
Counter-Intuitive Features of Particle Dynamics in Nanopores


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (i).
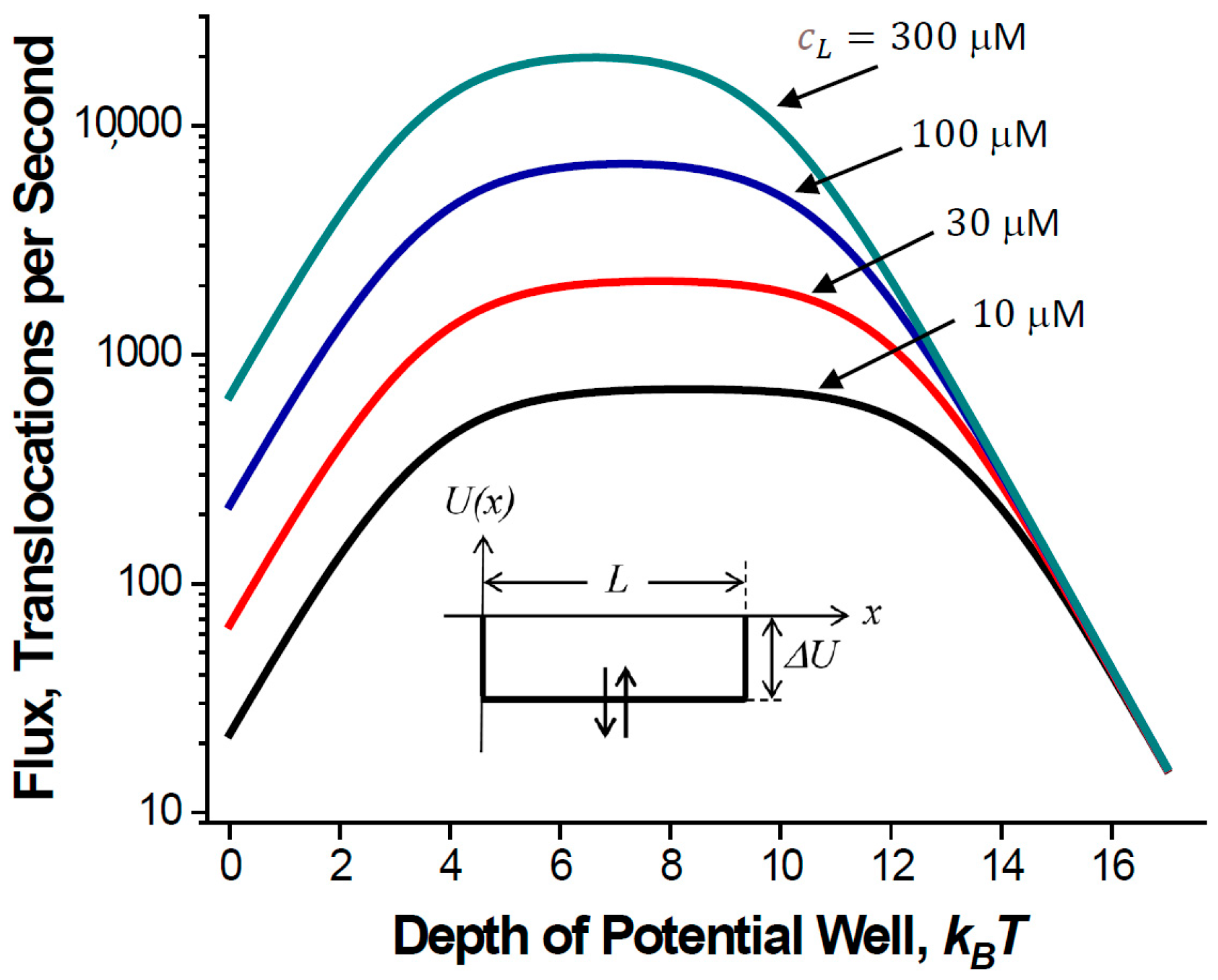
- Attractive interactions with the pore walls create a potential well inside the pore, which makes a particle spend more time in the channel but increases the particle flux.
- (ii).
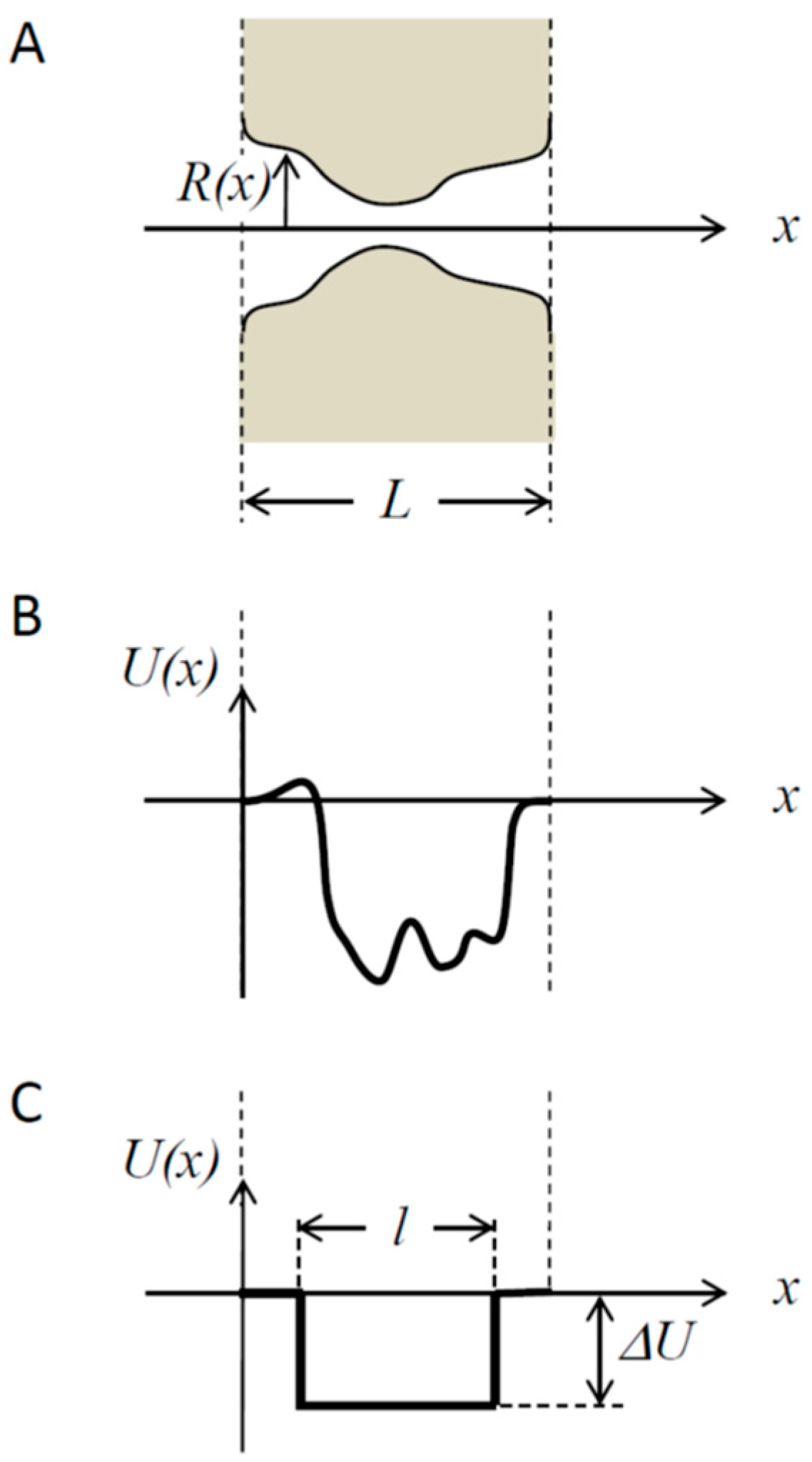
