1. Introduction
Oct4 is a POU domain transcription factor encoded by the
Pou5f1 gene, which is indispensable for maintaining the pluripotent state in embryonic stem cells (ESCs) [
1]. Although Oct4 functions and mechanisms of its action as a transcription factor have been extensively studied [
2,
3], information about the regulation of the
Pou5f1 gene remains limited. On the other hand, the gene ought to be precisely regulated because rather subtle variations in Oct4 protein levels have been shown to exert quite diverse effects on the fate of pluripotent cells [
4,
5]. Additionally, some
Pou5f1 gene elements are important in non-pluripotent cells, for example, in the context of autoimmune diseases [
6]. And while it is widely accepted that
Pou5f1, particularly via the expression of the Oct4 protein isoform A, plays important roles exclusively in pluripotent stem cells and cells of germline lineage [
1,
7], new data challenging this paradigm have been emerging, further emphasizing the necessity of studying
Pou5f1 regulation [
8,
9,
10].
In mice,
Pou5f1 is regulated via a TATA-less promoter and two upstream enhancers, the distal (DE) and proximal (PE) ones, which act in a stage-dependent manner [
11]. The most comprehensive examination of the functioning of these enhancers in naïve vs. primed mouse ESCs was performed by Choi et al., who demonstrated that DE directs
Pou5f1 transcription in naïve ESCs, which corresponds to the early preimplantation epiblast cells. Conversely, in the primed state corresponding to the post-implantation epiblast, control over
Pou5f1 expression shifts to the proximal enhancer (PE) [
12,
13,
14]. Similarly, in humans,
POU5F1 expression is thought to be governed primarily by three enhancers: the orthologs of the mouse DE and PE, as well as by an intronic enhancer responsible, along with DE, for maintaining
POU5F1 expression in the naïve state [
