

A Novel Lipase from Streptomyces exfoliatus DSMZ 41693 for Biotechnological Applications
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Search, Cloning, and Expression of Novel Lipases from Streptomyces exfoliatus DSMZ 41693
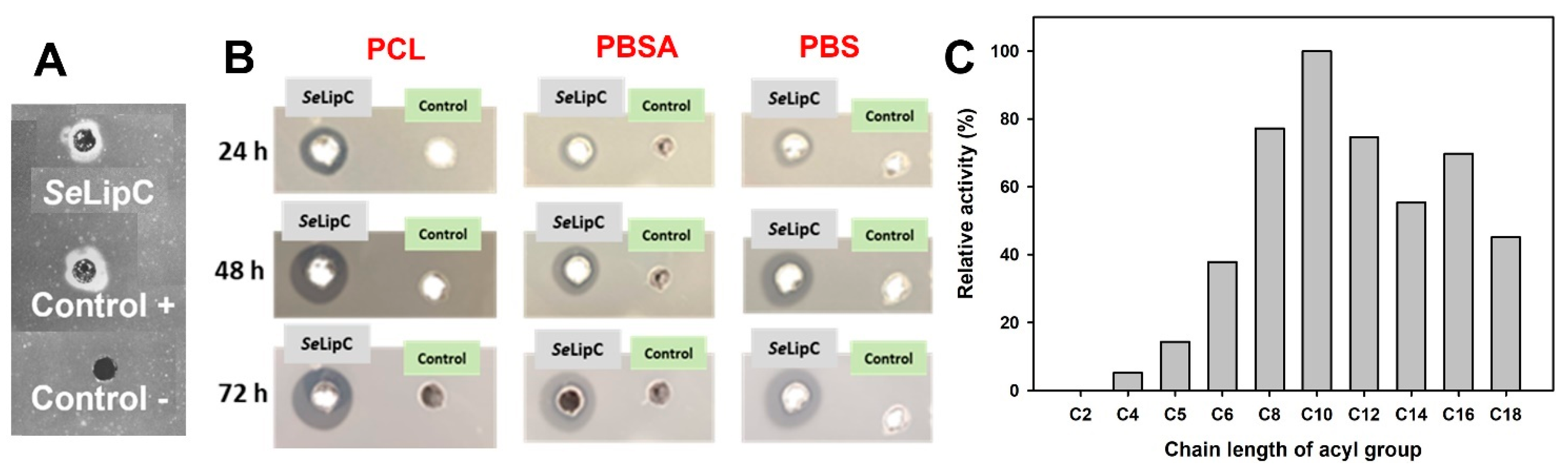
2.2. Purification and Structural Characterization of SeLipC
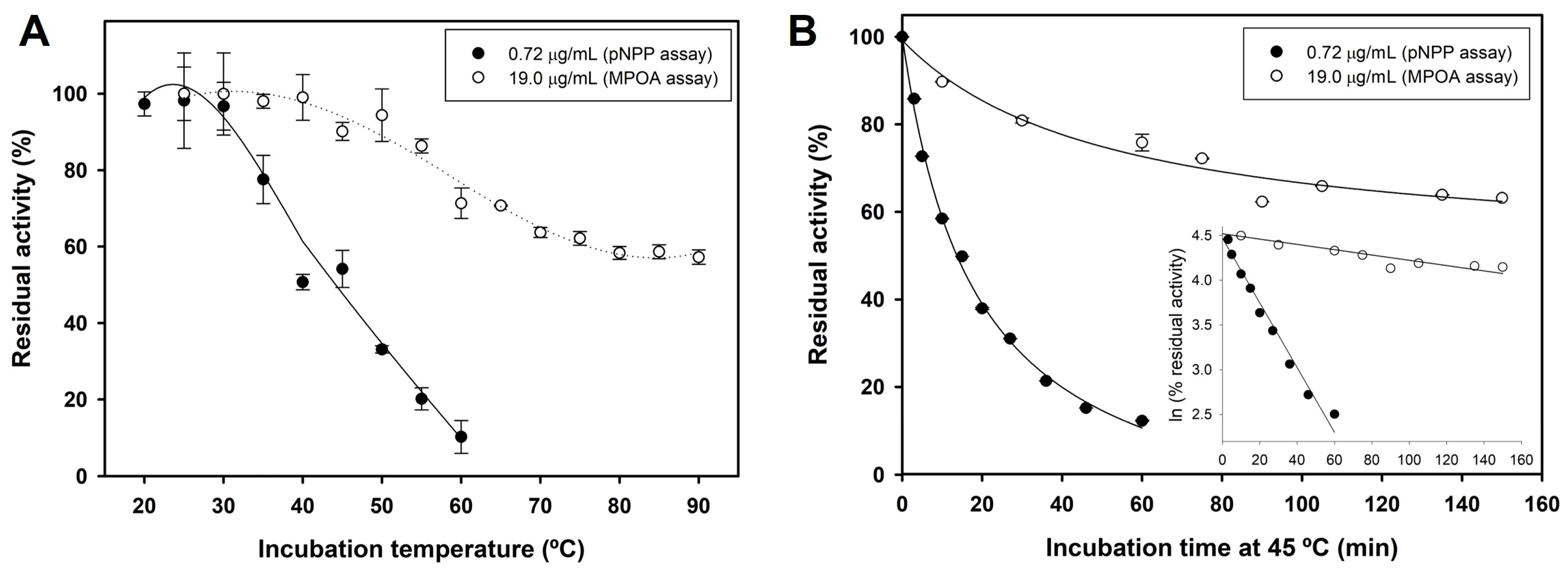
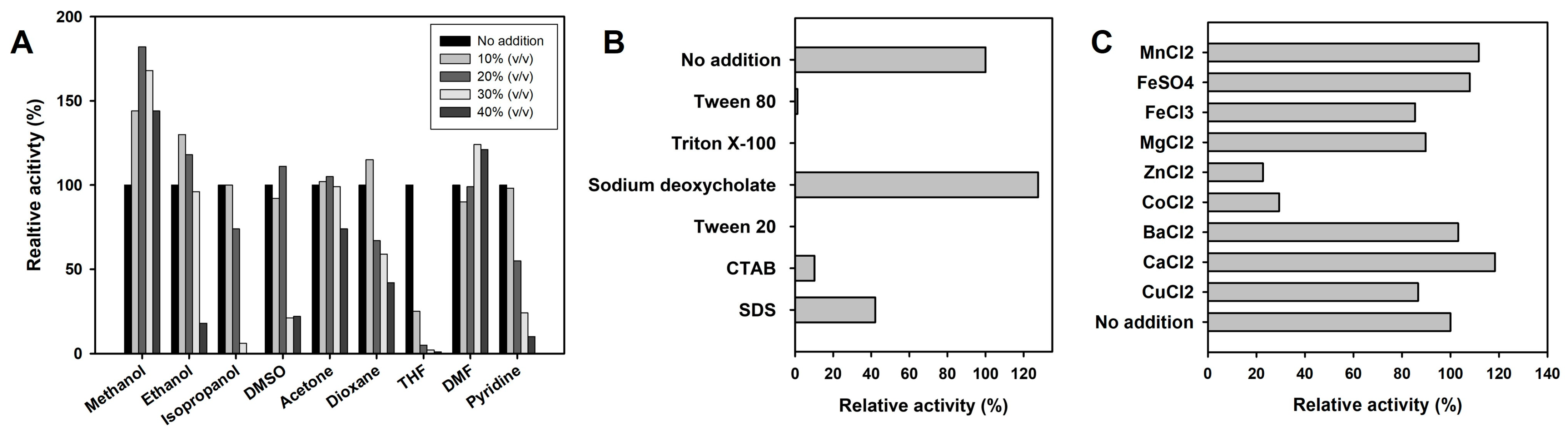
2.3. Biochemical Characterization of SeLipC
2.4. Synthesis of Glycosylated Lipids Catalyzed by SeLipC
3. Discussion


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipase Source | Maximum Activity at | Kinetic Parameters Using pNPP as a Substrate | Reference | ||||
|---|---|---|---|---|---|---|---|
| T (°C) | pH | Vmax (IU/mg) | KM (µM) | kcat (s−1) | kcat/KM (s−1 M−1) | ||
| Actinomycetes | |||||||
| Amycolatopsis mediterranei DSM 43304 | 60 | 8.0 | 2.53 × 103 | 99 | n.d. | n.d. | [46] |
| Streptomyces bambergiensis OC 25-4 | 50 | 8.0 | n.d. | n.d. | n.d. | n.d. | [48] |
| Streptomyces cellulosae AU-10 | 40 | 9.0 | n.d. | 340 | n.d. | n.d. | [19] |
| Streptomyces coelicolor A3(2) | 55 | 8.0 | n.d. | n.d. | n.d. | n.d. | [54] |
| Streptomyces fradiae var. k11 | 55 | 9.8 | 436.6 | 139 | n.d. | n.d. | [42] |
| Streptomyces rimosus R6-554W | 55 | 9.5 | n.d. | n.d. | n.d. | n.d. | [52] |
| Streptomyces sp. CS133 | 40 | 7.5 | n.d. | 152 | n.d. | n.d. | [81] |
| Streptomyces sp. CS268 | 30 | 8.0 | 206 × 103 | 2920 | n.d. | n.d. | [50] |
| Streptomyces sp. CS273 | 40 | 8.5 | n.d. | n.d. | n.d. | n.d. | [51] |
| Streptomyces sp. CS326 | 40 | 7.5 | 4.6 × 103 | 240 | n.d. | n.d. | [83] |
| Streptomyces sp. OC 119-7 | 50 | 8.0 | n.d. | n.d. | n.d. | n.d. | [49] |
| Streptomyces sp. W007 | 40 | 7.0 | n.d. | n.d. | n.d. | n.d. | [44] |
| Streptomyces violascens OC125-8 | 40 | 8.0 | 600 | 259 | n.d. | n.d. | [47] |
| Streptomyces exfoliatus DSMZ 41693 | 60 | 8.5 | 124 | 49.6 | 57 | 1.15 × 106 | This study |
| Other microorganisms | |||||||
| Bacillus thermoamylovorans BHK67 | 55 | 7.5 | n.d. | 7720 | 227 | 2.94 × 104 | [84] |
| Burkholderia multivorans V2 | 45 | 8.0 | 5.62 | 1560 | n.d. | n.d. | [85] |
| Cellulomonas flavigena | 30 | 7.0 | n.d. | 1330 | 7.22 | 5.43 × 103 | [86] |
| Pseudomonas punonensis | 30 | 9.0 | n.d. | 160 | 3.48 | 1.79 × 104 | [87] |
| Rhizopus chinensis | 40 | 8.5 | n.d. | 304 | 18.9 | 6.22 × 104 | [88] |
| Yarrowia lipolytica | 40 | 8.0 | n.d. | 108 | 158 | 1.53 × 106 | [89] |
4. Materials and Methods
4.1. Materials
4.2. Bacterial Strains, Culture Media, and Culture Conditions
4.3. Cloning and Heterologous Expression of Lip Genes from S. exfoliatus by Rhodococcus sp. T104
4.4. Purification of Recombinant SeLipC from Streptomyces exfoliatus
4.5. Protein Sequence and Structural Analysis of SeLipC
4.6. Standard Activity Assay Using p-Nitrophenyl Palmitate as Substrate (pNPP Assay)
4.7. Esterase Activity Assay Using Methyl Phenoxyacetate as Substrate (MPOA Assay)
4.8. Rhodamine Plate Assay for Lipase Activity Detection
4.9. Detection of Hydrolytic Activity on Polyester-Type Plastics
4.10. Biochemical Characterization of SeLipC
4.11. Enzymatic Synthesis of Sugar Fatty Acid Esters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casas-Godoy, L.; Gasteazoro, F.; Duquesne, S.; Bordes, F.; Marty, A.; Sandoval, G. Lipases: An overview. In Lipases and Phospholipases: Methods in Molecular Biology, 2nd ed.; Sandoval, G., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1835, pp. 3–38. [Google Scholar] [CrossRef]
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial applications of enzymes: Recent advances, techniques, and outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef]
