
The Effects of Mammary Gland ATIII Overexpression on the General Health of Dairy Goats and Their Anti-Inflammatory Response to LPS Stimulation


 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
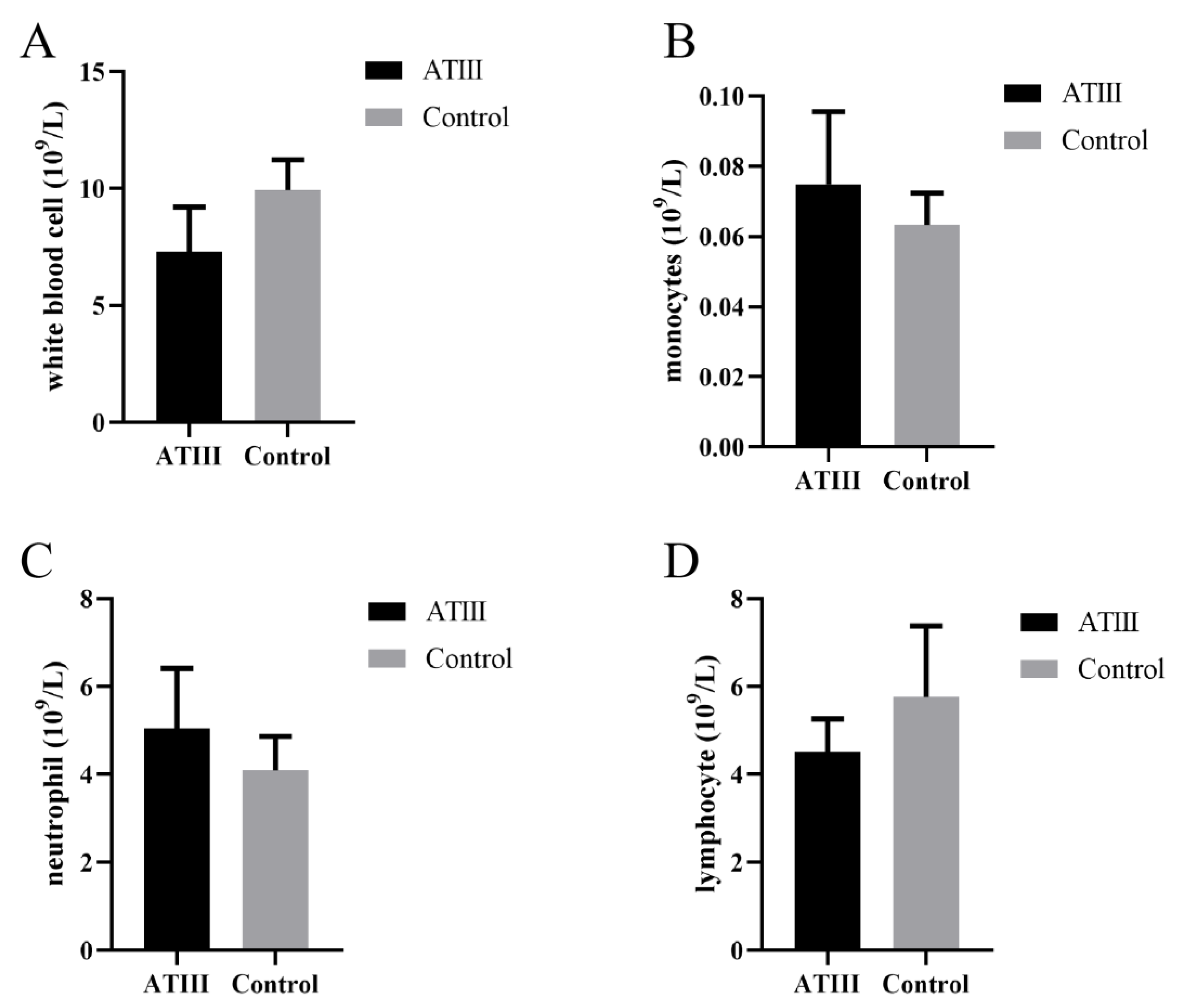
2.1. Effects of Mammary Gland ATIII Overexpression in Dairy Goats on Blood Cells under Normal Conditions
2.2. Effects of Mammary Gland ATIII Overexpression in Dairy Goats on Erythrocytes under Normal Conditions
2.3. Effects of Mammary Gland ATIII Overexpression in Dairy Goats on the Biochemical Parameters of Blood under Normal Conditions
2.4. Effects of Mammary Gland ATIII Overexpression in Dairy Goats on Dairy Herd Improvement (DHI) under Normal Conditions
2.5. Effects of Mammary Gland ATIII Overexpression in Dairy Goats on Immunoglobins and SOD under LPS Stimulation
2.6. Effects of Mammary Gland ATIII Overexpression in Dairy Goats on Pro-Inflammatory Factors after LPS Stimulation
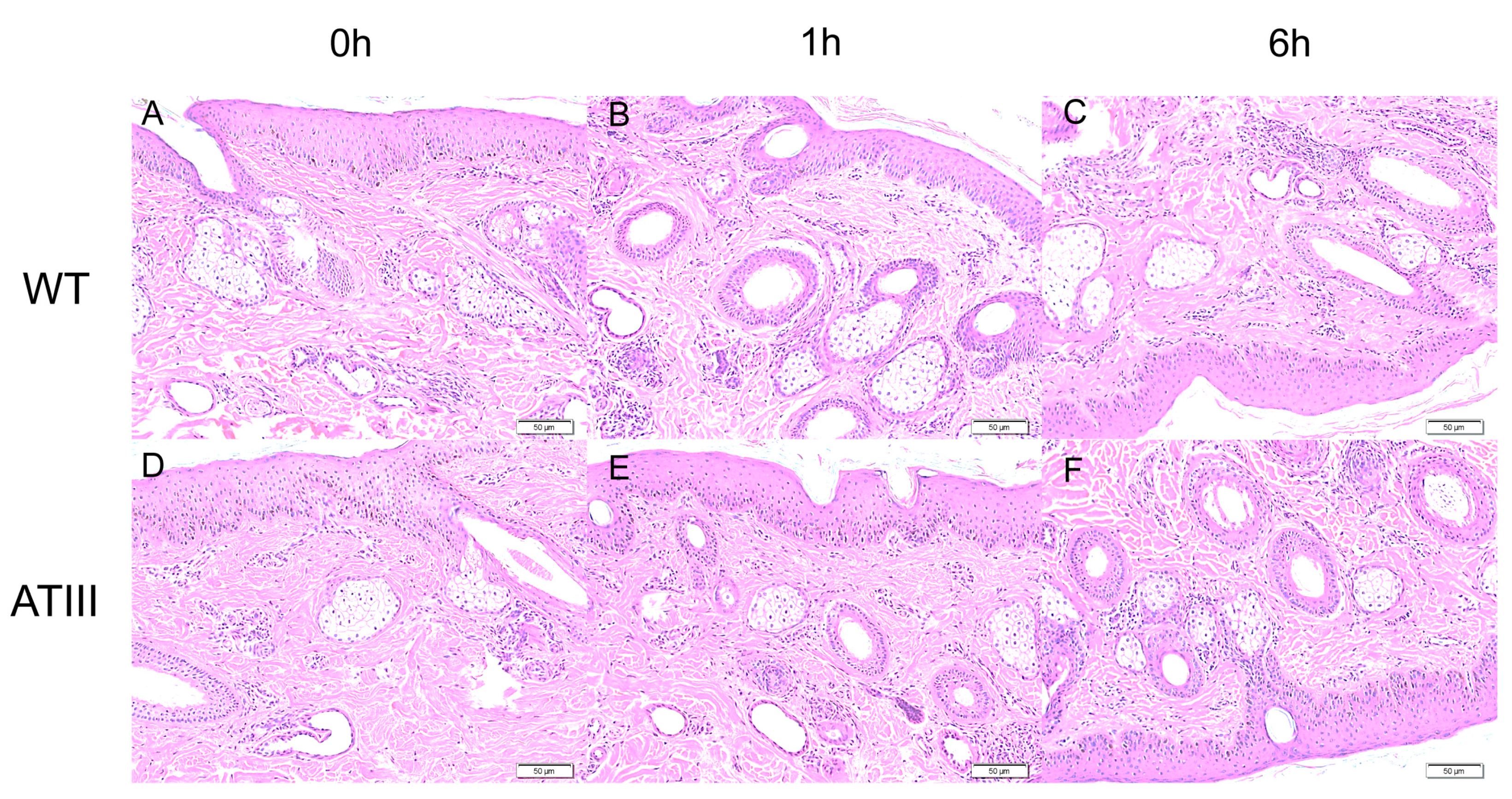
2.7. Effects of Mammary Gland ATIII Overexpression in Dairy Goats on Ear Local Inflammation in after LPS Stimulation
2.8. Effects of LPS Injection on Intestinal Microflora of Dairy Goats with/without Mammary Gland ATIII Overexpression
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Experimental Design
4.3. Analysis of Blood Cells and Blood Biochemical Parameters
4.4. Determination of Milk Composition
4.5. Assays of Proinflammatory Cytokines
4.6. Hematoxylin-Eosin Staining
4.7. Fecal Samples’ 16S rDNA Sequencing
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenberg, R.D. Biochemistry of heparin antithrombin interactions, and the physiologic role of this natural anticoagulant mechanism. Am. J. Med. 1989, 87, 2s–9s. [Google Scholar] [CrossRef]
- Souter, P.J.; Thomas, S.; Hubbard, A.R.; Poole, S.; Römisch, J.; Gray, E. Antithrombin inhibits lipopolysaccharide-induced tissue factor and interleukin-6 production by mononuclear cells, human umbilical vein endothelial cells, and whole blood. Crit. Care Med. 2001, 29, 134–139. [Google Scholar] [CrossRef]
- Menache, D. Antithrombin III: Introduction. Semin. Hematol. 1991, 28, 1–2. [Google Scholar]
- Horie, S.; Ishii, H.; Kazama, M. Heparin-like glycosaminoglycan is a receptor for antithrombin III-dependent but not for thrombin-dependent prostacyclin production in human endothelial cells. Thromb. Res. 1990, 59, 895–904. [Google Scholar] [CrossRef]