15]. Furthermore, high-throughput genome engineering approaches have been deployed to identify other non-canonical enhancers that can affect
POU5F1 expression. Diao et al. developed the method called CREST-seq, which helped to identify 45
cis-regulatory elements of the
POU5F1 gene located within the 2-Mbp
POU5F1 locus [
16]. Although these regulatory elements showed an impact on
POU5F1 expression in ESCs, the consequences of their perturbations in vivo are still to be evaluated. Also, the collective contribution of these elements remains unclear.
Here, we sought to explore putative regulatory elements located outside the established cis-regulatory elements of Pou5f1 in the context of pluripotency transition from naïve to primed states. Using a reverse genetic approach, we transferred the Pou5f1 gene fragment, previously shown to span essential functional elements, into ectopic chromosome context and found dramatic changes in Oct4 regulation along with severely compromised ESC differentiation capacity.
2. Results
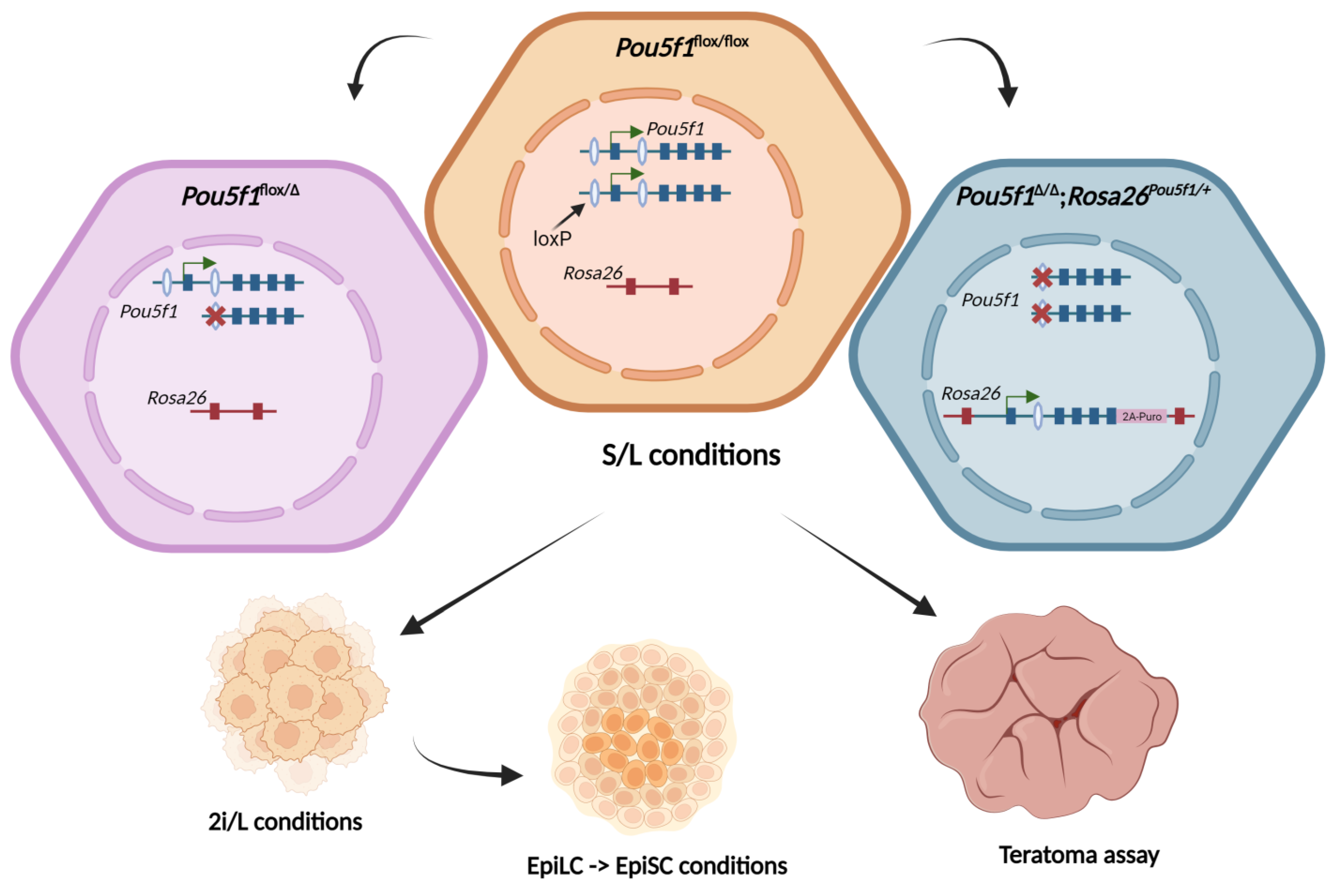
In our previous report, we were able to rescue the self-renewal of mouse null-Oct4 ESCs using the 9.8 kb fragment of the
Pou5f1 gene, which spanned the previously mapped DE and PE, by inserting this fragment into one of the
Rosa26 alleles (
Figure S5). The derived ESCs, referred to as
Pou5f1∆/∆;
Rosa26Pou5f1/+, showed normal proliferation and the expression of the main pluripotency markers (Oct4, Nanog, Klf4) when cultured in the presence of serum and leukemia inhibitory factor (LIF), hereafter designated as S/L conditions [
17]. Accordingly, previous data reported that the monoallelic expression of
Pou5f1 was sufficient to maintain the pluripotent state of
Pou5f1+/– ESCs, which was, surprisingly, even more robust than that of its bi-allelic counterparts [
18]. In this paper, we generated an analogous monoallelic ESC line,
