1. Introduction
In the growth process of aquatic organisms, they are constantly exposed to reactive oxygen species (ROS) and electrophilic substances produced through metabolic and environmental factors [
1,
2]. These threats can profoundly impact the development and growth of organisms, potentially resulting in tissue damage, senilism, and cell apoptosis. To effectively respond to and counteract these pressures, cells activate multiple defense systems that are closely linked to various cellular processes [
3,
4,
5]. Transcription factors are one of the important contributors to this system, regulating the expression of genes involved in cellular protection [
4,
6]. One such transcription factor is nuclear factor erythroid 2-related factor 2 (Nrf2), which belongs to the cap’n’collar (CNC) alkaline region leucine zip (BZip) family. Nrf2 plays a significant role in inducing the expression of cell protection genes by directly interacting with antioxidant response elements (ARE) [
5,
7]. Its downstream target genes include genes encoding antioxidant and detoxification enzymes such as glutathione S-transferase (GST), heme oxygenase-1 (HO-1), quinone oxidoreductase 1 (NQO1), and aldo keto reductase (AKR). These enzymes play a crucial role in the elimination of ROS, xenobiotic metabolism, and detoxification.
The role of Nrf2 in aquatic organisms has been extensively investigated, shedding light on its significance in maintaining antioxidant defense mechanisms. Research has demonstrated benzo[a]pyrene (Bap) irritation induced the expression of Nrf2 and a series of antioxidant enzymes, such as superoxide dismutase (SOD), glutathione peroxidase (GPX), catalase (CAT), and glutathione reductase (GR), in
Mytilus coruscus [
7]. Similarly, when
Scylla paramamosain was exposed to stress from
Vibrio parahaemolyticus, there was a notable increase in the expression levels of
Nrf2 and other antioxidant genes, including
HO-1 and
NQO-1 [
8]. In
Litopenaeus vannamei, the knockdown of
Nrf2 gene using dsRNA-mediated techniques resulted in inhibited antioxidant genes, reduction of antioxidant enzyme activity, upregulated expression of apoptosis and autophagy genes, and noticeable alterations in tissue structure [
9]. These findings underscore the crucial role of Nrf2 in orchestrating the response to external stressors in aquatic animals.
Nrf2 functions in cellular transcription by forming heterodimers with members of the small Maf family, including MafG, MafK, and MafF [
10]. Through its BZip domain, Nrf2 establishes these heterodimeric complexes, enabling it to regulate gene expression [
11,
12]. When cells are exposed to electrophilic agents or ROS, Nrf2 is released from its binding partner, the cytoplasmic protein Keap1, triggering its translocation into the cell nucleus. Once inside the nucleus, Nrf2 forms heterodimers with small Maf, leading to their association with ARE or electrophilic response elements. Remarkable advancements have been made in the study of the small Maf family, particularly with the successful isolation and cloning of
MafG and
MafK genes from human hematopoietic cells [
12,
13]. Subsequent investigations have revealed that MafG and MafK can form heterodimers not only with Nrf2 but also with Nrf1. These heterodimeric complexes play a crucial role in binding to the nuclear erythroid factor 2 (NF-E2) site, further highlighting their regulatory function [
14]. Extensive research on the genuine partner molecules involved in supporting
Nrf2 activity in vivo has shown that the absence of small
Maf can effectively reverse the cellular dysfunction caused by
Keap1 gene deficiency. Furthermore, it has been observed that this absence also leads to a decrease in the mortality rate in mice resulting from
Keap1 loss [
10]. These discoveries emphasize the critical collaboration between small Maf and Nrf2 within the organism. Moreover, the absence of
Nrf2 and the lack of small
Maf both result in highly similar and severe damage in inducing the response of antioxidant and exogenous metabolic enzyme genes to electrophilic agents [
15,
16]. This further underscores the pivotal role played by Nrf2 and small Maf as heterodimers operating within the intricate biological system. Mice lacking small
Maf exhibit liver steatosis and gene dysregulation related to lipid and amino acid metabolism, as well as proteasome subunit expression. At the same time, the expression levels of many Nrf2 target genes also decrease [
17].
