M.A. Leishmania RNA Virus Is Not Detected in All Species of the Leishmania Viannia Subgenus: The Case of L. (V.) panamensis in Colombia. Am. J. Trop. Med. Hyg. 2023, 108, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Noguera, L.A.; Hernández-Pereira, C.E.; Castillo-Castañeda, A.C.; Patiño, L.H.; Castañeda, S.; Herrera, G.; Mogollón, E.; Muñoz, M.; Duran, A.; Loyo, D.; et al. Diversity and geographical distribution of Leishmania species and the emergence of Leishmania (Leishmania) infantum and L. (Viannia) panamensis in Central-Western Venezuela. Acta Trop. 2023, 242, 106901. [Google Scholar] [CrossRef] [PubMed]
- Torrico, M.C.; Fernández-Arévalo, A.; Ballart, C.; Solano, M.; Rojas, E.; Ariza, E.; Tebar, S.; Lozano, D.; Abras, A.; Gascón, J.; et al. Tegumentary leishmaniasis by Leishmania braziliensis complex in Cochabamba, Bolivia including the presence of L. braziliensis outlier: Tegumentary leishmaniasis in Cochabamba, Bolivia: Tegumentary leishmaniasis in Cochabamba, Bolivia. Transbound. Emerg. Dis. 2022, 69, 2242–2255. [Google Scholar] [CrossRef] [PubMed]
- Kuhls, K.; Cupolillo, E.; Silva, S.O.; Schweynoch, C.; Boité, M.C.; Mello, M.N.; Mauricio, I.; Miles, M.; Wirth, T.; Schönian, G. Population structure and evidence for both clonality and recombination among Brazilian strains of the subgenus Leishmania (Viannia). PLoS Neglected Trop. Dis. 2013, 7, e2490. [Google Scholar] [CrossRef] [PubMed]
- Boité, M.C.; Mauricio, I.L.; Miles, M.A.; Cupolillo, E. New insights on taxonomy, phylogeny and population genetics of Leishmania (Viannia) parasites based on multilocus sequence analysis. PLoS Neglected Trop. Dis. 2012, 6, e1888. [Google Scholar] [CrossRef]
- Garay, A.F.G.; Fraenkel, S.; Diaz, J.J.A.R.; Recalde, O.D.S.; Gómez, M.C.V.; Riquelme, J.A.M.; Arze, P.V.; Centurión, G.N.R.; Britos, M.; Rolón, M. Sensitivity comparison for the Leishmania spp. detection in different canine tissues using PCR-HRM. Rev. Soc. Bras. Med. Trop. 2022, 55. [Google Scholar] [CrossRef]
- Alaeenovin, E.; Parvizi, P.; Ghafari, S.M. Two Leishmania species separation targeting the ITS-rDNA and Cyt b genes by developing and evaluating HRM- qPCR. Rev. Soc. Bras. Med. Trop. 2022, 55. [Google Scholar] [CrossRef]
- Davila, M.; Pineda, V.; Calzada, J.E.; Saldaña, A.; Samudio, F. Evaluation of cytochrome b sequence to identify Leishmania species and variants: The case of Panama. Mem. Inst. Oswaldo. Cruz. 2021, 116, e200572. [Google Scholar] [CrossRef]
- Ewing, B.; Hillier, L.; Wendl, M.C.; Green, P. Base-calling of automated sequencer traces using phred. I. Accuracy assessment. Genome Res. 1998, 8, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Espada, C.R.; Ortiz, P.A.; Shaw, J.J.; Barral, A.M.P.; Costa, J.M.L.; Uliana, S.R.B.; Coelho, A.C. Identification of Leishmania (Viannia) species and clinical isolates of Leishmania (Leishmania) amazonensis from Brazil using PCR-RFLP of the heat-shock protein 70 gene reveals some unexpected observations. Diagn. Microbiol. Infect. Dis. 2018, 91, 312–318. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Voucher | Species | Primer 1 | Primer 2 | Primer 1 | Primer 2 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Exp. 1 | Exp. 2 | Exp. 1 | Exp. 2 | Tm | SD | Tm | SD | ||
| IOCL 565 | L. (V.) guyanensis | 1D | 1D | 2A | 2A | 86.1 | 0.2 | 84.1 | 0.0 |
| IOCL 566 | L. (V.) braziliensis | 1D | 1D | 2B | 2B | 85.8 | 0.1 | 84.7 | 0.1 |
| IOCL 575 | L. (L.) amazonensis | 1A | 1A | - | - | 85.2 | 0.1 | 83.9 | 0.2 |
| IOCL 579 | L. (L.) infantum | 1A | 1A | - | - | 85.0 | 0.1 | 85.1 | 0.0 |
| IOCL 1023 | L. (V.) lainsoni | 1A | 1A | 2A | 2A | 85.4 | 0.1 | 84.2 | 0.1 |