Pou5f1flox/Δ (see
Section 4), and set out to compare it to the parental
Pou5f1flox/flox and
Pou5f1∆/∆;
Rosa26Pou5f1/+ cells for the ability to self-renew in different states of pluripotency, i.e., as naïve ESCs, epiblast-like cells (EpiLCs), and epiblast stem cells (EpiSCs) [
19,
20,
21,
22,
23], as well as for the ability to terminally differentiate in teratoma formation assay (as outlined in
Figure 1).
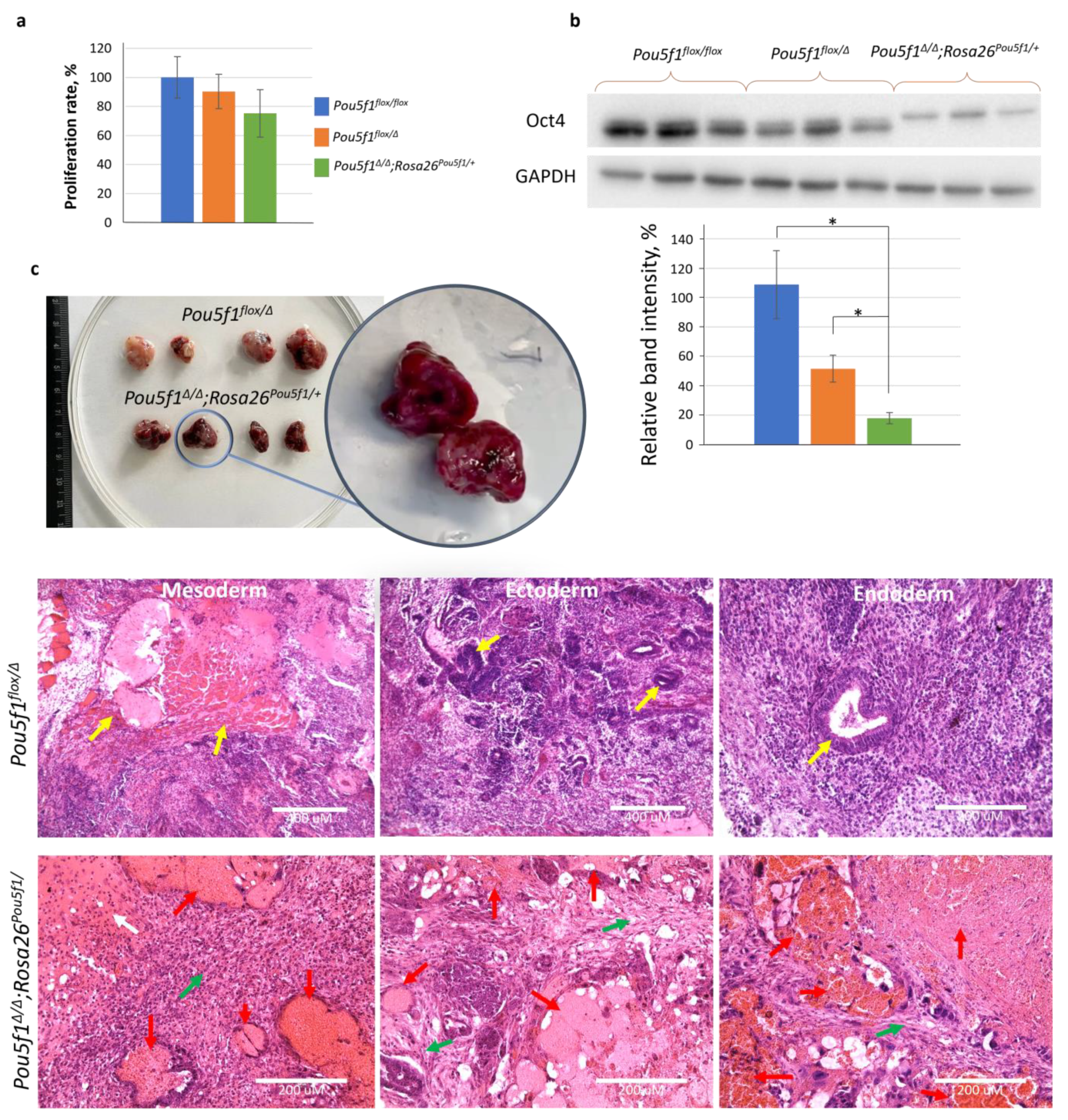
To characterize the ESC lines under the S/L conditions, we compared their proliferation rates and evaluated the Oct4 levels using a Western blot. The three cell lines exhibited similar proliferation characteristics (
Figure 2a), while the Oct4 protein level in
Pou5f1flox/Δ and
Pou5f1∆/∆;Rosa26Pou5f1 ESCs was, respectively, 50% and <25% of that in
Pou5f1flox/flox control cells (
Figure 2b). The molecular weight shift of Oct4 species in
Pou5f1∆/∆;Rosa26Pou5f1/+ cells was apparently due to a portion of the 2A polypeptide remaining covalently linked to the Oct4 C-terminus after self-cleavage of the Oct4-2A-Puro fusion protein. As proof of this statement, the same molecular weight Oct4 species was generated from the
Pou5f1flox−2A-Puro/∆ allele (
Figure S1). Teratomas formed using
Pou5f1flox/Δ ESCs contained derivatives of all three germ layers, whereas
Pou5f1∆/∆;Rosa26Pou5f1/+ ESCs gave rise to tumors with drastically different appearances, as already evident prior to histological analysis (
Figure 2c). This analysis showed that blood-filled cavities comprised most of
Pou5f1∆/∆;Rosa26Pou5f1/+ tumor volume. Solid tissue from these teratomas did not contain germ layer derivatives readily present in
Pou5f1flox/Δ sections but instead cartilage-like and mesenchymal connective tissues, providing a clue about
Pou5f1∆/∆;Rosa26Pou5f1/+ ESC differentiation bias towards the mesoderm lineage (
Figure 2c).