Presently, there have been reports of the successful cloning of small
Maf genes in aquatic organisms such as
Danio rerio,
Cristaria plicata and
Procambarus clarkia. Takagi et al. [
18] identified a novel small Maf protein known as MafT in
Danio rerio. Further experiments revealed that the co-overexpression of
MafT and
Nrf2 resulted in synergistic activation of MARE-mediated gene expression in
Danio rerio embryos [
18]. In another study, Wang identified
MafK in
Cristaria plicata, confirming the critical detoxification role of
CpMafK in microcystin toxin stress [
19]. In
Procambarus clarkii, the transcriptional expression of the
PcMafG-like gene and certain antioxidant genes in the hepatopancreas and gills was significantly up-regulated under Cu
2+/Cd
2+ stimulation [
20]. However, when
PcMafG-like was interfered with using dsRNA, the expression of antioxidant genes was inhibited, leading to more severe pathological damage. These findings corroborate the potential of small
Mafs in eliciting the activation of antioxidant genes in aquatic organisms. Nevertheless, there remains a dearth of research concerning the involvement of small Mafs in the thick shell mussel
M. coruscus, a crucial model organism in marine environmental studies.
M. coruscus is primarily distributed in the Yellow Sea and East China Sea, adopting a lifestyle of attachment and filter feeding [
21,
22]. As they inhabit coastal areas prone to pollution, pollutants accumulate in various tissues, potentially leading to disrupted ROS metabolism [
23]. This has profound implications for their survival, growth, development, and evolution, emphasizing the necessity of researching oxidative stress responses in
M. coruscus. In our previous study, we found that when
M. coruscus was exposed to acute Bap stress, the Nrf2-dependent antioxidant system was activated at both the transcriptional and enzyme levels [
7]. This activation serves as a protective mechanism to counteract the toxic effects of Bap. However, the underlying mechanisms of this potential action, including how small Mafs function, are still unclear. In this study, we focused on investigating the significance of the small
Maf gene in the Nrf2 pathway and its crucial functional role in the antioxidant process of mussel
M. coruscus.
3. Discussion
The Nrf2 pathway, as the key defense system against environmental damage and regulator of body homeostasis, has emerged as a critical research focus. Despite its significance, the understanding of the molecular mechanisms of Maf in aquatic organisms is presently limited. In light of this, our study aims to address this knowledge gap through the identification and characterization of Maf in
M. coruscus. Our findings revealed that
McMafF_G_K, an important Maf variant, contained a DNA-binding domain and the leucine zipper structures that play a crucial role in both self and other BZip transcription factors’ dimerization [
24,
25]. Notably,
McMafF_G_K also possesses the HER domain, a universally observed domain in all Mafs. This conserved structural domain facilitates stable DNA binding [
26]. These findings substantiate the categorization of the presently identified Maf molecule within the Maf family while illuminating the wider evolutionary backdrop of Mafs.
The expression profiles of small
Mafs were comprehensively characterized in various mouse tissues, yielding valuable insights into their differential gene expression patterns. Specifically,
MafK and
MafF exhibited prominent upregulation in the lung, underscoring their vital roles in pulmonary physiology, whereas
MafG displayed pronounced abundance in the heart, highlighting its significance in cardiac function [
27]. Furthermore, during hypercapnic stimulation in rats,
MafG mRNA was discernible not only in the heart but also in diverse tissues, including skeletal muscle, cerebral cortex, cerebellum, liver, stomach, and intestine, suggesting its potential involvement in multiple physiological processes in response to this stimulus [
28]. Intriguingly, an exploration of
Maf gene expression in zebrafish unveiled a widespread distribution of small
Mafs across different tissues, with the brain exhibiting particularly elevated expression levels, emphasizing the significance of these proteins in neural functions [
18]. In
Procambarus clarkia, the presence of
PcMafG-like mRNA was detected in all examined tissues, with muscle tissue exhibiting the highest expression levels, likely attributable to the integral role of muscle tissue in the organism’s biology [
20]. Additionally, the present study revealed the constitutive expression of
McMafF_G_K mRNA in all examined tissues, with heightened expression levels observed in digestive glands, gills, and hemocytes, all recognised as immune-associated tissues in bivalves, implying their potential contributions to immune responses. These comprehensive analysis of small
Maf gene expression profiles in different tissues provides valuable insights into their differential regulation and tissue-specific significance. These findings contribute to our understanding of the multiple physiological roles played by small Mafs.