- Akram, F.; Mir, A.S.; ul Haq, I.; Roohi, A. An appraisal on prominent industrial and biotechnological applications of bacterial lipases. Mol. Biotechnol. 2023, 65, 521–543. [Google Scholar] [CrossRef]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial lipases and their industrial applications: A comprehensive review. Microb. Cell Factories 2020, 19, 169. [Google Scholar] [CrossRef]
- Fatima, S.; Faryad, A.; Ataa, A.; Joyia, F.A.; Parvaiz, A. Microbial lipase production: A deep insight into the recent advances of lipase production and purification techniques. Biotechnol. Appl. Biochem. 2021, 68, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Hoyos, L.V.; Ramirez, L.; Yarce, C.J.; Alvarez-Vasco, C.; Ortega, N.H.C. Sustainable production of glycolipids by biocatalyst on renewable deep eutectic solvents. Catalysts 2021, 11, 13. [Google Scholar] [CrossRef]
- Perez, B.; Anankanbil, S.; Guo, Z. Synthesis of sugar fatty acid esters and their industrial utilizations. In Fatty Acids: Chemistry, Synthesis, and Applications; Ahmad, M.U., Ed.; AOCS Press: Champaign, IL, USA, 2017; pp. 329–354. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, H.Y.; Hao, T.Y.; Li, S.R. In vitro antibacterial activities and mechanism of sugar fatty acid esters against five food-related bacteria. Food Chem. 2015, 187, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Snoch, W.; Wnuk, D.; Witko, T.; Staron, J.; Bojarski, A.J.; Jarek, E.; Plou, F.J.; Guzik, M. In Search of effective anticancer agents-novel sugar esters based on polyhydroxyalkanoate monomers. Int. J. Mol. Sci. 2021, 22, 19. [Google Scholar] [CrossRef]
- Kobayashi, T. Lipase-catalyzed syntheses of sugar esters in non-aqueous media. Biotechnol. Lett. 2011, 33, 1911–1919. [Google Scholar] [CrossRef]
- Chakravorty, D.; Parameswaran, S.; Dubey, V.K.; Patra, S. Unraveling the rationale behind organic solvent stability of lipases. Appl. Biochem. Biotechnol. 2012, 167, 439–461. [Google Scholar] [CrossRef]
- Garcia-Oliva, C.; Perona, A.; Rumbero, A.; Hoyos, P.; Hernaiz, M.J. Enzymatic synthesis and molecular modelling studies of rhamnose esters using lipase from Pseudomonas stutzeri. Int. J. Mol. Sci. 2022, 23, 2239. [Google Scholar] [CrossRef]
- Del Carratore, F.; Hanko, E.K.R.; Breitling, R.; Takano, E. Biotechnological application of Streptomyces for the production of clinical drugs and other bioactive molecules. Curr. Opin. Biotechnol. 2022, 77, 102762. [Google Scholar] [CrossRef]
- Spasic, J.; Mandic, M.; Djokic, L.; Nikodinovic-Runic, J. Streptomyces spp. in the biocatalysis toolbox. Appl. Microbiol. Biotechnol. 2018, 102, 3513–3536. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.X.; Chen, S.Y.; Xu, L.; Cai, J.; Wang, J.; Wang, Y.H. Structural basis for the regiospecificity of a lipase from Streptomyces sp. W007. Int. J. Mol. Sci. 2022, 23, 5822. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Li, D.M.; Qu, M.; Durrani, R.; Yang, B.; Wang, Y.H. Immobilized MAS1 lipase showed high esterification activity in the production of triacylglycerols with n-3 polyunsaturated fatty acids. Food Chem. 2017, 216, 260–267. [Google Scholar] [CrossRef] [PubMed]
- H-Kittikun, A.; Prasertsan, P.; Zimmermann, W.; Seesuriyachan, P.; Chaiyaso, T. Sugar ester synthesis by thermostable lipase from Streptomyces thermocarboxydus ME168. Appl. Biochem. Biotechnol. 2012, 166, 1969–1982. [Google Scholar] [CrossRef] [PubMed]
- Sarac, N.; Ugur, A.; Sen, B. A green alternative for biodiesel production: Transesterification with Streptomyces sp. AU-1 lipase. Rom. Biotechnol. Lett. 2017, 22, 13060–13067. [Google Scholar]