- Levy, J.H.; Sniecinski, R.M.; Welsby, I.J.; Levi, M. Antithrombin: Anti-inflammatory properties and clinical applications. Thromb. Haemost. 2016, 115, 712–728. [Google Scholar] [CrossRef]
- Kellner, P.; Nestler, F.; Leimert, A.; Bucher, M.; Czeslick, E.; Sablotzki, A.; Raspè, C. Antithrombin III, but not C1 esterase inhibitor reduces inflammatory response in lipopolysaccharide-stimulated human monocytes in an ex-vivo whole blood setting. Cytokine 2014, 70, 173–178. [Google Scholar] [CrossRef]
- Lu, Z.; Wang, F.; Liang, M. SerpinC1/Antithrombin III in kidney-related diseases. Clin. Sci. 2017, 131, 823–831. [Google Scholar] [CrossRef]
- Hagiwara, S.; Iwasaka, H.; Shingu, C.; Matsumoto, S.; Uchida, T.; Noguchi, T. High-dose antithrombin III prevents heat stroke by attenuating systemic inflammation in rats. Inflamm. Res. 2010, 59, 511–518. [Google Scholar] [CrossRef]
- Opal, S.M.; Garber, G.E.; LaRosa, S.P.; Maki, D.G.; Freebairn, R.C.; Kinasewitz, G.T.; Dhainaut, J.F.; Yan, S.B.; Williams, M.D.; Graham, D.E.; et al. Systemic host responses in severe sepsis analyzed by causative microorganism and treatment effects of drotrecogin alfa (activated). Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2003, 37, 50–58. [Google Scholar] [CrossRef]
- Rauch, U.; Bonderman, D.; Bohrmann, B.; Badimon, J.J.; Himber, J.; Riederer, M.A.; Nemerson, Y. Transfer of tissue factor from leukocytes to platelets is mediated by CD15 and tissue factor. Blood 2000, 96, 170–175. [Google Scholar] [CrossRef]
- Dellinger, R.P.; Levy, M.M.; Rhodes, A.; Annane, D.; Gerlach, H.; Opal, S.M.; Sevransky, J.E.; Sprung, C.L.; Douglas, I.S.; Jaeschke, R.; et al. Surviving Sepsis Campaign: International guidelines for management of severe sepsis and septic shock, 2012. Intensive Care Med. 2013, 39, 165–228. [Google Scholar] [CrossRef]
- Brun-Buisson, C. The epidemiology of the systemic inflammatory response. Intensive Care Med. 2000, 26 (Suppl. S1), S64–S74. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, E.; Abbasi, F.; Mor, G.; Ishigatsubo, Y.; Klinman, D.M. Phenotype and frequency of cells secreting IL-2, IL-4, IL-6, IL-10, IFN and TNF-alpha in human peripheral blood. Cytokine 1995, 7, 815–822. [Google Scholar] [CrossRef]