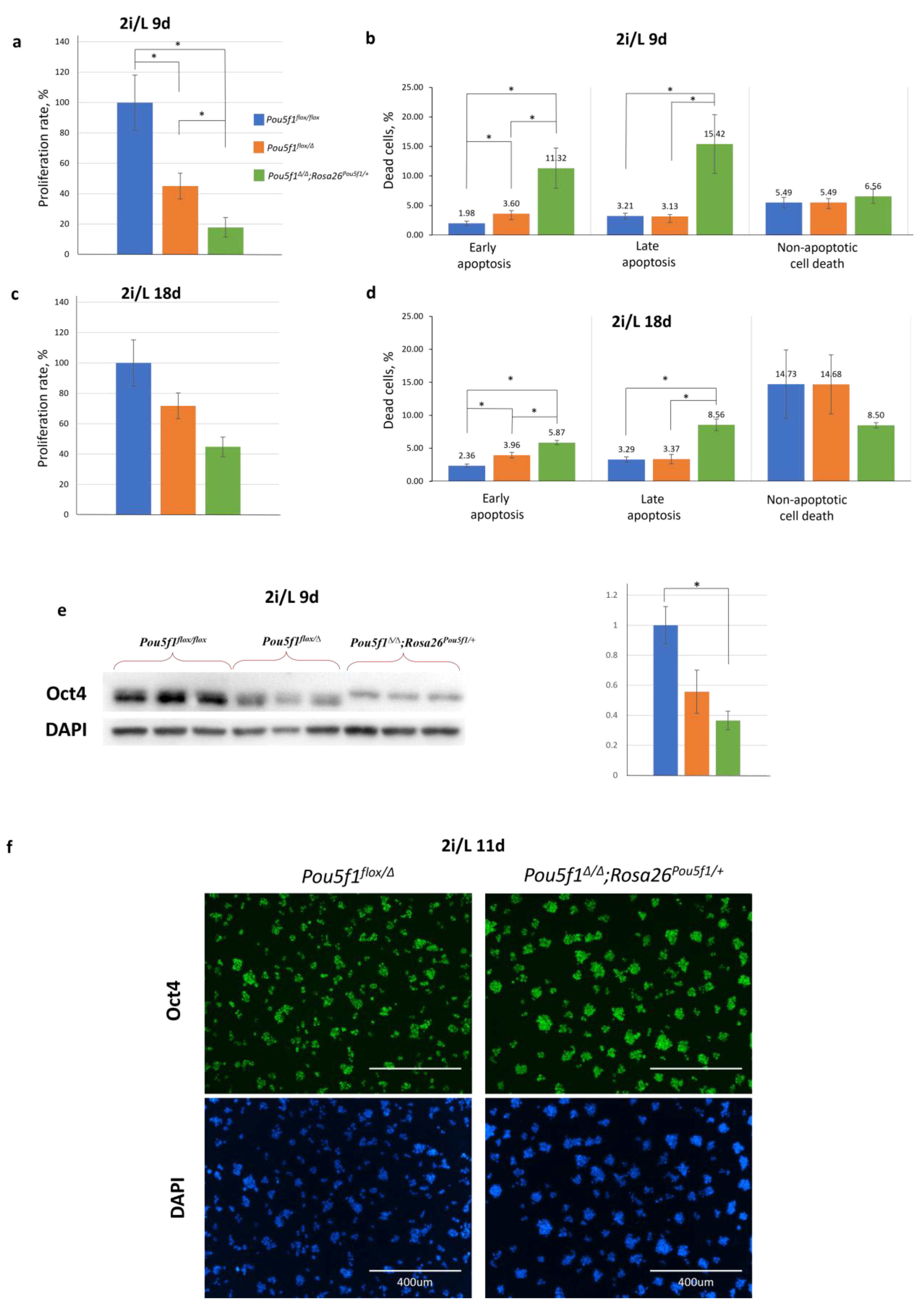
Contrary to the S/L medium, culturing in a serum-free medium supplemented using GSK/MAPK inhibitors (2i) and LIF, referred hereafter to as 2i/L medium, for 9 days resulted in a proliferation decrease in the
Pou5f1flox/Δ and especially, in
Pou5f1∆/∆;Rosa26Pou5f1/+ ESCs, as compared to the control
Pou5f1flox/flox ESCs (
Figure 3a). In addition, the
Pou5f1∆/∆;Rosa26Pou5f1/+ ESCs demonstrated a 3- to 5-fold increase in apoptosis compared to the other two lines (
Figure 3b). At the same time, the non-apoptotic cell death rate was approximately the same in all three cell lines. Interestingly, after an additional 9 days (18 days in total) of culturing under 2i/L conditions, the proliferation rate of
Pou5f1∆/∆;Rosa26Pou5f1/+ ESCs increased twofold (
Figure 3c), while apoptotic processes reduced almost twofold (
Figure 3d), indicating the progressive adaptation of these cells to the 2i/L conditions. Contrary to the S/L conditions (
Figure 2b), 2i/L conditions featured Oct4 levels in
Pou5f1∆/∆;Rosa26Pou5f1/+ ESCs, nearly reaching that in
Pou5f1flox/Δ cells, i.e., 50% of the bi-allelic expression level (
Figure 3e,f).
