
The Effect of the Gut Microbiota on Transplanted Kidney Function

 , , , and
, , , and 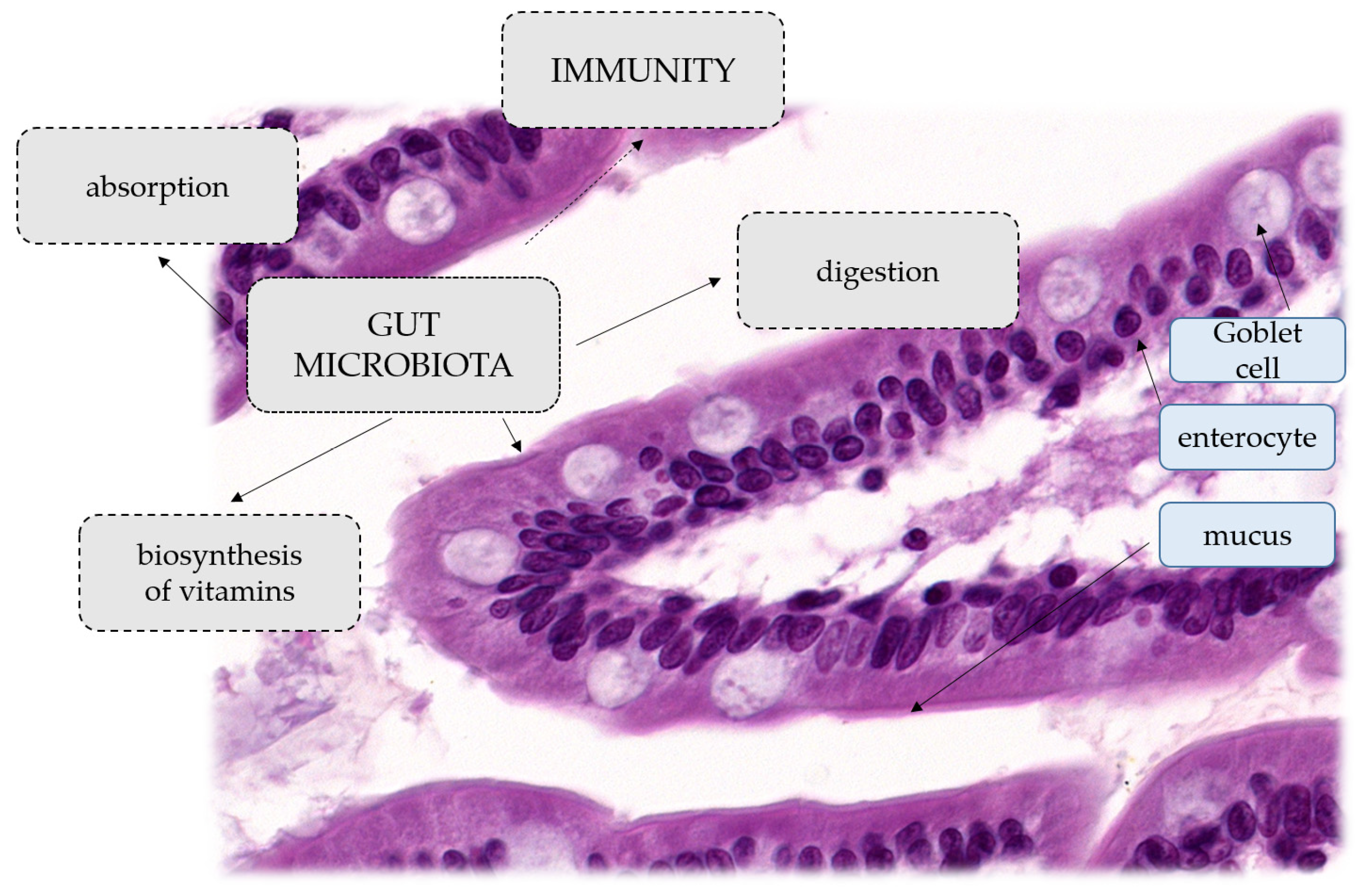{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Gut Microbiota