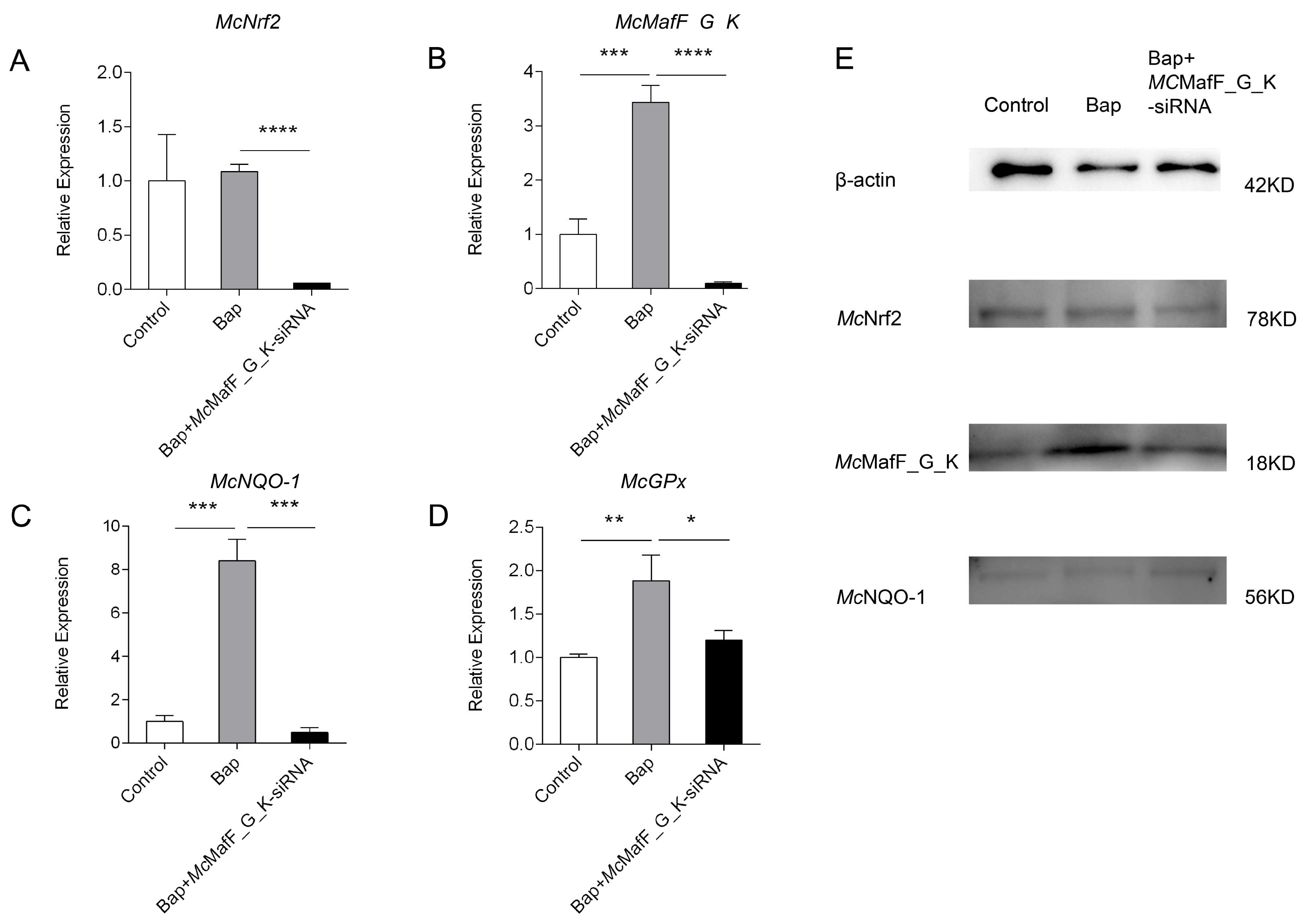
Small Mafs play a crucial role as transcription factors in the activation of ARE-dependent genes, which are responsible for cellular protection, effectively shielding the organism from environmental harm, and this is essential for
Nrf2-mediated gene activation. Research has shown that the expression of ARE-dependent genes in the liver of mice is influenced to varying degrees and exhibits high sensitivity to oxidative stress responses. Inducers such as hydrogen peroxide, ARE or electrophile responsive element (ARE/EpRE) inducers, and hypercapnia (elevated carbon dioxide levels) can stimulate the induction of small Mafs, activating the expression of ARE-dependent cell protective genes [
16,
28,
29]. In a study investigating whether inducers can also activate the expression of zebrafish small Mafs, it was found that diethylmaleate (DEM) can induce the expression of
MafT and
MafG1. Additionally, the co-expression of
MafT and
Nrf2 synergistically activates gene expression mediated by MARE in zebrafish [
18]. Upon Cu
2+/Cd
2+ stimulation, the expression of
PcMafG-like and downstream antioxidant genes is upregulated in the hepatopancreas and gills of
Procambarus clarkia [
20]. During this study, we observed an upregulation in the expression levels of
McMafK_G_F,
McNrf2, and downstream target genes
McGPx and
McNQO-1 in the digestive gland cells of
M. coruscus following a six-hour Bap exposure. At the same time, we used qPCR technology to assess gene expression after
McMafF_G_K gene interference. The results showed significant downregulation in the expression levels of
McMafK_G_F,
McNrf2,
McNQO-1, and
McGpx compared to the Bap stimulation group. Specialised research into
Mafs regulation unveiled that mice embryos with reduced
Maf expression exhibited a decreased basal expression of ARE-dependent cellular protective genes. This decreased expression of oxidative stress response genes may exacerbate cell apoptosis, embryonic growth retardation, and impaired liver function [
30]. To clarify the precise contributions of small Mafs to the functions of various CNC proteins, a mouse strain lacking small Mafs in the liver was established. It was found that the livers of small Maf-deficient mice shared a similar expression profile with those of Nrf1 and Nrf2-deficient mice. This was characterised by a reduction in the expression levels of Nrf2 target genes, along with liver fat degeneration and gene dysregulation related to lipid and amino acid metabolism, as well as proteasome subunits [