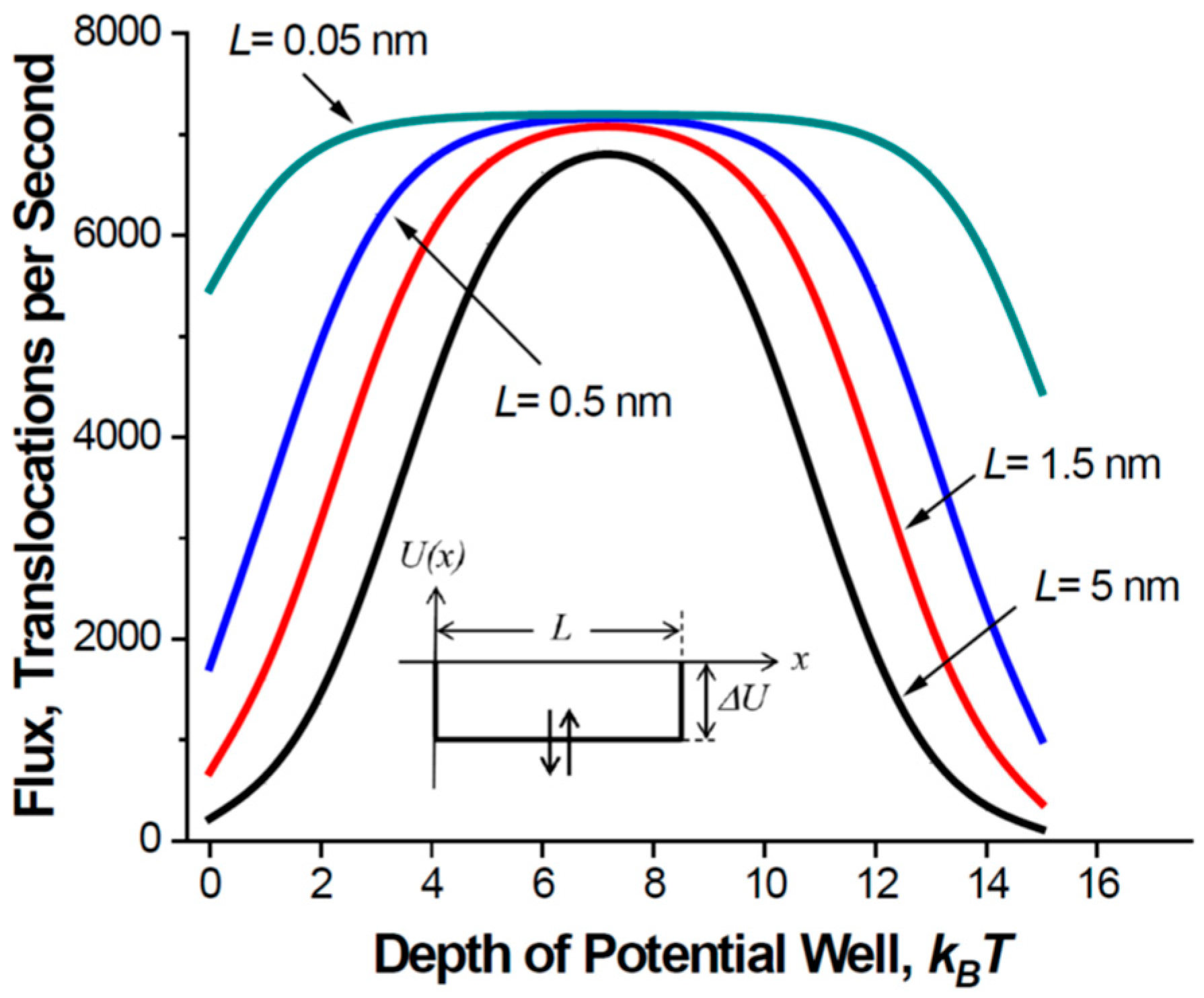
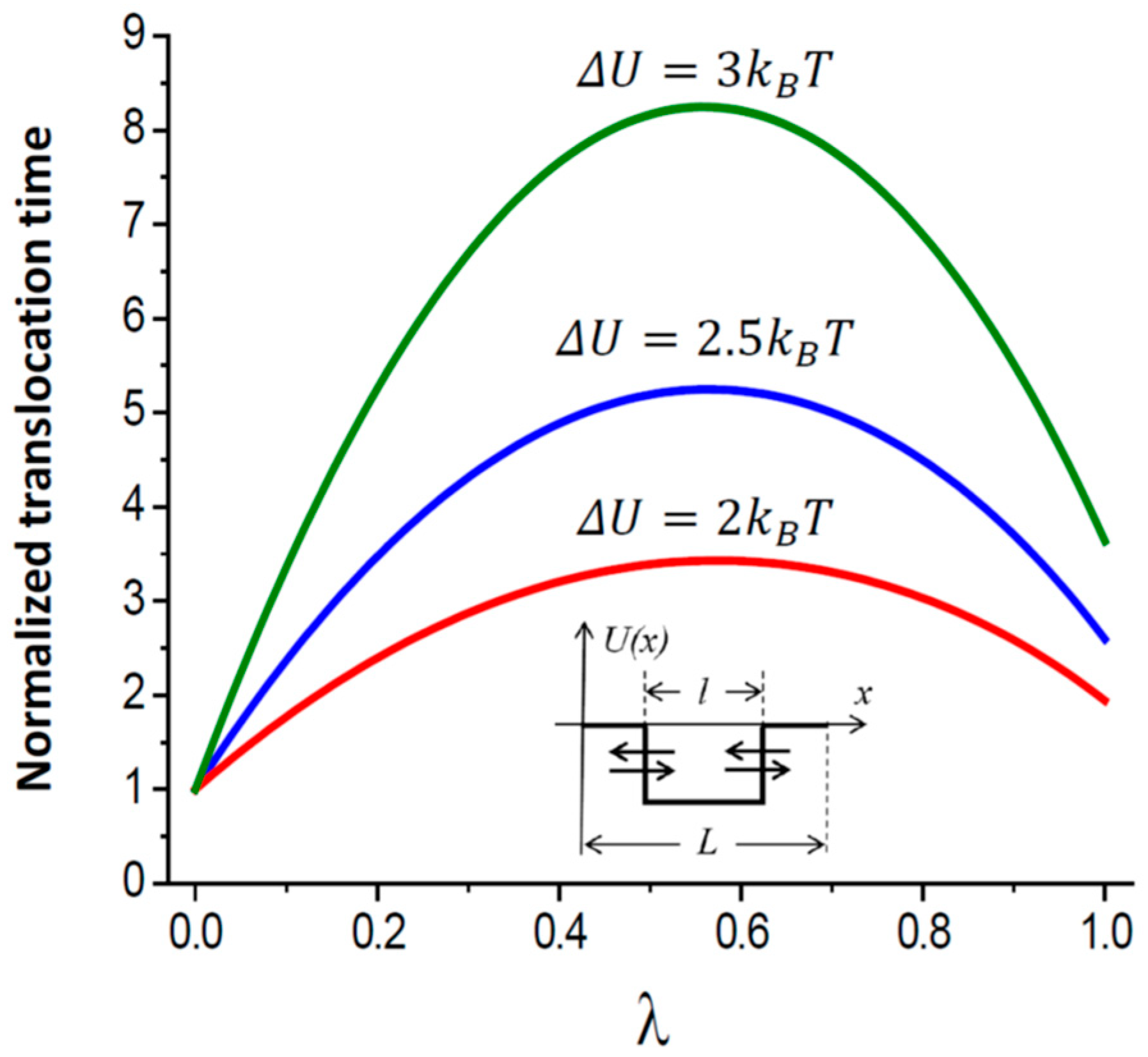
- If the potential well-describing particle-channel interaction occupies only part of the pore length, the mean translocation time is a non-monotonic function of the well width, first increasing and then decreasing with it.
- (iii).
- For a rectangular potential well occupying the entire pore, the mean residence time of a particle in the pore is independent of its intra-channel diffusivity and depends only on its diffusivity in the bulk.
- (iv).
- In the presence of a potential bias, the “downhill” particle flux is, as might be expected, higher than the “uphill” one; however, the mean particle downhill and uphill translocation times (as well as their distributions) are identical.
- (v).