3. Does the Microbiota Prolong Renal Graft Function?
3.1. Changes in the Microbiota following Kidney Transplantation
3.2. Bidirectional Relationship between the Gut Microbiota and Immunosuppressive Treatment
3.3. Relationship between the Gut Microbiota and Transplantation Complications
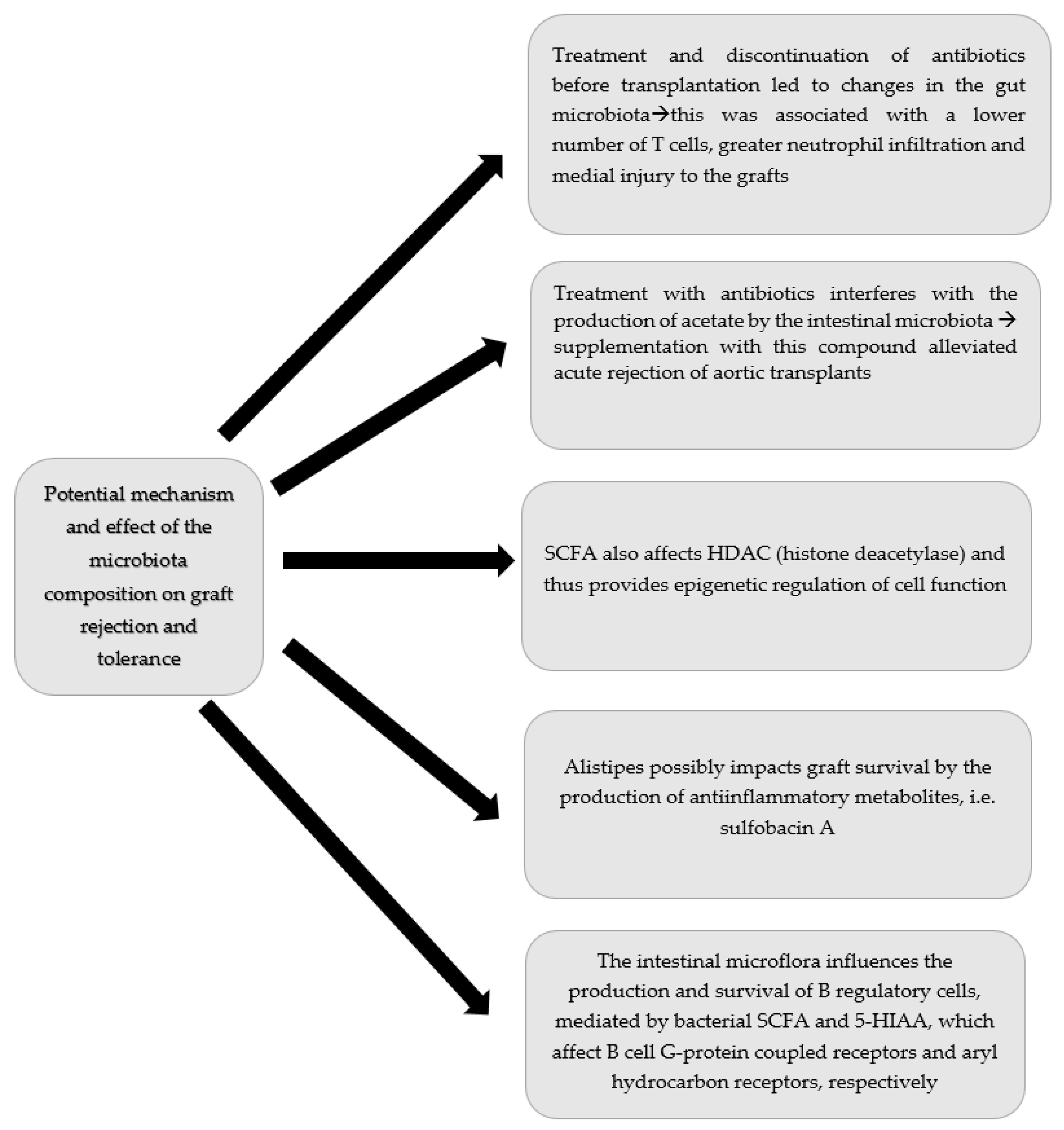
3.4. Effect of the Microbiota Composition on Graft Rejection and Tolerance
3.5. Role of the Microbiota Metabolites after Kidney Transplantation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chern, C.J.; Beutler, E. Biochemical and electrophoretic studies of erythrocyte pyridoxine kinase in white and black Americans. Am. J. Hum. Genet. 1976, 28, 9–17. [Google Scholar] [PubMed]
- Luckey, T.D. Introduction to intestinal microecology. Am. J. Clin. Nutr. 1972, 25, 1292–1294. [Google Scholar] [CrossRef] [PubMed]
- Blaser, M.J. The microbiome revolution. J. Clin. Investig. 2014, 124, 4162–4165. [Google Scholar] [CrossRef] [Green Version]
- Macpherson, A.J.; McCoy, K.D. Stratification and compartmentalisation of immunoglobulin responses to commensal intestinal microbes. Semin. Immunol. 2013, 25, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Chen, D.; Zhang, J.-N.; Lv, X.; Wang, K.; Duan, L.-P.; Nie, Y.; Wu, X.-L. Bacterial Community Mapping of the Mouse Gastrointestinal Tract. PLoS ONE 2013, 8, e74957. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Maurice, C.F.; Haiser, H.J.; Turnbaugh, P.J. Xenobiotics Shape the Physiology and Gene Expression of the Active Human Gut Microbiome. Cell 2013, 152, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Jernberg, C.; Löfmark, S.; Edlund, C.; Jansson, J.K. Long-term ecological impacts of antibiotic administration on the human intestinal microbiota. ISME J. 2007, 1, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Ng, K.M.; Ferreyra, J.A.; Higginbottom, S.K.; Lynch, J.B.; Kashyap, P.C.; Gopinath, S.; Naidu, N.; Choudhury, B.; Weimer, B.C.; Monack, D.M.; et al. Microbiota-liberated host sugars facilitate post-antibiotic expansion of enteric pathogens. Nature 2013, 502, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Pompei, A.; Cordisco, L.; Amaretti, A.; Zanoni, S.; Matteuzzi, D.; Rossi, M. Folate Production by Bifidobacteria as a Potential Probiotic Property. Appl. Environ. Microbiol. 2007, 73, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, J.-H.; Barg, H.; Warren, M.; Jahn, D. Microbial production of vitamin B12. Appl. Microbiol. Biotechnol. 2002, 58, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Petersson, J.; Schreiber, O.; Hansson, G.C.; Gendler, S.J.; Velcich, A.; Lundberg, J.O.; Roos, S.; Holm, L.; Phillipson, M. Importance and regulation of the colonic mucus barrier in a mouse model of colitis. Am. J. Physiol. Liver Physiol. 2011, 300, G327–G333. [Google Scholar] [CrossRef] [Green Version]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hevia, A.; Delgado, S.; Sánchez, B.; Margolles, A. Molecular Players Involved in the Interaction Between Beneficial Bacteria and the Immune System. Front. Microbiol. 2015, 6, 1285. [Google Scholar] [CrossRef] [Green Version]