17]. These findings are consistent with the results of this study, indicating that Mafs, as central regulators of the Nrf2 signaling pathway, are indispensable in the process of regulating the basal expression of ARE-dependent cellular protective genes during oxidative stress.
The continuous generation of ROS is a common response in aquatic animals when facing external stressors. Both non-biological water pollution and biological factors can induce the excessive accumulation of ROS in marine organisms, leading to oxidative stress. In our previous study, we observed that exposure to Bap significantly increased the activities of SOD, CAT, GPx, and GR enzymes in the digestive glands of
M. coruscus. This suggests that upon exposure to Bap, the antioxidant defense system is triggered to counteract oxidative stress, effectively protecting cells from BaP-induced oxidative damage [
7]. Huang et al. [
9] found that injecting Nrf2 dsRNA into
Litopenaeus vannamei resulted in decreased activities of SOD, CAT, and GPx enzymes, while malondialdehyde (MDA) activity increased. Similarly, Wang et al. [
31] demonstrated that silencing Nrf2 for 72 h using RNA interference in the digestive gland of
Cristaria plicata led to significantly higher levels of MDA, indicating elevated lipid peroxidation compared to the control group. In the binding and recognition process of the Nrf2/Maf heterodimer for activating the antioxidant stress response, Nrf2 and Maf play a crucial synergistic role. Interfering with Nrf2 and Maf may lead to similar results [
10,
32]. Additionally, it was observed that when discussing the significant role of MafG in the
Procambarus clarkii oxidative response, the activities of GSH, Cu/Zn-superoxide dismutase (CZ-SOD), and CAT significantly increased in the Cu
2+/Cd
2+ stimulation group, while interference with
PcMafG-like led to a reduction in enzyme activities [
20]. In this study, Bap stimulation resulted in a significant increase in T-AOC and ROS activities, indicating that Bap disrupts the redox balance within the organism, leading to the accumulation of ROS.