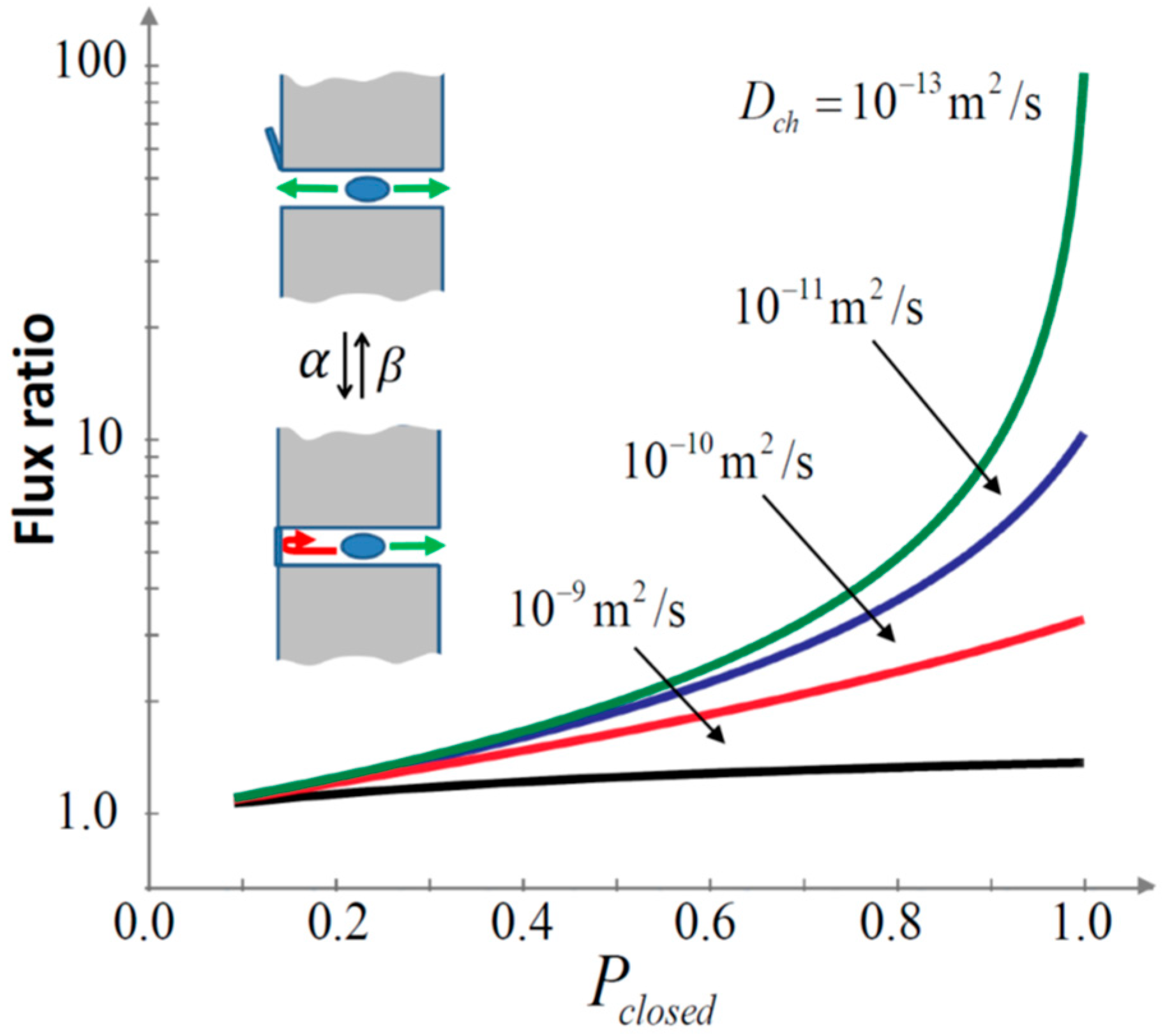
- Stochastic gating, if it is fast enough, provides a new selectivity mechanism that enhances relative transport rates of slower moving particles by orders of magnitude.
2. Results
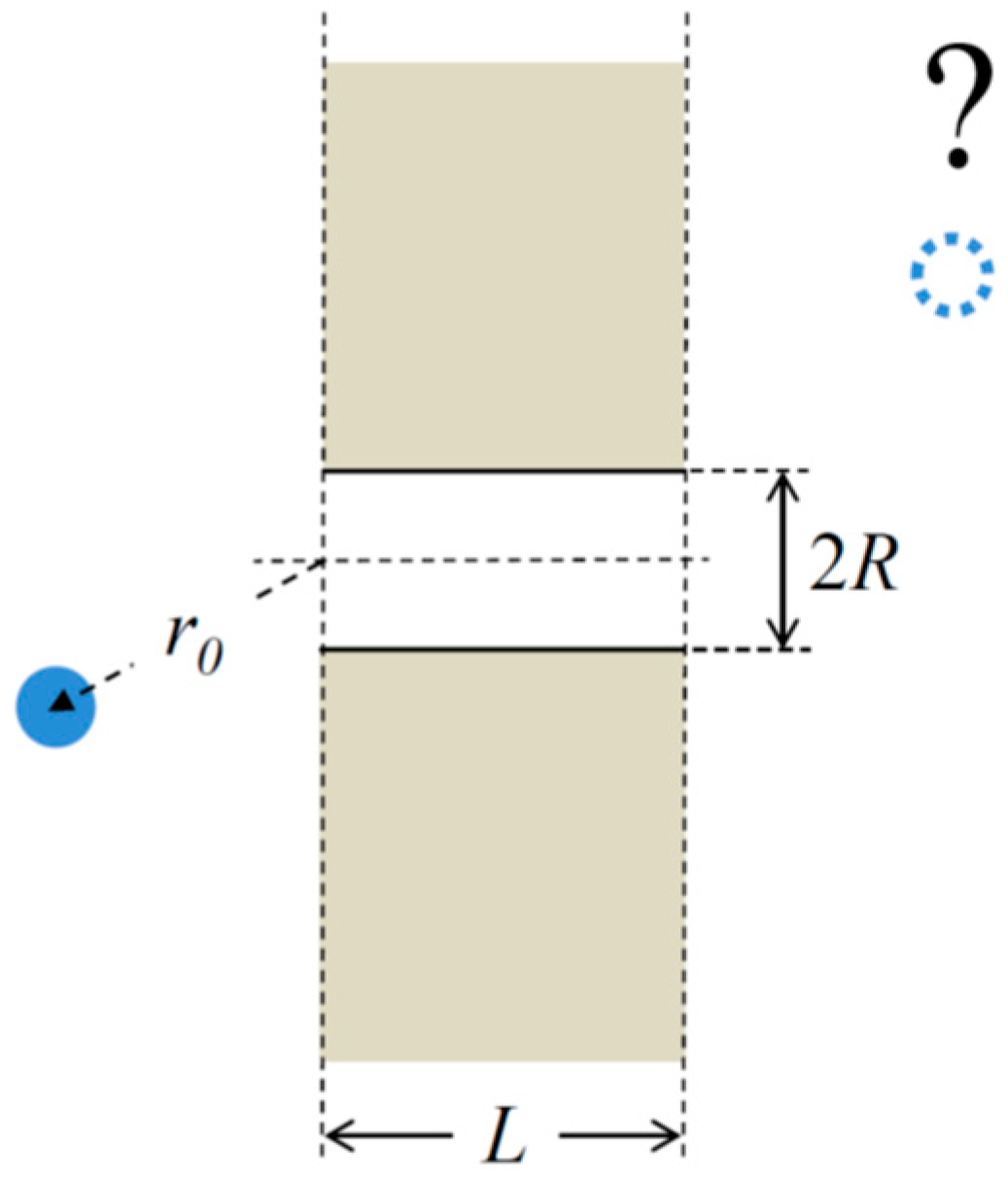
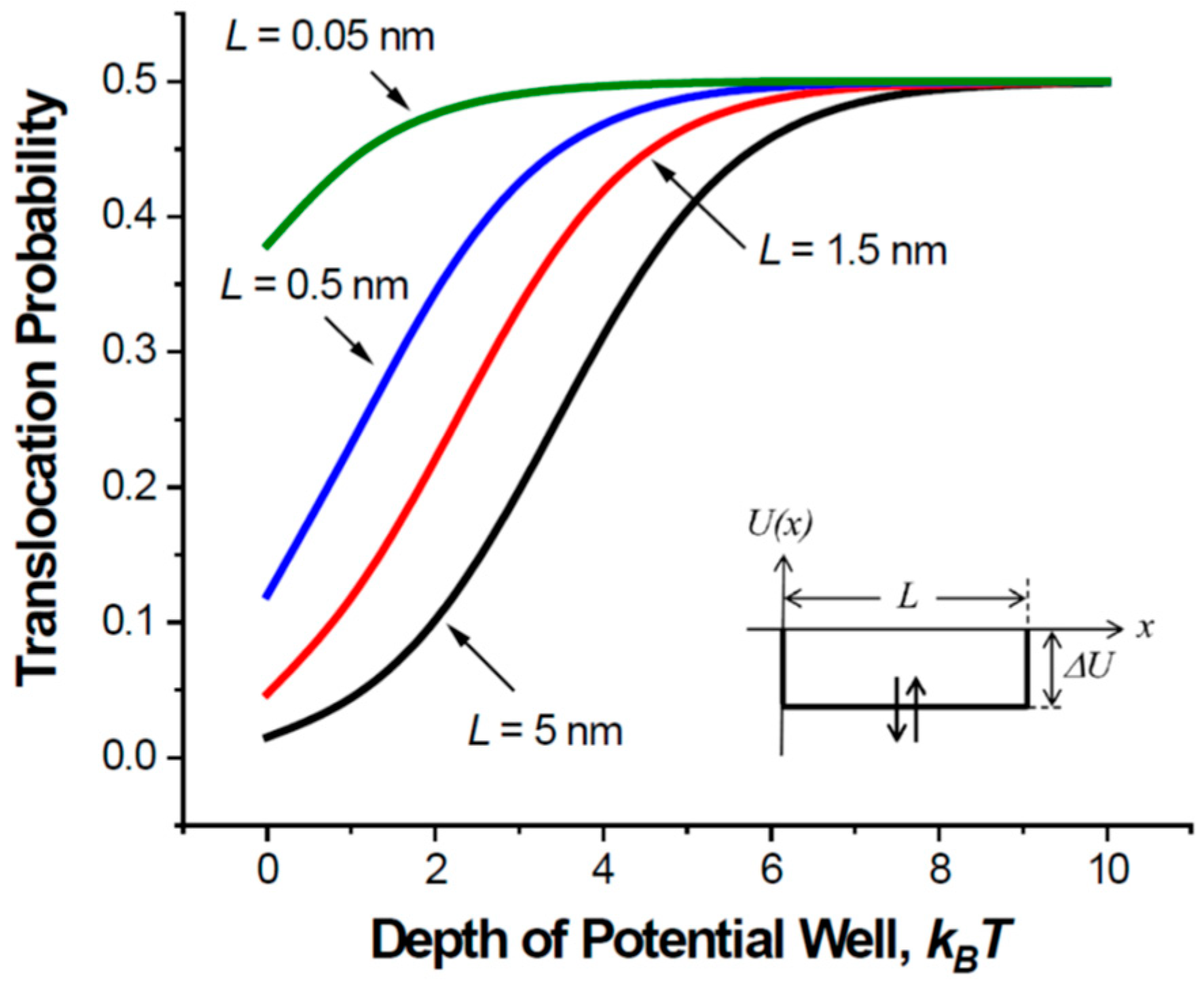
2.1. Translocation Probability and Steady-State Flux
2.2. Mean Translocation Time
2.3. Uphill and Downhill Translocations
2.4. Channel Gating as Selectivity Mechanism
3. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Andersen, C.; Jordy, M.; Benz, R. Evaluation of the Rate Constants of Sugar-Transport through Maltoporin (Lamb) of Escherichia-Coli from the Sugar-Induced Current Noise. J. Gen. Physiol. 1995, 105, 385–401. [Google Scholar] [CrossRef]
- Schirmer, T.; Keller, T.A.; Wang, Y.F.; Rosenbusch, J.P. Structural Basis for Sugar Translocation through Maltoporin Channels at 3.1-Angstrom Resolution. Science 1995, 267, 512–514. [Google Scholar] [CrossRef]
- Hilty, C.; Winterhalter, M. Facilitated substrate transport through membrane proteins. Phys. Rev. Lett. 2001, 86, 5624–5627. [Google Scholar] [CrossRef]
- Nestorovich, E.M.; Danelon, C.; Winterhalter, M.; Bezrukov, S.M. Designed to penetrate: Time-resolved interaction of single antibiotic molecules with bacterial pores. Prod. Natl. Acad. Sci. USA 2002, 99, 9789–9794. [Google Scholar] [CrossRef]
- Delcour, A.H. Solute uptake through general porins. Front. Biosci. 2003, 8, D1055–D1071. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. R. 2003, 67, 593–656. [Google Scholar]
- Vergalli, J.; Bodrenko, I.V.; Masi, M.; Moynie, L.; Acosta-Gutierrez, S.; Naismith, J.H.; Davin-Regli, A.; Ceccarelli, M.; van den Berg, B.; Winterhalter, M.; et al. Porins and small-molecule translocation across the outer membrane of Gram-negative bacteria. Nat. Rev. Microbiol. 2020, 18, 164–176. [Google Scholar]
- Winterhalter, M. Antibiotic uptake through porins located in the outer membrane of Gram-negative bacteria. Expert. Opin. Drug Del. 2021, 18, 449–457. [Google Scholar] [CrossRef]
- Masi, M.; Vergalli, J.; Ghai, I.; Barba-Bon, A.; Schembri, T.; Nau, W.M.; Lafitte, D.; Winterhalter, M.; Pages, J.M. Cephalosporin translocation across enterobacterial OmpF and OmpC channels, a filter across the outer membrane. Commun. Biol. 2022, 5, 1059. [Google Scholar]
- Chou, T. Kinetics and thermodynamics across single-file pores: Solute permeability and rectified osmosis. J. Chem. Phys. 1999, 110, 606–615. [Google Scholar] [CrossRef]
- Zilman, A. Effects of Multiple Occupancy and Interparticle Interactions on Selective Transport through Narrow Channels: Theory versus Experiment. Biophys. J. 2009, 96, 1235–1248. [Google Scholar]
- Zilman, A.; Pearson, J.; Bel, G. Effects of Jamming on Nonequilibrium Transport Times in Nanochannels. Phys. Rev. Lett. 2009, 103, 128103. [Google Scholar]
- Zilman, A.; Bel, G. Crowding effects in non-equilibrium transport through nano-channels. J. Phys.-Condens. Mat. 2010, 22, 454130. [Google Scholar] [CrossRef]