- Jourová, L.; Anzenbacher, P.; Anzenbacherova, E. Human gut microbiota plays a role in the metabolism of drugs. Biomed. Pap. 2016, 160, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.; Hawley, C.M.; Campbell, K.L.; Morrison, M.; Campbell, S.B.; Isbel, N.M.; Francis, R.S.; Playford, E.G.; Johnson, D.W. Transplant associated infections—The role of the gastrointestinal microbiota and potential therapeutic options. Nephrology 2020, 25, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Gabarre, P.; Loens, C.; Tamzali, Y.; Barrou, B.; Jaisser, F.; Tourret, J. Immunosuppressive therapy after solid organ transplantation and the gut microbiota: Bidirectional interactions with clinical consequences. Am. J. Transplant. 2022, 22, 1014–1030. [Google Scholar] [CrossRef]
- Gibson, C.M.; Childs-Kean, L.M.; Naziruddin, Z.; Howell, C.K. The alteration of the gut microbiome by immunosuppressive agents used in solid organ transplantation. Transpl. Infect. Dis. 2021, 23, e13397. [Google Scholar] [CrossRef]
- Huang, J.; Salinas, T.; Westblade, L.F.; Lee, J.R. The Potential Role of the Gut Microbiota in Kidney Transplantation. Kidney360 2021, 2, 890–893. [Google Scholar] [CrossRef]
- Salvadori, M.; Tsalouchos, A. Microbiota, renal disease and renal transplantation. World J. Transplant. 2021, 11, 16–36. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.R.; Magruder, M.; Zhang, L.; Westblade, L.F.; Satlin, M.J.; Robertson, A.; Edusei, E.; Crawford, C.; Ling, L.; Taur, Y.; et al. Gut microbiota dysbiosis and diarrhea in kidney transplant recipients. Am. J. Transplant. 2019, 19, 488–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarte, J.C.; Douwes, R.M.; Hu, S.; Vila, A.V.; Eisenga, M.F.; van Londen, M.; Gomes-Neto, A.W.; Weersma, R.K.; Harmsen, H.J.; Bakker, S.J. Characteristics and Dysbiosis of the Gut Microbiome in Renal Transplant Recipients. J. Clin. Med. 2020, 9, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fricke, W.F.; Maddox, C.; Song, Y.; Bromberg, J.S. Human Microbiota Characterization in the Course of Renal Transplantation. Am. J. Transplant. 2014, 14, 416–427. [Google Scholar] [CrossRef]
- Rani, A.; Ranjan, R.; McGee, H.S.; Andropolis, K.E.; Panchal, D.V.; Hajjiri, Z.; Brennan, D.C.; Finn, P.W.; Perkins, D.L. Urinary microbiome of kidney transplant patients reveals dysbiosis with potential for antibiotic resistance. Transl. Res. 2017, 181, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Modena, B.D.; Milam, R.; Harrison, F.; Cheeseman, J.A.; Abecassis, M.M.; Friedewald, J.J.; Kirk, A.D.; Salomon, D.R. Changes in Urinary Microbiome Populations Correlate in Kidney Transplants With Interstitial Fibrosis and Tubular Atrophy Documented in Early Surveillance Biopsies. Am. J. Transplant. 2017, 17, 712–723. [Google Scholar] [CrossRef] [Green Version]