M. coruscus alleviates the damage caused by ROS accumulation by enhancing its total antioxidant capacity. However,
McMafF_G_K siRNA interference resulted in increased ROS activity and decreased T-AOC activity, suggesting that Bap activates the Nrf2 signaling pathway to mitigate oxidative stress, and silencing MafF_G_K disrupts the normal antioxidant mechanisms, resulting in decreased total antioxidant capacity and elevated ROS levels. These results suggest that small Mafs may play a crucial regulatory role in the antioxidative stress process of bivalves, and their functional loss could potentially exacerbate damage to the organisms.
Small Mafs and Nrf2 belong to the CNC transcription factor family, both possessing a BZip structure that mediates DNA binding and dimerization [
15,
24,
25]. The Nrf2-small Maf heterodimer plays a crucial role in oxidative stress responses by transcriptionally activating a multitude of cell-protective genes through antioxidant response elements [
33]. Surface plasmon resonance microarray imaging technology has been employed to demonstrate the high affinity of the Nrf2-Maf heterodimer for MARE-like elements [
33]. In the context of the regulation of
C. plicata Prx5 within the Nrf2/ARE signaling pathway, mass spectrometry analysis has identified MafK protein as one of the interactors with Nrf2-BZip, which was validated through yeast two-hybrid experiments [
5]. Herein, we present the successful construction of recombinant vectors His-
McMafF_G_K and GST-
McNrf2, as well as the expression of the recombinant proteins in vitro. Our GST-pull down experiments have confirmed the interaction between
McNrf2 and
McMafF_G_K, thus highlighting the formation of heterodimers between
McMafF_G_K and
McNrf2 that participate in the transcriptional regulation of antioxidant stress responses.
4. Materials and Methods
4.1. Animals
A total of 200 M. coruscus individuals were collected from Donghe Market in Zhoushan city, Zhejiang province. They were then temporarily cultured for a week in seawater with a temperature of 25 ± 1 °C and salinity of 30‰. During the domestication process, 50% of the seawater was replaced daily, and spirulina powder was fed to them every day.
4.2. Silicon Cloning for McMafF_G_K
The sequence information of
McMafF_G_K cDNA was silicon cloned from the National Center for Biotechnology Information (NCBI) (accession number: CAC5373282). A specific primer pair (
Table 1) was designed to amplify the sequence of the
McMafF_G_K open reading frame (ORF) region. After DNA sequencing of the PCR products, the putative amino acid sequence was presumed. Theoretical calculations of the isoelectric point and predicted molecular weight of
McMafF_G_K were carried out utilizing the EXPASY platform (V3.0,
http://web.expasy.org, accessed on 24 September 2023). To predict the presence of conserved domains, we employed the SMART tool (V3.0,
http://smart.embl-heidelberg.de/, accessed on 15 October 2023). Homologous amino acid sequences of
McMafF_G_K were sourced from NCBI utilizing their Blast service (
https://www.ncbi.nlm.nih.gov/blast, accessed on 13 October 2023).