- Boran, R. Detergent compatible extracellular lipase from Streptomyces cellulosae AU-10: A green alternative for the detergent industry. J. Surfactants Deterg. 2018, 21, 565–573. [Google Scholar] [CrossRef]
- Barriuso, J.; Martínez, M.J. In silico metagenomes mining to discover novel esterases with industrial application by sequential search strategies. J. Microbiol. Biotechnol. 2015, 25, 732–737. [Google Scholar] [CrossRef]
- Barriuso, J.; Prieto, A.; Martínez, M.J. Fungal genomes mining to discover novel sterol esterases and lipases as catalysts. BMC Genom. 2013, 14, 712. [Google Scholar] [CrossRef]
- Schallmey, M.; Koopmeiners, J.; Wells, E.; Wardenga, R.; Schallmey, A. Expanding the halohydrin dehalogenase enzyme family: Identification of novel enzymes by database mining. Appl. Environ. Microbiol. 2014, 80, 7303–7315. [Google Scholar] [CrossRef]
- van Loo, B.; Kingma, J.; Arand, M.; Wubbolts, M.G.; Janssen, D.B. Diversity and biocatalytic potential of epoxide hydrolases identified by genome analysis. Appl. Environ. Microbiol. 2006, 72, 2905–2917. [Google Scholar] [CrossRef]
- Schmitz, L.M.; Schäper, J.; Rosenthal, K.; Lütz, S. Accessing the biocatalytic potential for c-h-activation by targeted genome mining and screening. ChemCatChem 2019, 11, 5766–5777. [Google Scholar] [CrossRef]
- Wiedner, R.; Gruber-Khadjawi, M.; Schwab, H.; Steiner, K. Discovery of a novel (R)-selective bacterial hydroxynitrile lyase from Acidobacterium capsulatum. Comput. Struct. Biotechnol. J. 2014, 10, 58–62. [Google Scholar] [CrossRef]
- Martinez, V.; Hormigo, D.; Del Cerro, C.; Gomez de Santos, P.; Garcia-Hidalgo, J.; Arroyo, M.; Prieto, A.; Garcia, J.L.; de la Mata, I. Genome sequence of Streptomyces exfoliatus DSMZ 41693, a source of poly(3-hydroxyalkanoate)-degrading enzymes. Genome Announc. 2014, 2, e01272-13. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Akoh, C.C.; Lee, G.C.; Liaw, Y.C.; Huang, T.H.; Shaw, J.F. GDSL family of serine esterases/lipases. Prog. Lipid Res. 2004, 43, 534–552. [Google Scholar] [CrossRef]
- Holmquist, M. Alpha/beta-hydrolase fold enzymes: Structures, functions and mechanisms. Curr. Protein Pept. Sci. 2000, 1, 209–235. [Google Scholar] [CrossRef]
- Stobiecka, A.; Wysocki, S.; Brzozowski, A.M. Fluorescence study of fungal lipase from Humicola lanuginosa. J. Photochem. Photobiol. B Biol. 1998, 45, 95–102. [Google Scholar] [CrossRef]
- Stobiecka, A. Acrylamide-quenching of Rhizomucor miehei lipase. J. Photochem. Photobiol. B Biol. 2005, 80, 9–18. [Google Scholar] [CrossRef]
- Kouker, G.; Jaeger, K.E. Specific and sensitive plate assay for bacterial lipases. Appl. Environ. Microbiol. 1987, 53, 211–213. [Google Scholar] [CrossRef]
- García-Hidalgo, J.; Hormigo, D.; Prieto, M.A.; Arroyo, M.; de la Mata, I. Extracellular production of Streptomyces exfoliatus poly(3-hydroxybutyrate) depolymerase in Rhodococcus sp. T104: Determination of optimal biocatalyst conditions. Appl. Microbiol. Biotechnol. 2012, 93, 1975–1988. [Google Scholar] [CrossRef]
- Martinez, V.; Gomez de Santos, P.; Garcia-Hidalgo, J.; Hormigo, D.; Auxiliadora Prieto, M.; Arroyo, M.; de la Mata, I. Novel extracellular medium-chain-length polyhydroxyalkanoate depolymerase from Streptomyces exfoliatus K10 DSMZ 41693: A promising biocatalyst for the efficient degradation of natural and functionalized mcl-PHAs. Appl. Microbiol. Biotechnol. 2015, 99, 9605–9615. [Google Scholar] [CrossRef]
- Garcia-Hidalgo, J.; Hormigo, D.; Arroyo, M.; de la Mata, I. Novel extracellular PHB depolymerase from Streptomyces ascomycinicus: PHB copolymers degradation in acidic conditions. PLoS ONE 2013, 8, e71699. [Google Scholar] [CrossRef]