- Wilson, I.D.; Nicholson, J.K. Gut microbiome interactions with drug metabolism, efficacy, and toxicity. Transl. Res. 2016, 179, 204–222. [Google Scholar] [CrossRef] [Green Version]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive impact of non-antibiotic drugs on human gut bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef]
- Nakamura, A.; Amada, N.; Haga, I.; Tokodai, K.; Kashiwadate, T. Effects of Elevated Tacrolimus Trough Levels in Association With Infectious Enteritis on Graft Function in Renal Transplant Recipients. Transplant. Proc. 2014, 46, 592–594. [Google Scholar] [CrossRef]
- Tourret, J.; Willing, B.P.; Dion, S.; MacPherson, J.; Denamur, E.; Finlay, B.B. Immunosuppressive Treatment Alters Secretion of Ileal Antimicrobial Peptides and Gut Microbiota, and Favors Subsequent Colonization by Uropathogenic Escherichia coli. Transplantation 2017, 101, 74–82. [Google Scholar] [CrossRef]
- Llorenç, V.; Nakamura, Y.; Metea, C.; Karstens, L.; Molins, B.; Lin, P. Antimetabolite Drugs Exhibit Distinctive Immunomodulatory Mechanisms and Effects on the Intestinal Microbiota in Experimental Autoimmune Uveitis. Investig. Opthalmol. Vis. Sci. 2022, 63, 30. [Google Scholar] [CrossRef] [PubMed]
- Zaza, G.; Gassa, A.D.; Felis, G.; Granata, S.; Torriani, S.; Lupo, A. Impact of maintenance immunosuppressive therapy on the fecal microbiome of renal transplant recipients: Comparison between an everolimus- and a standard tacrolimus-based regimen. PLoS ONE 2017, 12, e0178228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, L.; Tang, H.; Jiao, W.; Zeng, S.; Xu, Y.; Zhang, Q.; Sun, Z.; Mukherjee, A.; Zhang, X.; et al. Immunosuppressive effect of the gut microbiome altered by high-dose tacrolimus in mice. Am. J. Transplant. 2018, 18, 1646–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.R.; Muthukumar, T.; Dadhania, D.; Taur, Y.; Jenq, R.R.; Toussaint, N.C.; Ling, L.; Pamer, E.; Suthanthiran, M. Gut Microbiota and Tacrolimus Dosing in Kidney Transplantation. PLoS ONE 2015, 10, e0122399. [Google Scholar] [CrossRef] [Green Version]
- Jennings, D.L.; Bohn, B.; Zuver, A.; Onat, D.; Gaine, M.; Royzman, E.; Hupf, J.; Brunjes, D.; Latif, F.; Restaino, S.; et al. Gut microbial diversity, inflammation, and oxidative stress are associated with tacrolimus dosing requirements early after heart transplantation. PLoS ONE 2020, 15, e0233646. [Google Scholar] [CrossRef]
- Guo, Y.; Crnkovic, C.M.; Won, K.-J.; Yang, X.; Lee, J.R.; Orjala, J.; Lee, H.; Jeong, H. Commensal Gut Bacteria Convert the Immunosuppressant Tacrolimus to Less Potent Metabolites. Drug Metab. Dispos. 2019, 47, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Qian, L.; Ouyang, H.; Gordils-Valentin, L.; Hong, J.; Jayaraman, A.; Zhu, X. Identification of Gut Bacterial Enzymes for Keto-Reductive Metabolism of Xenobiotics. ACS Chem. Biol. 2022, 17, 1665–1671. [Google Scholar] [CrossRef]