- Zilman, A.; Di Talia, S.; Jovanovic-Talisman, T.; Chait, B.T.; Rout, M.P.; Magnasco, M.O. Enhancement of Transport Selectivity through Nano-Channels by Non-Specific Competition. PLoS Comput. Biol. 2010, 6, 1000804. [Google Scholar]
- Bauer, W.R.; Nadler, W. Cooperative transport in nanochannels. Phys. Rev. E 2013, 88, 010703(R). [Google Scholar]
- Bauer, W.R. Mechanisms of cooperation and competition of two-species transport in narrow nanochannels. Phys. Rev. E 2017, 96, 062109. [Google Scholar]
- Davtyan, A.; Kolomeisky, A.B. Theoretical insights into mechanisms of channel-facilitated molecular transport in the presence of stochastic gating. J. Chem. Phys. 2019, 150, 124111. [Google Scholar]
- Bauer, W.R. Impact of Interparticle Interaction on Thermodynamics of Nano-Channel Transport of Two Species. Entropy 2020, 22, 376. [Google Scholar]
- Bezrukov, S.M.; Berezhkovskii, A.M.; Pustovoit, M.A.; Szabo, A. Particle number fluctuations in a membrane channel. J. Chem. Phys. 2000, 113, 8206–8211. [Google Scholar] [CrossRef]
- Berezhkovskii, A.M.; Pustovoit, M.A.; Bezrukov, S.M. Channel-facilitated membrane transport: Transit probability and interaction with the channel. J. Chem. Phys. 2002, 116, 9952–9956. [Google Scholar] [CrossRef]
- Berezhkovskii, A.M.; Pustovoit, M.A.; Bezrukov, S.M. Channel-facilitated membrane transport: Average lifetimes in the channel. J. Chem. Phys. 2003, 119, 3943–3951. [Google Scholar]
- Bauer, W.R.; Nadler, W. Molecular transport through channels and pores: Effects of in-channel interactions and blocking. Prod. Natl. Acad. Sci. USA 2006, 103, 11446–11451. [Google Scholar]
- Zilman, A.; Di Talia, S.; Chait, B.T.; Rout, M.P.; Magnasco, M.O. Efficiency, selectivity, and robustness of nucleocytoplasmic transport. PLoS Comput. Biol. 2007, 3, 1281–1290. [Google Scholar]
- Berezhkovskii, A.M.; Bezrukov, S.M. Intrinsic diffusion resistance of a membrane channel, mean first-passage times between its ends, and equilibrium unidirectional fluxes. J. Chem. Phys. 2022, 156, 071103. [Google Scholar]
- Parsegian, A. Energy of an Ion Crossing a Low Dielectric Membrane-Solutions to 4 Relevant Electrostatic Problems. Nature 1969, 221, 844–846. [Google Scholar]
- Cherstvy, A.G. Electrostatic screening and energy barriers of ions in low-dielectric membranes. J. Phys. Chem. B 2006, 110, 14503–14506. [Google Scholar]
- Bossa, G.V.; May, S. Integral Representation of Electrostatic Interactions inside a Lipid Membrane. Molecules 2020, 25, 3824. [Google Scholar]
- Noskov, S.Y.; Berneche, S.; Roux, B. Control of ion selectivity in potassium channels by electrostatic and dynamic properties of carbonyl ligands. Nature 2004, 431, 830–834. [Google Scholar]
- Parsegian, V.A. Van-Der-Waals Forces; Cambridge University Press: Cambridge, UK, 2006; p. 380. [Google Scholar]
- Queralt-Martín, M.; Verdía-Báguena, C.; Aguilella, V.M.; Alcaraz, A. Electrostatic Interactions Drive the Nonsteric Directional Block of OmpF Channel by La. Langmuir 2013, 29, 15320–15327. [Google Scholar]
- Ma, T.J.; Janot, J.M.; Balme, S. Dynamics of long hyaluronic acid chains through conical nanochannels for characterizing enzyme reactions in confined spaces. Nanoscale 2020, 12, 7231–7239. [Google Scholar]