- Levi, M.; Sivapalaratnam, S. Disseminated intravascular coagulation: An update on pathogenesis and diagnosis. Expert Rev. Hematol. 2018, 11, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Niessen, R.W.; Lamping, R.J.; Jansen, P.M.; Prins, M.H.; Peters, M.; Taylor, F.B., Jr.; de Vijlder, J.J.; ten Cate, J.W.; Hack, C.E.; Sturk, A. Antithrombin acts as a negative acute phase protein as established with studies on HepG2 cells and in baboons. Thromb. Haemost. 1997, 78, 1088–1092. [Google Scholar] [CrossRef]
- Zhao, C.; Hu, X.; Bao, L.; Wu, K.; Feng, L.; Qiu, M.; Hao, H.; Fu, Y.; Zhang, N. Aryl hydrocarbon receptor activation by Lactobacillus reuteri tryptophan metabolism alleviates Escherichia coli-induced mastitis in mice. PLoS Pathog. 2021, 17, e1009774. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Yang, Z.; Weisshaar, J.C. Oxidative stress induced in E. coli by the human antimicrobial peptide LL-37. PLoS Pathog. 2017, 13, e1006481. [Google Scholar] [CrossRef]
- Yu, G.M.; Tan, W. Melatonin Inhibits Lipopolysaccharide-Induced Inflammation and Oxidative Stress in Cultured Mouse Mammary Tissue. Mediat. Inflamm. 2019, 2019, 8597159. [Google Scholar] [CrossRef]
- Murray, J.D.; Maga, E.A. Genetically engineered livestock for agriculture: A generation after the first transgenic animal research conference. Transgenic Res. 2016, 25, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Robl, J.M.; Wang, Z.; Kasinathan, P.; Kuroiwa, Y. Transgenic animal production and animal biotechnology. Theriogenology 2007, 67, 127–133. [Google Scholar] [CrossRef]
- Rogers, C.S. Genetically engineered livestock for biomedical models. Transgenic Res. 2016, 25, 345–359. [Google Scholar] [CrossRef]
- Brondyk, W.H. Selecting an appropriate method for expressing a recombinant protein. Methods Enzymol. 2009, 463, 131–147. [Google Scholar] [CrossRef]
- Keefer, C.L. Production of bioproducts through the use of transgenic animal models. Anim. Reprod. Sci. 2004, 82–83, 5–12. [Google Scholar] [CrossRef]
- Swanson, M.E.; Martin, M.J.; O’Donnell, J.K.; Hoover, K.; Lago, W.; Huntress, V.; Parsons, C.T.; Pinkert, C.A.; Pilder, S.; Logan, J.S. Production of functional human hemoglobin in transgenic swine. Bio/technology 1992, 10, 557–559. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Martin, M.J.; Okabe, J.F.; Truglio, R.A.; Dhanjal, N.K.; Logan, J.S.; Kumar, R. An isologous porcine promoter permits high level expression of human hemoglobin in transgenic swine. Bio/Technology 1994, 12, 55–59. [Google Scholar] [CrossRef]