- Zheng, Y.; Masand, A.; Wagner, M.; Kapur, S.; Dadhania, D.; Lubetzky, M.; Lee, J.R. Identification of Antibiotic Administration as a Potentially Novel Factor Associated With Tacrolimus Trough Variability in Kidney Transplant Recipients: A Preliminary Study. Transplant. Direct 2019, 5, e485. [Google Scholar] [CrossRef] [Green Version]
- Simpson, J.B.; Sekela, J.J.; Graboski, A.L.; Borlandelli, V.B.; Bivins, M.M.; Barker, N.K.; Sorgen, A.A.; Mordant, A.L.; Johnson, R.L.; Bhatt, A.P.; et al. Metagenomics combined with activity-based proteomics point to gut bacterial enzymes that reactivate mycophenolate. Gut Microbes 2022, 14, 2107289. [Google Scholar] [CrossRef]
- Khan, M.H.; Onyeaghala, G.C.; Rashidi, A.; Holtan, S.G.; Khoruts, A.; Israni, A.; Jacobson, P.A.; Staley, C. Fecal β-glucuronidase activity differs between hematopoietic cell and kidney transplantation and a possible mechanism for disparate dose requirements. Gut Microbes 2022, 14, 2108279. [Google Scholar] [CrossRef]
- Ishizaki, J.; Tsuda, T.; Suga, Y.; Ito, S.; Arai, K.; Sai, Y.; Miyamoto, K.-I. Change in Pharmacokinetics of Mycophenolic Acid as a Function of Age in Rats and Effect of Coadministered Amoxicillin/Clavulanate. Biol. Pharm. Bull. 2012, 35, 1009–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.R.; Flannigan, K.L.; Rahim, H.; Mohamud, A.; Lewis, I.A.; Hirota, S.A.; Greenway, S.C. Vancomycin relieves mycophenolate mofetil–induced gastrointestinal toxicity by eliminating gut bacterial β-glucuronidase activity. Sci. Adv. 2019, 5, eaax2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saqr, A.; Carlson, B.; Staley, C.; Rashidi, A.; Al-Kofahi, M.; Kaiser, T.; Holtan, S.; MacMillan, M.; Young, J.-A.; El Jurdi, N.; et al. Reduced Enterohepatic Recirculation of Mycophenolate and Lower Blood Concentrations Are Associated with the Stool Bacterial Microbiome after Hematopoietic Cell Transplantation. Transplant. Cell Ther. 2022, 28, 372.e1–372.e9. [Google Scholar] [CrossRef] [PubMed]
- Jardou, M.; Provost, Q.; Brossier, C.; Pinault, É.; Sauvage, F.-L.; Lawson, R. Alteration of the gut microbiome in mycophenolate-induced enteropathy: Impacts on the profile of short-chain fatty acids in a mouse model. BMC Pharmacol. Toxicol. 2021, 22, 66. [Google Scholar] [CrossRef] [PubMed]
- Robles-Vera, I.; de la Visitación, N.; Toral, M.; Sánchez, M.; Gómez-Guzmán, M.; Jiménez, R.; Romero, M.; Duarte, J. Mycophenolate mediated remodeling of gut microbiota and improvement of gut-brain axis in spontaneously hypertensive rats. Biomed. Pharmacother. 2021, 135, 111189. [Google Scholar] [CrossRef]
- Abdel-Nour, M.; Tsalikis, J.; Kleinman, D.; Girardin, S.E. The emerging role of mTOR signalling in antibacterial immunity. Immunol. Cell Biol. 2014, 92, 346–353. [Google Scholar] [CrossRef]
- Huang, E.Y.; Inoue, T.; Leone, V.A.; Dalal, S.; Touw, K.; Wang, Y.; Musch, M.W.; Theriault, B.; Higuchi, K.; Donovan, S.; et al. Using Corticosteroids to Reshape the Gut Microbiome. Inflamm. Bowel Dis. 2015, 21, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Yang, L.; Jiang, J.; Ni, Y.; Zhu, J.; Zheng, X.; Wang, Q.; Lu, X.; Fu, Z. Chronic glucocorticoid treatment induced circadian clock disorder leads to lipid metabolism and gut microbiota alterations in rats. Life Sci. 2018, 192, 173–182. [Google Scholar] [CrossRef]