- Tanyeri, M.; Ranka, M.; Sittipolkul, N.; Schroeder, C.M. A microfluidic-based hydrodynamic trap: Design and implementation. Lab. Chip 2011, 11, 1786–1794. [Google Scholar] [PubMed]
- Lu, B.; Hoogerheide, D.P.; Zhao, Q.; Zhang, H.B.; Zhipeng, T.P.; Yu, D.P.; Golovchenko, J.A. Pressure-Controlled Motion of Single Polymers through Solid-State Nanopores. Nano Lett. 2013, 13, 3048–3052. [Google Scholar]
- Lin, L.N.; Brandts, J.F.; Brandts, J.M.; Plotnikov, V. Determination of the volumetric properties of proteins and other solutes using pressure perturbation calorimetry. Anal. Biochem. 2002, 302, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Hoogerheide, D.P.; Albertorio, F.; Golovchenko, J.A. Escape of DNA from a Weakly Biased Thin Nanopore: Experimental Evidence for a Universal Diffusive Behavior. Phys. Rev. Lett. 2013, 111, 248301. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.E.; Lam, Y.C. Electroosmotic Flow Hysteresis for Fluids with Dissimilar pH and Ionic Species. Micromachines 2021, 12, 1031. [Google Scholar] [CrossRef]
- Veliz, L.A.; Toro, C.A.; Vivar, J.P.; Arias, L.A.; Villegas, J.; Castro, M.A.; Brauchi, S. Near-Membrane Dynamics and Capture of TRPM8 Channels within Transient Confinement Domains. PLoS ONE 2010, 5, e13290. [Google Scholar] [CrossRef]
- Mamonov, A.B.; Coalson, R.D.; Nitzan, A.; Kurnikova, M.G. The role of the dielectric barrier in narrow biological channels: A novel composite approach to modeling single-channel currents. Biophys. J. 2003, 84, 3646–3661. [Google Scholar] [CrossRef]
- Dagdug, L.; Berezhkovskii, A.M.; Bezrukov, S.M. Trapping of single diffusing particles by a circular disk on a reflecting flat surface. Absorbing hemisphere approximation. Phys. Chem. Chem. Phys. 2023, 25, 2035–2042. [Google Scholar] [CrossRef]
- Zwanzig, R. Diffusion Past an Entropy Barrier. J. Phys. Chem. 1992, 96, 3926–3930. [Google Scholar] [CrossRef]
- Berezhkovskii, A.M.; Bezrukov, S.M. On the applicability of entropy potentials in transport problems. Eur. Phys. J-Spec. Top. 2014, 223, 3063–3077. [Google Scholar] [CrossRef]
- Berezhkovskii, A.M.; Bezrukov, S.M. Capturing single molecules by nanopores: Measured times and thermodynamics. Phys. Chem. Chem. Phys. 2021, 23, 1610–1615. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.L. Effect of Rotation on Diffusion-Controlled Rate of Ligand-Protein Association. Prod. Natl. Acad. Sci. USA 1975, 72, 4918–4922. [Google Scholar] [CrossRef] [PubMed]
- Berg, H.C.; Purcell, E.M. Physics of Chemoreception. Biophys. J. 1977, 20, 193–219. [Google Scholar] [CrossRef] [PubMed]
- Berezhkovskii, A.M.; Bezrukov, S.M. Optimizing transport of metabolites through large channels: Molecular sieves with and without binding. Biophys. J. 2005, 88, L17–L19. [Google Scholar] [CrossRef]
- Kolomeisky, A.B. Channel-facilitated molecular transport across membranes: Attraction, repulsion, and asymmetry. Phys. Rev. Lett. 2007, 98, 048105. [Google Scholar]
- Rostovtseva, T.; Colombini, M. ATP flux is controlled by a voltage-gated channel from the mitochondrial outer membrane. J. Biol. Chem. 1996, 271, 28006–28008. [Google Scholar] [CrossRef]