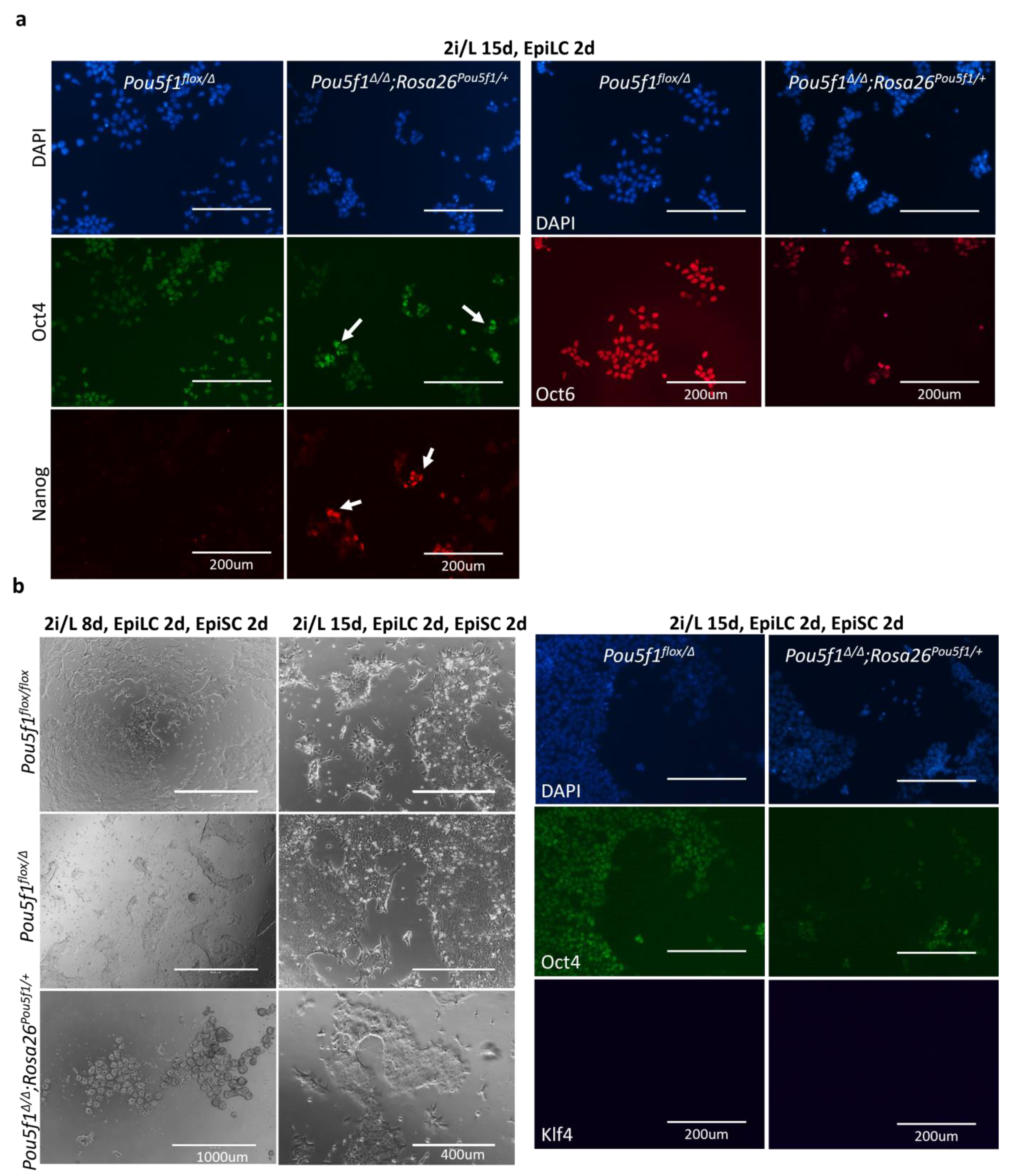
As terminal differentiation of ESCs proceeds through formative and primed pluripotency, we took a closer look at these intermediate states, featured by the cultured epiblast-like cells (EpiLCs) and epiblast stem cells (EpiSCs), respectively [
21,
22,
23]. In the EpiLCs, the differences between the
Pou5f1∆/∆;Rosa26Pou5f1/+ and
Pou5f1flox/Δ ESCs, observed under the 2i/L conditions, further increased. Expression of the pluripotency state markers Oct4, Nanog, and Oct6 was notably perturbed in
Pou5f1∆/∆;Rosa26Pou5f1/+ cells (
Figure 4a). Nanog expression, which is normally suppressed in the EpiLC state, was retained in a significant fraction of
Pou5f1∆/∆;Rosa26Pou5f1/+cells, while the Oct4 expression showed a heterogenous expression pattern, with some cells retaining high levels and the others showing decreased levels of the Oct4 protein. Oct6, in turn, was expressed only in a minor subset of
Pou5f1∆/∆;Rosa26Pou5f1/+ cells, compared to the control
Pou5f1flox/Δ cells (
Figure 4a).
When we proceeded further to EpiSC differentiation,
Pou5f1∆/∆;Rosa26Pou5f1/+ cells progressively lost their ability to attach to the fibronectin-coated surface, completely changing their morphology. Interestingly, when culturing under the 2i/L conditions was extended to 15 days prior to the primed state induction, differentiated
Pou5f1∆/∆;Rosa26Pou5f1/+ cells were notably better at attaching to the fibronectin-coated surface, although they still showed deeply perturbed expressions of the EpiSC markers Oct4 and Oct6 (
Figure 4b and
Figure S3). At the same time, these cells were negative for Klf4 (
Figure 4b), indicating their exit from a naïve state. In sum, the obtained data suggest that pluripotency continuum is severely perturbed in
Pou5f1∆/∆;Rosa26Pou5f1/+ cells. These cells can exit the naïve state; however, they fail (1) to properly maintain the Oct4 levels in formative and primed states, (2) to down-regulate Nanog in the formative state, and (3) to fully establish formative and primed states of pluripotency.