- He, Z.; Kong, X.; Shao, T.; Zhang, Y.; Wen, C. Alterations of the Gut Microbiota Associated With Promoting Efficacy of Prednisone by Bromofuranone in MRL/lpr Mice. Front. Microbiol. 2019, 10, 978. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A.L. Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 2019, 570, 462–467. [Google Scholar] [CrossRef]
- O’Reilly, C.; O’Sullivan, Ó.; Cotter, P.; O’Connor, P.M.; Shanahan, F.; Cullen, A.; Rea, M.C.; Hill, C.; Coulter, I.; Ross, R.P. Encapsulated cyclosporine does not change the composition of the human microbiota when assessed ex vivo and in vivo. J. Med. Microbiol. 2020, 69, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ma, R.; Riordan, S.M.; Grimm, M.; Liu, L.; Wang, Y.; Zhang, L. Azathioprine, Mercaptopurine, and 5-Aminosalicylic Acid Affect the Growth of IBD-Associated Campylobacter Species and Other Enteric Microbes. Front. Microbiol. 2017, 8, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.R.; Wang, C.Y.; Tang, C.; He, Q.; Li, N.; Li, J.S. Reciprocal Interaction Between Intestinal Microbiota and Mucosal Lymphocyte in Cynomolgus Monkeys After Alemtuzumab Treatment. Am. J. Transplant. 2013, 13, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Bromberg, J.S. Current status of the microbiome in renal transplantation. Curr. Opin. Nephrol. Hypertens. 2016, 25, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Dery, K.J.; Kadono, K.; Hirao, H.; Górski, A.; Kupiec-Weglinski, J.W. Microbiota in organ transplantation: An immunological and therapeutic conundrum? Cell Immunol. 2020, 351, 104080. [Google Scholar] [CrossRef] [PubMed]
- Cooper, L.; Oz, N.; Fishman, G.; Shohat, T.; Rahamimov, R.; Mor, E.; Green, H.; Grossman, A. New onset diabetes after kidney transplantation is associated with increased mortality-A retrospective cohort study. Diabetes/Metab. Res. Rev. 2017, 33, e2920. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.; Zhang, Z.; Xu, Y.; Gong, L.; Zhang, W.; Tang, H.; Zeng, S.; Zhang, Q.; Sun, Z.; Liu, L.; et al. Butyric acid normalizes hyperglycemia caused by the tacrolimus-induced gut microbiota. Am. J. Transplant. 2020, 20, 2413–2424. [Google Scholar] [CrossRef]
- Bhat, M.; Pasini, E.; Copeland, J.; Angeli, M.; Husain, S.; Kumar, D.; Renner, E.; Teterina, A.; Allard, J.; Guttman, D.S.; et al. Impact of Immunosuppression on the Metagenomic Composition of the Intestinal Microbiome: A Systems Biology Approach to Post-Transplant Diabetes. Sci. Rep. 2017, 7, 10277. [Google Scholar] [CrossRef] [Green Version]
- Lecronier, M.; Tashk, P.; Tamzali, Y.; Tenaillon, O.; Denamur, E.; Barrou, B.; Aron-Wisnewsky, J.; Tourret, J. Gut microbiota composition alterations are associated with the onset of diabetes in kidney transplant recipients. PLoS ONE 2020, 15, e0227373. [Google Scholar] [CrossRef] [Green Version]