- Zou, X.; Yuan, S.; Xian, J.; Zhao, Y.; Li, D.; Wang, Y.; Hao, R.; Jie, J.; Liu, S.; Du, M. Large Scale Production of Recombinant Human Antithrombin Il(rhATI) in Transgenic Cloned Goats. Chin. J. Biotechnol. 2008, 109, 117–123. (In Chinese) [Google Scholar] [CrossRef]
- Hand, T.W.; Reboldi, A. Production and Function of Immunoglobulin A. Annu. Rev. Immunol. 2021, 39, 695–718. [Google Scholar] [CrossRef]
- Boes, M.; Prodeus, A.P.; Schmidt, T.; Carroll, M.C.; Chen, J. A critical role of natural immunoglobulin M in immediate defense against systemic bacterial infection. J. Exp. Med. 1998, 188, 2381–2386. [Google Scholar] [CrossRef] [PubMed]
- Roemisch, J.; Gray, E.; Hoffmann, J.N.; Wiedermann, C.J. Antithrombin: A new look at the actions of a serine protease inhibitor. Blood Coagul. Fibrinolysis Int. J. Haemost. Thromb. 2002, 13, 657–670. [Google Scholar] [CrossRef]
- Uchiba, M.; Okajima, K.; Murakami, K.; Okabe, H.; Takatsuki, K. Effects of antithrombin III (AT III) and Trp49-modified AT III on plasma level of 6-keto-PGF1 alpha in rats. Thromb. Res. 1995, 80, 201–208. [Google Scholar] [CrossRef]
- Yamauchi, T.; Umeda, F.; Inoguchi, T.; Nawata, H. Antithrombin III stimulates prostacyclin production by cultured aortic endothelial cells. Biochem. Biophys. Res. Commun. 1989, 163, 1404–1411. [Google Scholar] [CrossRef]
- Mizutani, A.; Okajima, K.; Uchiba, M.; Isobe, H.; Harada, N.; Mizutani, S.; Noguchi, T. Antithrombin reduces ischemia/reperfusion-induced renal injury in rats by inhibiting leukocyte activation through promotion of prostacyclin production. Blood 2003, 101, 3029–3036. [Google Scholar] [CrossRef]
- Tateson, J.E.; Moncada, S.; Vane, J.R. Effects of prostacyclin (PGX) on cyclic AMP concentrations in human platelets. Prostaglandins 1977, 13, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Riva, C.M.; Morganroth, M.L.; Ljungman, A.G.; Schoeneich, S.O.; Marks, R.M.; Todd, R.F., 3rd; Ward, P.A.; Boxer, L.A. Iloprost inhibits neutrophil-induced lung injury and neutrophil adherence to endothelial monolayers. Am. J. Respir. Cell Mol. Biol. 1990, 3, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Okajima, K.; Kushimoto, S.; Isobe, H.; Tanaka, K. Antithrombin reduces ischemia/reperfusion injury of rat liver by increasing the hepatic level of prostacyclin. Blood 1999, 93, 157–164. [Google Scholar] [CrossRef]