To rule out the possibility of an unspecific impact of the
Rosa26 locus
cis-regulatory elements on
Pou5f1 transgene expression within the
Rosa26Pou5f1 allele, we inserted a cassette expressing tdTomato under the CAG promoter control into the same locus position; we then compared the tdTomato intensity under S/L and 2i/L conditions. We did not observe any increase in the tdTomato signal upon switching from the S/L to 2i/L conditions (
Figure S4), supporting the conclusion that the observed differentiation defects of
Pou5f1∆/∆;Rosa26Pou5f1/+ ESCs occur because of some essential
cis-regulatory elements missing in the 9.8 kb
Pou5f1 transgene, rather than because of those elements within
Rosa26 modulating the transcriptional activity of this transgene.
3. Discussion
In the present work, we evaluated the consequences of the translocation of the
Pou5f1 gene fragment spanning
cis-regulatory elements, previously defined as directing proper spatiotemporal
Pou5f1 expression during mouse ontogeny—the DE and PE [
11]—in an ectopic chromosomal context, as featured by the
Pou5f1∆/∆;
Rosa26Pou5f1/+ ESCs. We found that these ESCs resemble those with monoallelic
Pou5f1 expression within a native chromosomal context (
Pou5f1flox/∆) in terms of their self-renewal under S/L conditions; however, these ESCs showed severely compromised differentiation capacity in teratoma assay with a significant bias towards mesoderm. Accordingly,
Pou5f1∆/∆;Rosa26Pou5f1/+ cells showed decreased proliferation and increased apoptosis rates. When we proceeded to differentiation, these cells failed to properly maintain the Oct4 levels in the formative and primed states, as well as to down-regulate Nanog in the formative state. Overall,
Pou5f1∆/∆;
Rosa26Pou5f1/+ cells failed to properly enter these states of pluripotency on their way to terminal differentiation.
It is likely that observed defects of
Pou5f1∆/∆;Rosa26Pou5f1/+ cells are caused due to an improper regulation of
Pou5f1 translocated to the
Rosa26 locus. Although DE was previously defined DE as crucial for
Pou5f1 expression in the naïve state, there may be additional regulatory elements that maintain
Pou5f1 expression at an appropriate level. Of note, recently described
Pou5f1 elements may serve this purpose [
16]. Mouse orthologs should be examined for their ability to regulate
Pou5f1 expression as well. These elements were revealed in human ESCs, which were in the primed pluripotency state, although
Pou5f1∆/∆;Rosa26Pou5f1/+ cells exhibited the most severe abnormalities during the transition from naïve to primed pluripotency states. Furthermore, under the naïve conditions, Oct4 expression in
Pou5f1∆/∆;Rosa26Pou5f1/+ ESCs nearly caught up with that in
Pou5f1flox/∆ cells, while the elements revealed by Diao et al. acted as enhancers, and their lack led to a reduced Oct4 expression. More relevant to our case, the deletion of an element just downstream of the
Pou5f1 RNA-coding sequence, which is not considered critical for
Pou5f1 regulation (contrary to the DE and PE), led to approximately 50% up-regulation of randomly integrated
Pou5f1(GOF)-lacZ transgene in the inner cell mass of preimplantation embryos, which contain epiblast cells in the naïve state of the pluripotency [
11]. The authors did not assess the impact of this element in the context of a naïve-to-primed state transition, but the
Rosa26Pou5f1 allele in our cells also lacked this downstream element, which might be the main reason for the observed phenotype. An alternative possibility is that the observed mis-regulation was caused by genetic elements located in the
Rosa26 locus. In this case, these putative elements should act selectively upon 9.8 kb
Pou5f1 transgene as they do not affect the expression of CAG- transgene upon S/L to 2i/L condition switch. Further experiments using
Pou5f1 translocation into other safe-harbor or random genomic loci should address the issue of
Pou5f1 transgene–interaction with the genetic surrounding.
Another observation is rather intriguing. Though
Pou5f1∆/∆;
Rosa26Pou5f1/+ cells after 9–11 days of culturing under 2i/L conditions were close to the control cells in terms of naïve state markers expression (Oct4, Nanog, and Klf4) and showed no sign of premature transition to the formative/primed state (Oct6), prolonged exposure to the 2i/L conditions partially restored cell proliferation and survival of
Pou5f1∆/∆;
Rosa26Pou5f1/+ ESCs (
Figure 3 and
Figure S2). This means that these ESCs can adapt to
Pou5f1 mis-regulation via mechanisms such as epigenetic erasing, which is a known feature of the naïve pluripotency state [
25,
26,
27].