- Toral, M.; Romero, M.; Rodríguez-Nogales, A.; Jimenez, R.; Robles-Vera, I.; Algieri, F.; Chueca-Porcuna, N.; Sánchez, M.; de la Visitación, N.; Olivares, M.; et al. Lactobacillus fermentum Improves Tacrolimus-Induced Hypertension by Restoring Vascular Redox State and Improving eNOS Coupling. Mol. Nutr. Food Res. 2018, 62, 1800033. [Google Scholar] [CrossRef]
- Pirozzolo, I.; Sepulveda, M.; Chen, L.; Wang, Y.; Lei, Y.M.; Li, Z.; Li, R.; Sattar, H.; Theriault, B.; Belkaid, Y.; et al. Host-versus-commensal immune responses participate in the rejection of colonized solid organ transplants. J. Clin. Investig. 2022, 132, e153403. [Google Scholar] [CrossRef] [PubMed]
- Rey, K.; Manku, S.; Enns, W.; Van Rossum, T.; Bushell, K.; Morin, R.; Brinkman, F.; Choy, J. Disruption of the Gut Microbiota With Antibiotics Exacerbates Acute Vascular Rejection. Transplantation 2018, 102, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, X.; Wu, X.; Wang, Z.; Zhang, C.; Cao, G.; Liu, S.; Yan, T. Gut microbiota alterations associated with antibody-mediated rejection after kidney transplantation. Appl. Microbiol. Biotechnol. 2021, 105, 2473–2484. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, C.M.; Chen, L.; Shaiber, A.; Eren, A.M.; Alegre, M.-L. Gut microbes contribute to variation in solid organ transplant outcomes in mice. Microbiome 2018, 6, 96. [Google Scholar] [CrossRef] [Green Version]
- Pacaud, M.; Colas, L.; Brouard, S. Microbiota and immunoregulation: A focus on regulatory B lymphocytes and transplantation. Am. J. Transplant. 2021, 21, 2341–2347. [Google Scholar] [CrossRef]
- Alhabbab, R.; Blair, P.; Elgueta, R.; Stolarczyk, E.; Marks, E.; Becker, P.D.; Ratnasothy, K.; Smyth, L.; Safinia, N.; Sharif-Paghaleh, E.; et al. Diversity of gut microflora is required for the generation of B cell with regulatory properties in a skin graft model. Sci. Rep. 2015, 5, 11554. [Google Scholar] [CrossRef] [Green Version]
- Lei, Y.M.; Chen, L.; Wang, Y.; Stefka, A.; Molinero, L.L.; Theriault, B.; Aquino-Michaels, K.; Sivan, A.S.; Nagler, C.R.; Gajewski, T.F.; et al. The composition of the microbiota modulates allograft rejection. J. Clin. Investig. 2016, 126, 2736–2744. [Google Scholar] [CrossRef]
- Alegre, M.-L. Can Diet Induce Transplantation Tolerance? J. Am. Soc. Nephrol. 2020, 31, 1417–1418. [Google Scholar] [CrossRef]
- Wu, H.; Singer, J.; Kwan, T.K.; Loh, Y.W.; Wang, C.; Tan, J.; Li, Y.J.; Lai, S.W.C.; Macia, L.; Alexander, S.I.; et al. Gut Microbial Metabolites Induce Donor-Specific Tolerance of Kidney Allografts through Induction of T Regulatory Cells by Short-Chain Fatty Acids. J. Am. Soc. Nephrol. 2020, 31, 1445–1461. [Google Scholar] [CrossRef]