- Zhong, K. Curcumin Mediates a Protective Effect Via TLR-4/NF-κB Signaling Pathway in Rat Model of Severe Acute Pancreatitis. Cell Biochem. Biophys. 2015, 73, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.H.; Zegarra-Ruiz, D.F.; Diehl, G.E. Intestinal Microbes in Autoimmune and Inflammatory Disease. Front. Immunol. 2020, 11, 597966. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.S.R.; de Lima, V.C.O.; Piuvezam, G.; de Azevedo, K.P.M.; Maciel, B.L.L.; Morais, A.H.A. Mechanisms of action of anti-inflammatory proteins and peptides with anti-TNF-alpha activity and their effects on the intestinal barrier: A systematic review. PLoS ONE 2022, 17, e0270749. [Google Scholar] [CrossRef]
- Mohammed, Y.; Kootte, R.S.; Kopatz, W.F.; Borchers, C.H.; Büller, H.R.; Versteeg, H.H.; Nieuwdorp, M.; van Mens, T.E. The intestinal microbiome potentially affects thrombin generation in human subjects. J. Thromb. Haemost. JTH 2020, 18, 642–650. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, L.; Wu, H.; Guan, S.; Ma, W.; Fu, Y.; Ji, P.; Lian, Z.; Zhang, L.; Xing, Y.; Wang, B.; et al. The Effects of Mammary Gland ATIII Overexpression on the General Health of Dairy Goats and Their Anti-Inflammatory Response to LPS Stimulation. Int. J. Mol. Sci. 2023, 24, 15303. https://doi.org/10.3390/ijms242015303
Yan L, Wu H, Guan S, Ma W, Fu Y, Ji P, Lian Z, Zhang L, Xing Y, Wang B, et al. The Effects of Mammary Gland ATIII Overexpression on the General Health of Dairy Goats and Their Anti-Inflammatory Response to LPS Stimulation. International Journal of Molecular Sciences. 2023; 24(20):15303. https://doi.org/10.3390/ijms242015303
Chicago/Turabian StyleYan, Laiqing, Hao Wu, Shengyu Guan, Wenkui Ma, Yao Fu, Pengyun Ji, Zhengxing Lian, Lu Zhang, Yiming Xing, Bingyuan Wang, and et al. 2023. "The Effects of Mammary Gland ATIII Overexpression on the General Health of Dairy Goats and Their Anti-Inflammatory Response to LPS Stimulation" International Journal of Molecular Sciences 24, no. 20: 15303. https://doi.org/10.3390/ijms242015303
APA StyleYan, L., Wu, H., Guan, S., Ma, W., Fu, Y., Ji, P., Lian, Z., Zhang, L., Xing, Y., Wang, B., & Liu, G. (2023). The Effects of Mammary Gland ATIII Overexpression on the General Health of Dairy Goats and Their Anti-Inflammatory Response to LPS Stimulation. International Journal of Molecular Sciences, 24(20), 15303. https://doi.org/10.3390/ijms242015303