- Serrano-Aguirre, L.; Velasco-Bucheli, R.; Garcia-Alvarez, B.; Saborido, A.; Arroyo, M.; de la Mata, I. Novel bifunctional acylase from Actinoplanes utahensis: A versatile enzyme to synthesize antimicrobial compounds and use in quorum quenching processes. Antibiotics 2021, 10, 922. [Google Scholar] [CrossRef]
- Saxena, R.K.; Sheoran, A.; Giri, B.; Davidson, W.S. Purification strategies for microbial lipases. J. Microbiol. Methods 2003, 52, 1–18. [Google Scholar] [CrossRef]
- Arpigny, J.L.; Jaeger, K.E. Bacterial lipolytic enzymes: Classification and properties. Biochem. J. 1999, 343, 177–183. [Google Scholar] [CrossRef]
- Kovacic, F.; Babic, N.; Krauss, U.; Jaeger, K.E. Classification of lipolytic enzymes from bacteria. In Aerobic Utilization of Hydrocarbons, Oils, and Lipids: Handbook of Hydrocarbon and Lipid Microbiology; Rojo, F., Ed.; Springer: Cham, Switzerland, 2019; pp. 255–289. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Meng, K.; Wang, Y.R.; Luo, H.Y.; Yang, P.L.; Shi, P.J.; Wu, N.F.; Fan, Y.; Li, J.; Yao, B. A novel proteolysis-resistant lipase from keratinolytic Streptomyces fradiae var. k11. Enzyme Microb. Technol. 2008, 42, 346–352. [Google Scholar] [CrossRef]
- Sommer, P.; Bormann, C.; Gotz, F. Genetic and biochemical characterization of a new extracellular lipase from Streptomyces cinnamomeus. Appl. Environ. Microbiol. 1997, 63, 3553–3560. [Google Scholar] [CrossRef]
- Yuan, D.J.; Lan, D.M.; Xin, R.P.; Yang, B.; Wang, Y.H. Screening and characterization of a thermostable lipase from marine Streptomyces sp. strain W007. Biotechnol. Appl. Biochem. 2016, 63, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.X.; Hou, S.L.; Lan, D.M.; Wang, X.M.; Liu, J.S.; Khan, F.I.; Wang, Y.H. Crystal structure of a lipase from Streptomyces sp. strain W007-implications for thermostability and regiospecificity. FEBS J. 2017, 284, 3506–3519. [Google Scholar] [CrossRef] [PubMed]
- Dheeman, D.S.; Henehan, G.T.M.; Frias, J.M. Purification and properties of Amycolatopsis mediterranei DSM 43304 lipase and its potential in flavour ester synthesis. Bioresour. Technol. 2011, 102, 3373–3379. [Google Scholar] [CrossRef] [PubMed]
- Boran, R.; Ugur, A.; Sarac, N.; Ceylan, O. Characterisation of Streptomyces violascens OC125-8 lipase for oily wastewater treatment. 3 Biotech 2019, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Ugur, A.; Sarac, N.; Boran, R.; Ayaz, B.; Ceylan, O.; Okmen, G. New lipase for biodiesel production: Partial purification and characterization of LipSB 25-4. ISRN Biochem. 2014, 2014, 289749. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, B.; Ugur, A.; Boran, R. Purification and characterization of organic solvent-tolerant lipase from Streptomyces sp. OC119-7 for biodiesel production. Biocatal. Agric. Biotechnol. 2015, 4, 103–108. [Google Scholar] [CrossRef]
- Mander, P.; Cho, S.S.; Simkhada, J.R.; Choi, Y.H.; Park, D.J.; Ha, J.W.; Yoo, J.C. An organic solvent-tolerant alkaline lipase from Streptomyces sp. CS268 and its application in biodiesel production. Biotechnol. Bioprocess Eng. 2012, 17, 67–75. [Google Scholar] [CrossRef]
- Mander, P.; Yoo, H.Y.; Kim, S.W.; Choi, Y.H.; Cho, S.S.; Yoo, J.C. Transesterification of waste cooking oil by an organic solvent-tolerant alkaline lipase from Streptomyces sp. CS273. Appl. Biochem. Biotechnol. 2014, 172, 1377–1389. [Google Scholar] [CrossRef]
- Abramic, M.; Lescic, I.; Korica, T.; Vitale, L.; Saenger, W.; Pigac, J. Purification and properties of extracellular lipase from Streptomyces rimosus. Enzyme Microb. Technol. 1999, 25, 522–529. [Google Scholar] [CrossRef]