4.3. Bap Exposure
The Bap exposure assay was conducted following the methodology described in our previous publication [
7]. In brief, 15 mussels assigned to the experimental cohorts were subjected to a 50 μg/L concentration of Bap, while 3 in the control cohort were solely exposed to the vehicle DMSO at an equivalent dosage. Subsequent to stimulation, digestive glands were dissected at multiple time intervals (0, 12, 24, 36, 72, and 96 h). To minimize the impact of individual variability, digestive gland samples from three mussels were pooled together as a single replicate, and three replicates were conducted for each time point. Additionally, six different tissues, including the adductor muscle, gills, mantle, gonad, hemocytes, and digestive glands, were extracted from nine untreated, healthy mussels to investigate the tissue distribution of
McMafF_G_K. 4.4. Cell Transfection
Digestive gland tissue weighing 0.3–0.35 g was carefully disinfected by immersing it in 25 mL of Hank’s solution (Solarbio, Shanghai, China) supplemented with 1% penicillin-streptomycin-gentamicin solution (Solarbio, Shanghai, China). Thereafter, the tissue was thoroughly rinsed with PBS and finely minced using surgical scissors. To facilitate digestion, 8 mL of 0.25% trypsin solution (Solarbio, Shanghai, China) was added and allowed to react for 20 min with vigorous shaking and stirring every 5 min. After the hydrolysis was terminated, the resulting filtrate was collected and centrifuged at 3000 rpm for 15 min at 4 °C. Subsequently, the cell precipitate was suspended in L-15 medium (Solarbio, Shanghai, China) containing 10% fetal bovine serum (Solarbio, Shanghai, China). The resultant solution was evenly distributed onto a six-well plate and cultured in a cell incubator at 28 °C. Transfection with McMafF_G_K-siRNA using GP-Transfect-Mate (Genepharma, Shanghai, China) was performed when the cell density reached 60% to 80%. To evaluate the impact of McMafF_G_K on the Nrf2 pathway, the cells were then exposed to Bap (50 μmol/mL) for a period of 6 h.
4.5. qRT-PCR
qRT-PCR assays were performed conventionally. Total RNA was extracted from the tissues and cells using Trizol (Invitrogen, Carlsbad, CA, USA) following established protocols. The extracted RNA was subsequently converted into cDNA using the Go Script
TM Reverse Transcription System kit (Promega, Madison, WI, USA). For gene detection, the SYBR Green Master Mix, included in the TB Green
®Fast qPCR Mix Kit (TaKaRa, Kusatsu, Japan), was employed on the 7500 system. The experiment utilized specific primers, as detailed in
Table 1. Each sample was analyzed in triplicate, and gene transcription levels were quantified using the 2
−ΔΔCt method. Finally, data were statistically analyzed and visualized using SPSS 19.0 and GraphPad Prism 8.0.
4.6. Western Blotting
The Western blot analysis followed the protocol outlined in our previous study [
34]. Initially, cultured cells were lysed using RIPA buffer (Beyotime, Shanghai, China), and the protein concentration was quantified using a BCA protein assay kit (Beyotime, Shanghai, China). Subsequently, 20 μg of total protein was isolated and separated on 8% and 12% SDS-PAGE gels and then transferred to a PVDF membrane. After blocking with 5% skim milk at room temperature for 1 h, the membrane was incubated overnight at 4 °C with primary antibodies (HuaBio, Hangzhou, China, 1:500). Following three washes, the membrane was incubated with a horseradish peroxidase-conjugated secondary antibody (ABclonal, Wuhan, China, 1:500). The target protein bands were visualized using the enhanced chemiluminescence (ECL) system (Beyotime, Shanghai, China) and subsequently normalized to β-actin.