- Garaj, S.; Hubbard, W.; Reina, A.; Kong, J.; Branton, D.; Golovchenko, J.A. Graphene as a subnanometre trans-electrode membrane. Nature 2010, 467, 190–193. [Google Scholar] [CrossRef]
- Pagliara, S.; Dettmer, S.L.; Keyser, U.F. Channel-Facilitated Diffusion Boosted by Particle Binding at the Channel Entrance. Phys. Rev. Lett. 2014, 113, 048102. [Google Scholar] [CrossRef]
- Bier, M.; Derenyi, I.; Kostur, M.; Astumian, R.D. Intrawell relaxation of overdamped Brownian particles. Phys. Rev. E 1999, 59, 6422–6432. [Google Scholar] [CrossRef]
- Berezhkovskii, A.M.; Hummer, G.; Bezrukov, S.M. Identity of distributions of direct uphill and downhill translocation times for particles traversing membrane channels. Phys. Rev. Lett. 2006, 97, 020601. [Google Scholar] [CrossRef]
- Redner, S. A Guide to First-Passage Processes, 1st ed.; Cambridge University Press: Boston, MA, USA, 2007. [Google Scholar]
- Alvarez, J.; Hajek, B. Equivalence of trans paths in ion channels. Phys. Rev. E 2006, 73, 046126. [Google Scholar] [CrossRef]
- Chung, H.S.; Eaton, W.A. Protein folding transition path times from single molecule FRET. Curr. Opin. Struct. Biol. 2018, 48, 30–39. [Google Scholar] [CrossRef]
- Eaton, W.A. Modern Kinetics and Mechanism of Protein Folding: A Retrospective. J. Phys. Chem. B 2021, 125, 3452–3467. [Google Scholar] [CrossRef]
- Berezhkovskii, A.M.; Bezrukov, S.M. Stochastic Gating as a Novel Mechanism for Channel Selectivity. Biophys. J. 2018, 114, 1026–1029. [Google Scholar] [CrossRef] [PubMed]
- Hille, B. Ion Channels in Excitable Membranes; Sinauer Associates: Sunderland, MA, USA, 2001. [Google Scholar]
- Colombini, M. VDAC structure, selectivity, and dynamics. BBA-Biomembr. 2012, 1818, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Ujwal, R.; Cascio, D.; Colletier, J.P.; Faham, S.; Zhang, J.; Toro, L.; Ping, P.P.; Abramson, J. The crystal structure of mouse VDAC1 at 2.3 angstrom resolution reveals mechanistic insights into metabolite gating. Prod. Natl. Acad. Sci. USA 2008, 105, 17742–17747. [Google Scholar] [CrossRef] [PubMed]
- Colombini, M. The published 3D structure of the VDAC channel: Native or not? Trends Biochem. Sci. 2009, 34, 382–389. [Google Scholar] [CrossRef]
- Goncalves, R.P.; Buzhysnskyy, N.; Scheuring, S. Mini review on the structure and supramolecular assembly of VDAC. J. Bioenerg. Biomembr. 2008, 40, 133–138. [Google Scholar] [CrossRef]
- Rostovtseva, T.; Colombini, M. VDAC channels mediate and gate the flow of ATP: Implications for the regulation of mitochondrial function. Biophys. J. 1997, 72, 1954–1962. [Google Scholar] [CrossRef]
- Lee, A.C.; Xu, X.F.; Colombini, M. The role of pyridine dinucleotides in regulating the permeability of the mitochondrial outer membrane. J. Biol. Chem. 1996, 271, 26724–26731. [Google Scholar] [CrossRef]
- Simson, P.; Jepihhina, N.; Laasmaa, M.; Peterson, P.; Birkedal, R.; Vendelin, M. Restricted ADP movement in cardiomyocytes: Cytosolic diffusion obstacles are complemented with a small number of open mitochondrial voltage-dependent anion channels. J. Mol. Cell Cardiol. 2016, 97, 197–203. [Google Scholar] [CrossRef]