- Cote, A.; Shareck, F. Cloning, purification and characterization of two lipases from Streptomyces coelicolor A3(2). Enzyme Microb. Technol. 2008, 42, 381–388. [Google Scholar] [CrossRef]
- Bielen, A.; Cetkovic, H.; Long, P.F.; Schwab, H.; Abramic, M.; Vujaklija, D. The SGNH-hydrolase of Streptomyces coelicolor has (aryl)esterase and a true lipase activity. Biochimie 2009, 91, 390–400. [Google Scholar] [CrossRef]
- Perez, C.; Juarez, K.; Garciacastells, E.; Soberon, G.; Servingonzalez, L. Cloning, characterization, and expression in Streptomyces lividans 66 of an extracellular lipase-encoding gene from Streptomyces sp. M11. Gene 1993, 123, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.Y.; Swenson, L.; Castro, C.; Derewenda, U.; Minor, W.; Arai, H.; Aoki, J.; Inoue, K.; Servin-Gonzalez, L.; Derewenda, Z.S. Structure of a microbial homologue of mammalian platelet-activating factor acetylhydrolases: Streptomyces exfoliatus lipase at 1.9 angstrom resolution. Structure 1998, 6, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Vujaklija, D.; Schroder, W.; Abramic, M.; Zou, P.J.; Lescic, I.; Franke, P.; Pigac, J. A novel streptomycete lipase: Cloning, sequencing and high-level expression of the Streptomyces rimosus GDS(L)-lipase gene. Arch. Microbiol. 2002, 178, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Cruz, H.; Perez, C.; Wellington, E.; Castro, C.; Servin-Gonzalez, L. Sequence of the Streptomyces albus G lipase-encoding gene reveals the presence of a prokaryotic lipase family. Gene 1994, 144, 141–142. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.P.; Wang, X.F.; Jiang, H.; Sun, J.A.; Mao, X.Z. Identification of a GDSL lipase from Streptomyces bacillaris and its application in the preparation of free astaxanthin. J. Biotechnol. 2021, 325, 207–214. [Google Scholar] [CrossRef]
- Berger, R.; Hoffmann, M.; Keller, U. Molecular analysis of a gene encoding a cell-bound esterase from Streptomyces chrysomallus. J. Bacteriol. 1998, 180, 6396–6399. [Google Scholar] [CrossRef] [PubMed]
- Soror, S.H.; Verma, V.; Rao, R.; Rasool, S.; Koul, S.; Qazi, G.N.; Cullum, J. A cold-active esterase of Streptomyces coelicolor A3(2): From genome sequence to enzyme activity. J. Ind. Microbiol. Biotechnol. 2007, 34, 525–531. [Google Scholar] [CrossRef]
- Brault, G.; Shareck, F.; Hurtubise, Y.; Lepine, F.; Doucet, N. Isolation and characterization of EstC, a new cold-active esterase from Streptomyces coelicolor A3(2). PLoS ONE 2012, 7, e32041. [Google Scholar] [CrossRef]
- Wang, B.J.; Wang, A.; Cao, Z.Y.; Zhu, G.P. Characterization of a novel highly thermostable esterase from the Gram-positive soil bacterium Streptomyces lividans TK64. Biotechnol. Appl. Biochem. 2016, 63, 334–343. [Google Scholar] [CrossRef]
- Hale, V.; McGrew, M.; Carlson, B.; Schottel, J.L. Heterologous expression and secretion of a Streptomyces scabies esterase in Streptomyces lividans and Escherichia coli. J. Bacteriol. 1992, 174, 2431–2439. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11 molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Brzozowski, A.M.; Savage, H.; Verma, C.S.; Turkenburg, J.P.; Lawson, D.M.; Svendsen, A.; Patkar, S. Structural origins of the interfacial activation in Thermomyces (Humicola) lanuginosa lipase. Biochemistry 2000, 39, 15071–15082. [Google Scholar] [CrossRef] [PubMed]