Overall, our data suggest that correct spatiotemporal regulation of the
Pou5f1 gene requires
cis-regulatory elements in addition to the previously characterized DE and PE [
11,
12]. Future attempts should focus on identifying these elements, and we believe that our approach, combined with modern techniques of chromatin state assessment, will be of great help in this effort.
4. Materials and Methods
4.1. Animals
All animal procedures were performed according to the guidelines for the humane use of laboratory animals, using standards corresponding to those prescribed by the American Physiological Society. The derivation of mouse embryonic fibroblast (MEFs) and teratoma formation was performed in the Institute of Cytology strictly in agreement with the animal protection legislation acts of the Russian Federation and was approved as humane use of laboratory animals by the Institute’s Ethical Board (protocol No. 17/22, signed 9 November 2022).
4.2. Cell Culture
All cells were cultured in humidified CO
2 incubators at 20% O
2 and 5% CO
2 at 37 °C. MEFs derived from CBA embryos as per the established procedure [
28] were routinely cultured on gelatin-coated cell culture dishes in DMEM GlutaMAX (Gibco, ThermoFischer Scientific, Waltham, MA USA) supplemented with 10% FBS (Biosera, Cholet, France) and 1 × penicillin/streptomycin (Gibco) passaged using 0.05% Trypsin-0.01% EDTA (Gibco). On passage 3–5, MEFs were treated for 2.5 h with 10 μg/mL mitomycin C (MMC; Sigma-Aldrich M0503, Darmstadt, Germany). Mouse ESCs were cultured on the MMC-inactivated MEFs (pre-seeded at a density of approximately 30,000 cells/cm
2 on gelatin-coated cell culture dishes) in KnockOut DMEM (Gibco) supplemented with 15% FBS (Biosera), 1× NEAA (Gibco), 1× penicillin/streptomycin (Gibco), 0.1 mM 2-mercaptoethanol (Sigma-Aldrich), 2 mM l-glutamine (Gibco), and 1:5000 of in-house-made hLIF, referred to as S/L medium. ESCs were put into naïve pluripotency state by culturing on poly-L-ornithine (0.01%; Sigma)-coated plates in the N2B27 media supplemented with, 3 μM CHIR99021 (Axon, Groningen, The Netherlands), 1 μM PD0325901 (Axon), and 1:5000 of in-house made hLIF, designated as the 2i/L medium. The N2B27 media included DMEM/F12 (Gibco) and Neurobasal medium (1:1), supplemented with 1× N2, 1× B27 (without RA) (Gibco), 50 μM β-mercaptoethanol (Sigma Aldrich), 0.005% BSA (Sigma), 1× penicillin/streptomycin (Gibco), and 2 mM L-glutamine (Gibco). To promote differentiation into EpiLCs, ESCs cultured in the 2i/L medium were plated for 2 days on fibronectin-coated plates in N2B27 media supplemented with 1% knockout serum replacement (KSR; ThermoFisher Scientific, Waltham, MA USA), 20 ng/mL activin, and 12 ng/mL bFGF. To further proceed to the differentiation into EpiSCs, media was additionally supplemented with 6.25 mg/mL XAV 939.
4.3. Transient Transfection
ESCs were seeded at a density of 5 × 103/cm2 per well of 24-well plates. The next day, media were changed to fresh S/L media. After two hours, the transfection mix (1:2 ratio, mixed in 250 µL of OptiMEM) was added into the wells. Twelve hours post-transfection, the media were changed to the fresh S/L medium; after an additional 24 h, medium was replaced with the same medium containing antibiotics.
4.4. Establishment of Mutant ESC Lines
Derivation of
Pou5f1∆/∆;Rosa26Pou5f1/+ ESCs was previously reported [
17]. To generate
Pou5f1∆/flox ESCs, ESCs derived from
Pou5f1flox/flox, blastocysts were seeded at a density of 5 × 10
3 per cm
2 and, with the purpose to achieve partial recombination, transiently transfected with the Cre-expressing plasmid
pRosa26-TRE-CAG-Frt(Ert2CreErt2-STOP)Frt-tdTomatoPGKneo [
10,
24]. Twenty-four hours after transfection, the media were changed to the S/L media containing 4-hydroxytamoxifen (4-OHT). The next day, cells were seeded in a 6-well plate and cultured in the presence of 500 µg/mL Geneticin (G-418) (#108321-42-2, bioWORLD, Dublin, Ohio, USA) for 4 days.