| IOCL 1365 | L. (V.) naiffi | 1E | 1E | 2B | 2B | 85.4 | 0.1 | 84.7 | 0.2 |
| IOCL 1545 | L. (V.) shawi | 1E | 1E | 2A | 2A | 86.7 | 0.1 | 84.0 | 0.1 |
| IOCL 2689 | L. (V.) utingensis | 1E | 1E | 2B | 2B | 87.0 | 0.1 | 84.8 | 0.2 |
| IOCL 2690 | L. (V.) lindenbergi | 1D | 1D | 2A | 2A | 85.9 | 0.1 | 84.1 | 0.1 |
| IOCL 3394 | L. (V.) braziliensis | 1D | 1D | 2B | 2B | 86.1 | 0.1 | 84.6 | 0.0 |
| IOCL 3398 | L. (V.) lainsoni | 1D | 1D | 2A | 2A | 85.7 | 0.2 | 84.1 | 0.0 |
| IOCL 3399 | L. (L.) amazonensis | 1A | 1A | - | - | 85.2 | 0.3 | 84.6 | 0.0 |
| IOCL 3538 | L. (V.) guyanensis | 1D | 1D | 2A | 2A | 86.2 | 0.1 | 84.2 | 0.0 |
| V 1 | V 2 | Voucher (IOCL) | MLEE Characterization | DNA Sequencing | HRM | Tm V1 | Tm V2 |
|---|---|---|---|---|---|---|---|
| 1D | 2A | 3358 | L. braziliensis/L. guyanensis (hybrid) | L. guyanensis | L. guyanensis | 86.5 | 84.1 |
| 1A | 2A | 2490 | L. naiffi/L. lainsoni (hybrid) | L. lainsoni | L. lainsoni | 85.7 | 84.3 |
| 1D | 2B | 390 | L. guyanensis/L. braziliensis (hybrid) | L. braziliensis | L. braziliensis | 86.5 | 84.8 |
| Assays | Patient Samples (n = 60) | ||
| HRM + | HRM − | Total | |
| Microscopy + | 38 (82.6%) | 8 (17.4%) | 46 (100%) |
| Microscopy – | 8 (57.1%) | 6 (42.9%) | 14 (100%) |
| cPCR + | 42 (85.7%) | 7 (14.3%) | 49 (100%) |
| cPCR – | 4 (36.4%) | 7 (63.6%) | 11 (100%) |
| Assays | HSP70 qPCR + | HSP70 qPCR − | Total |
| Microscopy + | 37 (80.4%) | 9 (19.6%) | 46 (100%) |
| Microscopy – | 10 (71.4%) | 4 (28.6%) | 14 (100%) |
| cPCR + | 40 (81.6%) | 9 (18.4%) | 49 (100%) |
| cPCR – | 4 (36.4%) | 7 (63.6%) | 11 (100%) |
| HRM Outcome | Samples Successfully Typed by RFLP (n = 45/60) |
| TOTAL | 45 (100%) |
| Inconclusive | 5/45 (11.2%) |
| Negative | 6/45 (13.3%) |
| Species typing achieved | 34/45 (75.5%) |
| HRM outcome | Samples with HRM species typing data (n = 34/45) |
| Total | 34 (100%) |
| Divergent typing result | 8/34 (23.5%) |
| Convergent typing result | 26/34 (76.5%) |
| ID | Sequence | Fragment | Reference |
|---|---|---|---|
| P1 | HSP70 F2 5′-GGAGAACTACGCGTACTCGATGAAG-3′ HSP70C R 5′- TCCTTCGACGCCTCCTGGTTG-3′ | 144 pb | Zampieri et al. 2016 [19]/Graça et al. 2012 [12] |
| P2 | HSP70 F1 5′-AGCGCATGGTGAACGATGCGTC-3′ HSP70 R1 5′-CTTCATCGAGTACGCGTAGTTCTCC-3′ | 104 pb | Zampieri et al. 2016 [19] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filgueira, C.P.B.; Pitta-Pereira, D.; Cantanhêde, L.M.; Ferreira, G.E.M.; Dos Reis, S.; Cupolillo, E.; Moreira, O.C.; Britto, C.; Boité, M.C. HRM Accuracy and Limitations as a Species Typing Tool for Leishmania Parasites. Int. J. Mol. Sci. 2023, 24, 14784. https://doi.org/10.3390/ijms241914784
Filgueira CPB, Pitta-Pereira D, Cantanhêde LM, Ferreira GEM, Dos Reis S, Cupolillo E, Moreira OC, Britto C, Boité MC. HRM Accuracy and Limitations as a Species Typing Tool for Leishmania Parasites. International Journal of Molecular Sciences. 2023; 24(19):14784. https://doi.org/10.3390/ijms241914784
Chicago/Turabian StyleFilgueira, Camila Patricio Braga, Daniela Pitta-Pereira, Lilian Motta Cantanhêde, Gabriel Eduardo Melim Ferreira, Sayonara Dos Reis, Elisa Cupolillo, Otacilio C. Moreira, Constança Britto, and Mariana Côrtes Boité. 2023. "HRM Accuracy and Limitations as a Species Typing Tool for Leishmania Parasites" International Journal of Molecular Sciences 24, no. 19: 14784. https://doi.org/10.3390/ijms241914784
APA StyleFilgueira, C. P. B., Pitta-Pereira, D., Cantanhêde, L. M., Ferreira, G. E. M., Dos Reis, S., Cupolillo, E., Moreira, O. C., Britto, C., & Boité, M. C. (2023). HRM Accuracy and Limitations as a Species Typing Tool for Leishmania Parasites. International Journal of Molecular Sciences, 24(19), 14784. https://doi.org/10.3390/ijms241914784