- Palomo, J.M.; Fuentes, M.; Fernandez-Lorente, G.; Mateo, C.; Guisan, J.M.; Fernandez-Lafuente, R. General trend of lipase to self-assemble giving bimolecular aggregates greatly modifies the enzyme functionality. Biomacromolecules 2003, 4, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.; Palomo, J.M.; Fernandez-Lorente, G.; Illanes, A.; Guisan, J.M.; Fernandez-Lafuente, R. Effect of lipase-lipase interactions in the activity, stability and specificity of a lipase from Alcaligenes sp. Enzyme Microb. Technol. 2006, 39, 259–264. [Google Scholar] [CrossRef]
- Fernandez-Lorente, G.; Palomo, J.M.; Fuentes, M.; Mateo, C.; Guisan, J.M.; Fernandez-Lafuente, R. Self-assembly of Pseudomonas fluorescens lipase into bimolecular aggregates dramatically affects functional properties. Biotechnol. Bioeng. 2003, 82, 232–237. [Google Scholar] [CrossRef]
- Timucin, E.; Sezerman, O.U. The conserved lid tryptophan, W211, potentiates thermostability and thermoactivity in bacterial thermoalkalophilic lipases. PLoS ONE 2013, 8, e85186. [Google Scholar] [CrossRef]
- Zhu, K.; Jutila, A.; Tuominen, E.K.J.; Kinnunen, P.K.J. Effects of i-propanol on the structural dynamics of Thermomyces lanuginosa lipase revealed by tryptophan fluorescence. Protein Sci. 2001, 10, 339–351. [Google Scholar] [CrossRef]
- Priyanka, P.; Tan, Y.Q.; Kinsella, G.K.; Henehan, G.T.; Ryan, B.J. Solvent stable microbial lipases: Current understanding and biotechnological applications. Biotechnol. Lett. 2019, 41, 203–220. [Google Scholar] [CrossRef]
- Lescic, I.; Vukelic, B.; Majeric-Elenkov, M.; Saenger, W.; Abramic, M. Substrate specificity and effects of water-miscible solvents on the activity and stability of extracellular lipase from Streptomyces rimosus. Enzyme Microb. Technol. 2001, 29, 548–553. [Google Scholar] [CrossRef]
- Khmelnitsky, Y.L.; Mozhaev, V.V.; Belova, A.B.; Sergeeva, M.V.; Martinek, K. Denaturation capacity: A new quantitative criterion for selection of organic solvents as reaction media in biocatalysis. Eur. J. Biochem. 1991, 198, 31–41. [Google Scholar] [CrossRef]
- Arroyo, M.; Torres-Guzman, R.; de la Mata, I.; Castillon, M.; Acebal, C. Prediction of penicillin V acylase stability in water-organic co-solvent monophasic systems as a function of solvent composition. Enzyme Microb. Technol. 2000, 27, 122–126. [Google Scholar] [CrossRef]
- Mateos-Diaz, E.; Amara, S.; Roussel, A.; Longhi, S.; Cambillau, C.; Carriere, F. Probing conformational changes and interfacial recognition site of lipases with surfactants and inhibitors. Methods Enzymol. 2017, 583, 279–307. [Google Scholar] [CrossRef] [PubMed]
- Delorme, V.; Dhouib, R.; Canaan, S.; Fotiadu, F.; Carriere, F.; Cavalier, J.F. Effects of surfactants on lipase structure, activity, and inhibition. Pharm. Res. 2011, 28, 1831–1842. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D. Lipase catalysis in mixed micelles. ChemBioEng Rev. 2022, 9, 409–418. [Google Scholar] [CrossRef]
- Vo, C.V.T.; Luu, N.V.H.; Nguyen, T.T.H.; Nguyen, T.T.; Ho, B.Q.; Nguyen, T.H.; Tran, T.D.; Nguyen, Q.T. Screening for pancreatic lipase inhibitors: Evaluating assay conditions using p-nitrophenyl palmitate as substrate. All Life 2022, 15, 13–22. [Google Scholar] [CrossRef]
- Masomian, M.; Abd Rahman, R.; Salleh, A.; Basri, M. Analysis of comparative sequence and genomic data to verify phylogenetic relationship and explore a new subfamily of bacterial lipases. PLoS ONE 2016, 11, e0149851. [Google Scholar] [CrossRef] [PubMed]
- Mander, P.; Cho, S.S.; Simkhada, J.R.; Choi, Y.H.; Park, D.J.; Yoo, J.C. An organic solvent-tolerant lipase from Streptomyces sp. CS133 for enzymatic transesterification of vegetable oils in organic media. Process Biochem. 2012, 47, 635–642. [Google Scholar] [CrossRef]