4.7. McMafF_G_K Recombinant
The prokaryotic expression system pet28-McMafF_G_K was successfully transformed into Escherichia coli BL21 host cells (Qingke, Beijing, China) to initiate protein expression. To induce the expression of His-McMafF_G_K protein, the transformed cells were incubated at 37 °C for 4 h with 0.2 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) (Sangon Biotech, Shanghai, China). Similarly, the plasmid pGEX-4T-1-McNrf2 was transformed into TSsetta (DE3) (Qingke, Beijing, China) and allowed to culture overnight at 25 °C. The induction of GST-McNrf2 recombinant protein expression was carried out using 0.2 mM IPTG (Sangon Biotech, Shanghai, China). Following cell culture, the culture medium was centrifuged to remove the supernatant. The bacterial cells in suspension were then subjected to sonication for disruption, followed by centrifugation to separate the supernatant from the pellet. Subsequently, the protein samples were verified through 8% and 12% SDS-polyacrylamide gel electrophoresis. The purification of His-McMafF_G_K and GST-McNrf2 proteins was achieved by utilizing Ni-affinity resin and glutathione affinity resin (Huiyan Bio, Wuhan, China), respectively. Solution exchange was performed using Amicon@Ultra-15 ultrafiltration tubes (Millipore, Rockland, MA, USA). Finally, to prevent protein degradation, the His-McMafF_G_K and GST-McNrf2 proteins were stored in BupTM Tris-buffered saline (TBS) (Thermo Fisher Scientific, San Jose, CA, USA) supplemented with 1 mM phenylmethylsulfonyl fluoride (PMSF) (Sangon Biotech, Shanghai, China).
4.8. GST Pull Down
In the GST-pull down experiment, 100 μg of GST-McNrf2 protein was initially combined with glutathione agarose resin and gently oscillated at 4 °C for 1 h. Following this, 150 μg of His-McMafF_G_K protein was added to the GST-McNrf2 solution, and the mixture was gently oscillated at 4 °C for 2 h. The eluate was collected and then heated at 95 °C for 5 min for subsequent Western blotting analysis. For Western blotting, protein separation was performed using a 4–20% linear gradient SDS-polyacrylamide gel (GenScript ProBio, Billerica, MA, USA). After electrophoresis, the proteins were transferred onto a membrane using a standard transfer protocol. The membrane was then subjected to blocking using skim milk powder to minimize non-specific binding. The membrane was then incubated overnight at 4 °C with mouse monoclonal anti-GST (1:10,000) and anti-His (1:10,000) antibodies (Sanying Bio, Wuhan, China). The membrane was washed three times with TBST to remove any unbound antibodies. Following that, the membrane was exposed to a secondary antibody conjugated with horseradish peroxidase (Sanying Bio, Wuhan, China, 1:30,000) at room temperature for 1 h. The bands corresponding to the target proteins were visualized using the ECL system.
4.9. Enzyme Biometry
Enzyme activity tests are based on our previous research [
35]. The measurement of ROS levels was performed according to the instructions provided with the ROS detection kit (Jiancheng, Nanjing, China). In brief, cells were collected and suspended in PBS. After incubation with the 7-Dichlorofluorescin Diacetate probe (10 μmol/L) at 37 °C for 1 h, the cells were collected and resuspended in PBS. Subsequently, the fluorescence intensity was determined using a fluorescence spectrophotometer. The protein content in the cell samples was measured using the BCA protein concentration determination kit (Beyotime, Shanghai, China), and the final results were expressed as fluorescence units per milligram of protein.
The assessment of T-AOC was conducted using the Total Antioxidant Capacity Detection Kit (Beyotime, Shanghai, China) based on the 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) method. Following the collection and washing of cells with PBS for the subsequent detection step, the supernatant was transferred to a 96-well plate, and the ABTS working solution was added. Subsequently, the 96-well plate was incubated at room temperature for 6 min and measured using the Spark® Multimode Microplate Reader (V2.3). The protein content in the cell samples was determined using the BCA protein concentration determination kit (Beyotime, Shanghai, China China). The total antioxidant capacity of the samples was determined by referencing the standard curve of the Trolox standard solution provided in the kit. The final results of total antioxidant capacity are typically expressed as mmol/mg or mmol/g of protein weight.
4.10. Statistical Analysis
The mean values and standard deviation (SD) were used to describe the data. Statistical significance was set at a p-value less than 0.05 (p < 0.05). Group differences were assessed using either one-way ANOVA or two-way ANOVA. Statistical analyses were conducted using GraphPad Prism software version 6.0.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}