- Bean, R.C.; Shepherd, W.C.; Chan, H.; Eichner, J. Discrete Conductance Fluctuations in Lipid Bilayer Protein Membranes. J. Gen. Physiol. 1969, 53, 741–757. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. A Quantitative Description of Membrane Current and Its Application to Conduction and Excitation in Nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef]
- Jackson, M.B. Molecular and Cellular Biophysics; Cambridge University Press: Cambridge, UK, 2006; pp. 1–512. [Google Scholar]
- Hoogerheide, D.P.; Gurnev, P.A.; Rostovtseva, T.K.; Bezrukov, S.M. Voltage-activated complexation of alpha-synuclein with three diverse beta-barrel channels: VDAC, MspA, and alpha-hemolysin. Proteomics 2021, 22, 2100060. [Google Scholar] [CrossRef]
- Astumian, R.D.; Hanggi, P. Brownian motors. Phys. Today 2002, 55, 33–39. [Google Scholar] [CrossRef]
- Hanggi, P.; Marchesoni, F. Artificial Brownian motors: Controlling transport on the nanoscale. Rev. Mod. Phys. 2009, 81, 387–442. [Google Scholar] [CrossRef]
- Archana, G.R.; Barik, D. Roughness in the periodic potential induces absolute negative mobility in a driven Brownian ratchet. Phys. Rev. E 2022, 106, 044129. [Google Scholar]
- Ehrenstein, G.; Lecar, H.; Nossal, R. Nature of Negative Resistance in Bimolecular Lipid Membranes Containing Excitability-Inducing Material. J. Gen. Physiol. 1970, 55, 119–133. [Google Scholar] [CrossRef]
- Bezanilla, F.; Armstrong, C.M. Gating Currents of Sodium Channels—3 Ways to Block Them. Science 1974, 183, 753–754. [Google Scholar] [CrossRef]
- Bezanilla, F. How membrane proteins sense voltage. Nat. Rev. Mol. Cell Biol. 2008, 9, 323–332. [Google Scholar] [CrossRef]
- Swartz, K.J. Sensing voltage across lipid membranes. Nature 2008, 456, 891–897. [Google Scholar] [CrossRef]
- Mandala, V.S.; MacKinnon, R. Voltage-sensor movements in the Eag Kv channel under an applied electric field. Prod. Natl. Acad. Sci. USA 2022, 119, e2214151119. [Google Scholar] [CrossRef]
- Mierle, G. The Effect of Cell-Size and Shape on the Resistance of Unstirred Layers to Solute Diffusion. Biochim. Biophys. Acta 1985, 812, 835–839. [Google Scholar] [CrossRef]
- Hall, J.E. Access Resistance of a Small Circular Pore. J. Gen. Physiol. 1975, 66, 531–532. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berezhkovskii, A.M.; Bezrukov, S.M. Counter-Intuitive Features of Particle Dynamics in Nanopores. Int. J. Mol. Sci. 2023, 24, 15923. https://doi.org/10.3390/ijms242115923
Berezhkovskii AM, Bezrukov SM. Counter-Intuitive Features of Particle Dynamics in Nanopores. International Journal of Molecular Sciences. 2023; 24(21):15923. https://doi.org/10.3390/ijms242115923
Chicago/Turabian StyleBerezhkovskii, Alexander M., and Sergey M. Bezrukov. 2023. "Counter-Intuitive Features of Particle Dynamics in Nanopores" International Journal of Molecular Sciences 24, no. 21: 15923. https://doi.org/10.3390/ijms242115923
APA StyleBerezhkovskii, A. M., & Bezrukov, S. M. (2023). Counter-Intuitive Features of Particle Dynamics in Nanopores. International Journal of Molecular Sciences, 24(21), 15923. https://doi.org/10.3390/ijms242115923