- Chakraborty, K.; Paulraj, R. Purification and biochemical characterization of an extracellular lipase from Pseudomonas fluorescens MTCC 2421. J. Agric. Food Chem. 2009, 57, 3859–3866. [Google Scholar] [CrossRef]
- Cho, S.S.; Park, D.J.; Simkhada, J.R.; Hong, J.H.; Sohng, J.K.; Lee, O.H.; Yoo, J.C. A neutral lipase applicable in biodiesel production from a newly isolated Streptomyces sp. CS326. Bioprocess Biosyst. Eng. 2012, 35, 227–234. [Google Scholar] [CrossRef]
- Sharma, A.; Meena, K.R.; Kanwar, S.S. Molecular characterization and bioinformatics studies of a lipase from Bacillus thermoamylovorans BHK67. Int. J. Biol. Macromol. 2018, 107, 2131–2140. [Google Scholar] [CrossRef] [PubMed]
- Dandavate, V.; Jinjala, J.; Keharia, H.; Madamwar, D. Production, partial purification and characterization of organic solvent tolerant lipase from Burkholderia multivorans V2 and its application for ester synthesis. Bioresour. Technol. 2009, 100, 3374–3381. [Google Scholar] [CrossRef]
- Prajapati, V.; Patel, H.; Trivedi, U.; Patel, K. Kinetic and thermodynamic characterization of lipase produced by Cellulomonas flavigena UNP3. J. Basic Microbiol. 2014, 54, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Sonkar, K.; Singh, D.P. Biochemical characterization and thermodynamic study of lipase from psychrotolerant Pseudomonas punonensis. Biocatal. Agric. Biotechnol. 2020, 28, 101686. [Google Scholar] [CrossRef]
- Yu, X.W.; Tan, N.J.; Xiao, R.; Xu, Y. Engineering a disulfide bond in the lid hinge region of Rhizopus chinensis lipase: Increased thermostability and altered acyl chain length specificity. PLoS ONE 2012, 7, e46388. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Liu, Z.M.; Xu, L.; Zhang, H.J.; Yan, Y.J. Probing role of key residues in the divergent evolution of Yarrowia lipolytica lipase 2 and Aspergillus niger eruloyl esterase A. Microbiol. Res. 2015, 178, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Urbanek, A.K.; Mironczuk, A.M.; Garcia-Martin, A.; Saborido, A.; de la Mata, I.; Arroyo, M. Biochemical properties and biotechnological applications of microbial enzymes involved in the degradation of polyester-type plastics. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 13. [Google Scholar] [CrossRef]
- Hoshino, A.; Isono, Y. Degradation of aliphatic polyester films by commercially available lipases with special reference to rapid and complete degradation of poly(L-lactide) film by lipase PL derived from Alcaligenes sp. Biodegradation 2002, 13, 141–147. [Google Scholar] [CrossRef]
- Rosato, A.; Romano, A.; Totaro, G.; Celli, A.; Fava, F.; Zanaroli, G.; Sisti, L. Enzymatic degradation of the most common aliphatic bio-polyesters and evaluation of the mechanisms involved: An extended study. Polymers 2022, 14, 1850. [Google Scholar] [CrossRef]
- Khan, I.; Dutta, J.R.; Ganesan, R. Lactobacillus sps. lipase mediated poly (ε-caprolactone) degradation. Int. J. Biol. Macromol. 2017, 95, 126–131. [Google Scholar] [CrossRef]
- Thirunavukarasu, K.; Purushothaman, S.; Sridevi, J.; Aarthy, M.; Gowthaman, M.K.; Nakajima-Kambe, T.; Kamini, N.R. Degradation of poly(butylene succinate) and poly(butylene succinate-co-butylene adipate) by a lipase from yeast Cryptococcus sp. grown on agro-industrial residues. Int. Biodeterior. Biodegrad. 2016, 110, 99–107. [Google Scholar] [CrossRef]
- Li, F.; Hu, X.; Guo, Z.Q.; Wang, Z.Y.; Wang, Y.; Liu, D.B.; Xia, H.M.; Chen, S. Purification and characterization of a novel poly(butylene succinate)-degrading enzyme from Aspergillus sp. XH0501-a. World J. Microbiol. Biotechnol. 2011, 27, 2591–2596. [Google Scholar] [CrossRef]