4.5. Immunocytochemistry
Cells were fixed in 4% PFA (Sigma) for 10 min, permeabilized with 0.1% Triton X-100 for 15 min, blocked in 3% BSA for 1 h at RT, and incubated with primary antibodies overnight at 4 °C. The next day, cells were washed 5–6 times with PBS, incubated with appropriate secondary fluorescent antibodies for 2 h at RT, and then washed and stained with DAPI. Immunochemistry images were taken on the fluorescent microscope EVOS FL AUTO (Life Technologies, ThermoFischer Scientific, Waltham, MA USA). The following primary antibodies were used: mouse anti-Oct4 (1:500, C-10, sc-5279, Santa Cruz Biotechnology, Dallas, TX, USA), rabbit anti-Oct4 (1:300, ab19857, Abcam, Cambridge, UK), rat anti-Nanog (1:250, 14-5761-80, eBioscience, ThermoFischer Scientific, Waltham, MA USA), rabbit anti-Klf4 (1:250, ab129473, Abcam), and rabbit anti-Oct6 (1:250, ab272925, Abcam). The following secondary antibodies were used at 1:500 dilutions: goat-anti-rat Alexa Fluor 555 (Invitrogen, ThermoFischer Scientific, Waltham, MA USA), goat-anti-mouse Alexa Fluor 488, and goat-anti-rabbit Alexa Fluor 647 (Jackson ImmunoResearch, West Grove, PA, USA).
4.6. Annexin V-FITC/PI Staining
Annexin V and PI staining were performed using the Annexin V-FITC Apoptosis Detection Kit (ab14085, Abcam) according to the manufacturer’s instructions. In brief, cells were washed with PBS and harvested at 300 g for 3 min; the supernatant was discarded, and the pellet was resuspended in 1× binding buffer at a density of 1–5 × 105 cells per mL. Samples were then incubated with 5 μL of FITC-conjugated annexin V for 15 min at RT in a dark place. Propidium iodide (PI) was added to the samples right before analysis. Flow cytometry was performed using the CytoFLEX flow cytometer (Beckman Coulter, Brea, CA, USA), and data were analyzed using CytoFLEX CytExpert software (ver 2.3).
4.7. Teratoma Formation Assay
Mouse ESCs, cultured on gelatin-coated dishes in ESC medium, were harvested with 0.05% Trypsin-0.01% EDTA (Gibco), resuspended in PBS, and injected subcutaneously (2 × 106 cells) into athymic CD-1 NUDE mice maintained under specific pathogen-free conditions. Four animals were used, with two injections per mouse. Five weeks after the injections, the mice were sacrificed by cervical dislocation and the teratomas were surgically removed. We then proceeded to sectioning and histological analysis.
4.8. Preparation of Sections for Histological Analysis
Excised teratomas were washed in PBS and fixed for 24 h at RT in two types of fixators: Bouin’s fluid and 10% formalin. After performing Bouin’s fixation, the teratomas were washed in several changes of 70% alcohol until the solution was lightened. After the formalin fixation, teratomas were washed in running water for 10 min and transferred to 70% alcohol. Specimens were dehydrated in an ethanol series and isobutanol:paraffin series and then embedded in paraffin (McCormick Scientific, now Leica Biosystems, Nussloch, Germany). For each teratoma, blocks of approximately 5 × 5 mm were used for analysis. Paraffin sections were washed in xylene, rehydrated using an ethanol series (100–70%), and then washed with water. Next, sections were incubated in hematoxylin for 6 min, washed with water, and incubated with eosin for 3 min. After washing and dehydration, sections were mounted in Vitrogel (BioVitrum, St-Petersburg, Russia) for further analysis.
4.9. Western Blotting
We lysed 1 × 106 cells in the RIPA buffer (Invitrogen) containing cOmplete™ ULTRA protease-inhibitor (Roche, Basel, Switzerland) and sonicated the products. SDS-PAGE electrophoresis was performed using 10% polyacrylamide gel in a Tris-HCl buffer. Protein transfer was performed using the CD10 semi-dry system (Cleaver Scientific, Rugby, UK). After blocking with 3% BSA, the membranes were incubated with the following primary antibodies: mouse anti-Oct4 (1:1000, C-10, sc-5279, Santa Cruz Biotechnology), rabbit anti-Oct4 (1:1000, ab19857, Bethyl, Fortis Life Sciences, Waltham, MA USA), rabbit anti-GAPDH (1:2000, 2118s, Cell signaling, Danvers, MA, USA). After washing with PBS, membranes were incubated with HRP-conjugated secondary antibodies (1:10,000, Jackson ImmunoResearch, Ely, UK). Visualization was performed using a Chemidoc Touch device (BIO-RAD, Hercules, CA, US).
4.10. Quantification and Statistical Analysis
To determine cellular proliferation, all cell lines were seeded with the same cell concentration and monitored by flow cytometry every 3–4 days. The number of cells in control (Pou5f1flox/flox) ESCs was taken as 100%. Four biological replicates were used for each cell line to calculate growth rates and apoptosis levels under the S/L and 2iL conditions. Three biological replicates were employed to evaluate the relative intensity of the band. IMAGEJ Fiji was utilized to carry out the analysis of Western blot images. The data is presented as mean ± SEM (Standard error of the mean). The statistical significance of small samples was evaluated using the non-parametric Mann-Whitney U-test. Differences were considered statistically significant at p < 0.05.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}