- Colas, L.; Mongodin, E.F.; Montassier, E.; Chesneau, M.; Guerif, P.; Hittle, L.; Giral, M.; Bromberg, J.S.; Brouard, S. Unique and specific Proteobacteria diversity in urinary microbiota of tolerant kidney transplanted recipients. Am. J. Transplant. 2020, 20, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Kim, H.-E.; Cho, H.; Park, J.I.; Kwak, M.-J.; Kim, B.-Y.; Yang, S.H.; Lee, J.P.; Kim, D.K.; Joo, K.W.; et al. Effect of the similarity of gut microbiota composition between donor and recipient on graft function after living donor kidney transplantation. Sci. Rep. 2020, 10, 18881. [Google Scholar] [CrossRef] [PubMed]
- Evenepoel, P.; Poesen, R.; Meijers, B. The gut–kidney axis. Pediatr. Nephrol. 2017, 32, 2005–2014. [Google Scholar] [CrossRef] [PubMed]
- Assem, M.; Lando, M.; Grissi, M.; Kamel, S.; Massy, Z.A.; Chillon, J.-M.; Hénaut, L. The Impact of Uremic Toxins on Cerebrovascular and Cognitive Disorders. Toxins 2018, 10, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosola, C.; Rocchetti, M.T.; Cupisti, A.; Gesualdo, L. Microbiota metabolites: Pivotal players of cardiovascular damage in chronic kidney disease. Pharmacol. Res. 2018, 130, 132–142. [Google Scholar] [CrossRef]
- Poesen, R.; Evenepoel, P.; De Loor, H.; Bammens, B.; Claes, K.; Sprangers, B.; Naesens, M.; Kuypers, D.; Augustijns, P.; Meijers, B. The influence of renal transplantation on retained microbial–human co-metabolites. Nephrol. Dial. Transplant. 2016, 31, 1721–1729. [Google Scholar] [CrossRef] [Green Version]
- Liabeuf, S.; Desjardins, L.; Massy, Z.A.; Brazier, F.; Westeel, P.F.; Mazouz, H.; Titeca-Beauport, D.; Diouf, M.; Glorieux, G.; Vanholder, R.; et al. Levels of Indoxyl Sulfate in Kidney Transplant Patients, and the Relationship With Hard Outcomes. Circ. J. 2016, 80, 722–730. [Google Scholar] [CrossRef] [Green Version]
- Kouidhi, S.; Zidi, O.; Alhujaily, M.; Souai, N.; Mosbah, A.; Belali, T.; Ghedira, K.; El Kossai, I.; El Manaa, J.; Mnif, W.; et al. Fecal Metabolomics Reveals Distinct Profiles of Kidney Transplant Recipients and Healthy Controls. Diagnostics 2021, 11, 807. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Przybyciński, J.; Drożdżal, S.; Wilk, A.; Dziedziejko, V.; Szumilas, K.; Pawlik, A. The Effect of the Gut Microbiota on Transplanted Kidney Function. Int. J. Mol. Sci. 2023, 24, 1260. https://doi.org/10.3390/ijms24021260
Przybyciński J, Drożdżal S, Wilk A, Dziedziejko V, Szumilas K, Pawlik A. The Effect of the Gut Microbiota on Transplanted Kidney Function. International Journal of Molecular Sciences. 2023; 24(2):1260. https://doi.org/10.3390/ijms24021260
Chicago/Turabian StylePrzybyciński, Jarosław, Sylwester Drożdżal, Aleksandra Wilk, Violetta Dziedziejko, Kamila Szumilas, and Andrzej Pawlik. 2023. "The Effect of the Gut Microbiota on Transplanted Kidney Function" International Journal of Molecular Sciences 24, no. 2: 1260. https://doi.org/10.3390/ijms24021260
APA StylePrzybyciński, J., Drożdżal, S., Wilk, A., Dziedziejko, V., Szumilas, K., & Pawlik, A. (2023). The Effect of the Gut Microbiota on Transplanted Kidney Function. International Journal of Molecular Sciences, 24(2), 1260. https://doi.org/10.3390/ijms24021260