- Raku, T.; Tokiwa, Y. Chemoenzymatic synthesis of fucose- or rhamnose-branched polymer. Macromol. Biosci. 2003, 3, 151–156. [Google Scholar] [CrossRef]
- Pohnlein, M.; Slomka, C.; Kukharenko, O.; Gartner, T.; Wiemann, L.O.; Sieber, V.; Syldatk, C.; Hausmann, R. Enzymatic synthesis of amino sugar fatty acid esters. Eur. J. Lipid Sci. Technol. 2014, 116, 423–428. [Google Scholar] [CrossRef]
- Zhao, G.; Wang, J.R.; Tang, Q.Y.; Lan, D.M.; Wang, Y.H. Improving the catalytic activity and thermostability of MAS1 lipase by alanine substitution. Mol. Biotechnol. 2018, 60, 319–328. [Google Scholar] [CrossRef]
- Wang, X.M.; Qin, X.L.; Li, D.M.; Yang, B.; Wang, Y.H. One-step synthesis of high-yield biodiesel from waste cooking oils by a novel and highly methanol-tolerant immobilized lipase. Bioresour. Technol. 2017, 235, 18–24. [Google Scholar] [CrossRef]
- Wang, X.M.; Zhao, X.X.; Qin, X.L.; Zhao, Z.X.; Yang, B.; Wang, Y.H. Properties of immobilized MAS1-H108A lipase and its application in the efficient synthesis of n-3 PUFA-rich triacylglycerols. Bioprocess Biosyst. Eng. 2021, 44, 575–584. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Hernandez, B.S.; Koh, S.C.; Chial, M.; Focht, D.D. Terpene-utilizing isolates and their relevance to enhanced biotransformation of polychlorinated biphenyls in soil. Biodegradation 1997, 8, 153–158. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Claveage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Speicher, D.W. Methods and strategies for the sequence analysis of proteins on PVDF membranes. Methods 1994, 6, 262–273. [Google Scholar] [CrossRef]
- Miles, A.J.; Ramalli, S.G.; Wallace, B.A. DichroWeb, a website for calculating protein secondary structure from circular dichroism spectroscopic data. Protein Sci. 2022, 31, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Comparison of CONTIN, SELCON, and CDSSTR methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Urbanek, A.K.; Rymowicz, W.; Strzelecki, M.C.; Kociuba, W.; Franczak, L.; Mironczuk, A.M. Isolation and characterization of Arctic microorganisms decomposing bioplastics. AMB Express 2017, 7, 148. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Alonso, G.; Toledo-Marcos, J.; Serrano-Aguirre, L.; Rumayor, C.; Pasero, B.; Flores, A.; Saborido, A.; Hoyos, P.; Hernáiz, M.J.; de la Mata, I.; et al. A Novel Lipase from Streptomyces exfoliatus DSMZ 41693 for Biotechnological Applications. Int. J. Mol. Sci. 2023, 24, 17071. https://doi.org/10.3390/ijms242317071
Rodríguez-Alonso G, Toledo-Marcos J, Serrano-Aguirre L, Rumayor C, Pasero B, Flores A, Saborido A, Hoyos P, Hernáiz MJ, de la Mata I, et al. A Novel Lipase from Streptomyces exfoliatus DSMZ 41693 for Biotechnological Applications. International Journal of Molecular Sciences. 2023; 24(23):17071. https://doi.org/10.3390/ijms242317071
Chicago/Turabian StyleRodríguez-Alonso, Guillermo, Juan Toledo-Marcos, Lara Serrano-Aguirre, Carlos Rumayor, Beatriz Pasero, Aida Flores, Ana Saborido, Pilar Hoyos, María J. Hernáiz, Isabel de la Mata, and et al. 2023. "A Novel Lipase from Streptomyces exfoliatus DSMZ 41693 for Biotechnological Applications" International Journal of Molecular Sciences 24, no. 23: 17071. https://doi.org/10.3390/ijms242317071
APA StyleRodríguez-Alonso, G., Toledo-Marcos, J., Serrano-Aguirre, L., Rumayor, C., Pasero, B., Flores, A., Saborido, A., Hoyos, P., Hernáiz, M. J., de la Mata, I., & Arroyo, M. (2023). A Novel Lipase from Streptomyces exfoliatus DSMZ 41693 for Biotechnological Applications. International Journal of Molecular Sciences, 24(23), 17071. https://doi.org/10.3390/ijms242317071