
Association of the Telomerase Reverse Transcriptase rs10069690 Polymorphism with the Risk, Age at Onset and Prognosis of Triple Negative Breast Cancer

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. The TERT rs10069690 SNV and Breast Cancer Risk
2.2. Exploratory Analysis of rs10069690 and Breast Cancer Risk in Subpopulations
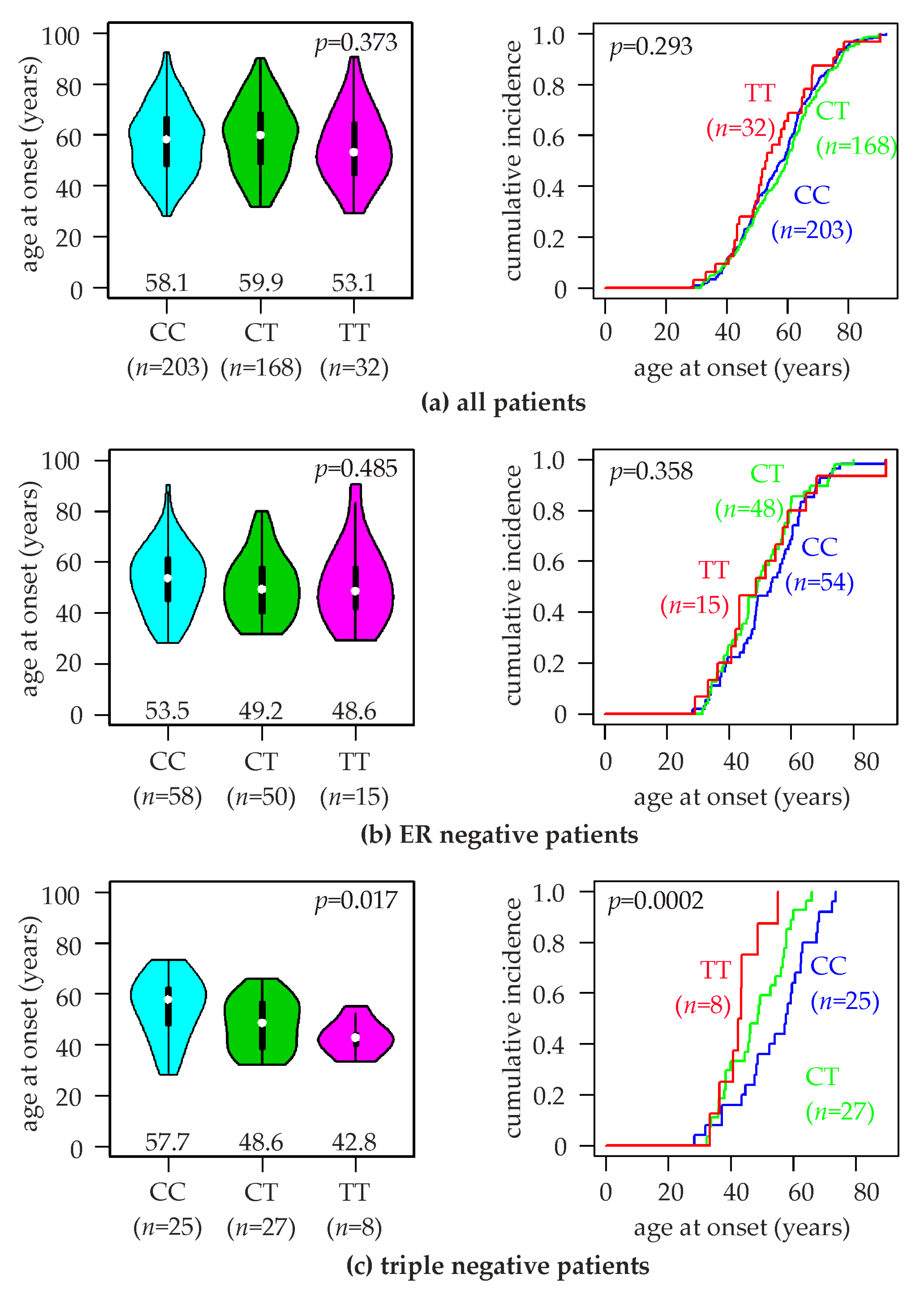
2.3. Association of the TERT rs10069690 SNV with the Age at Breast Cancer Onset
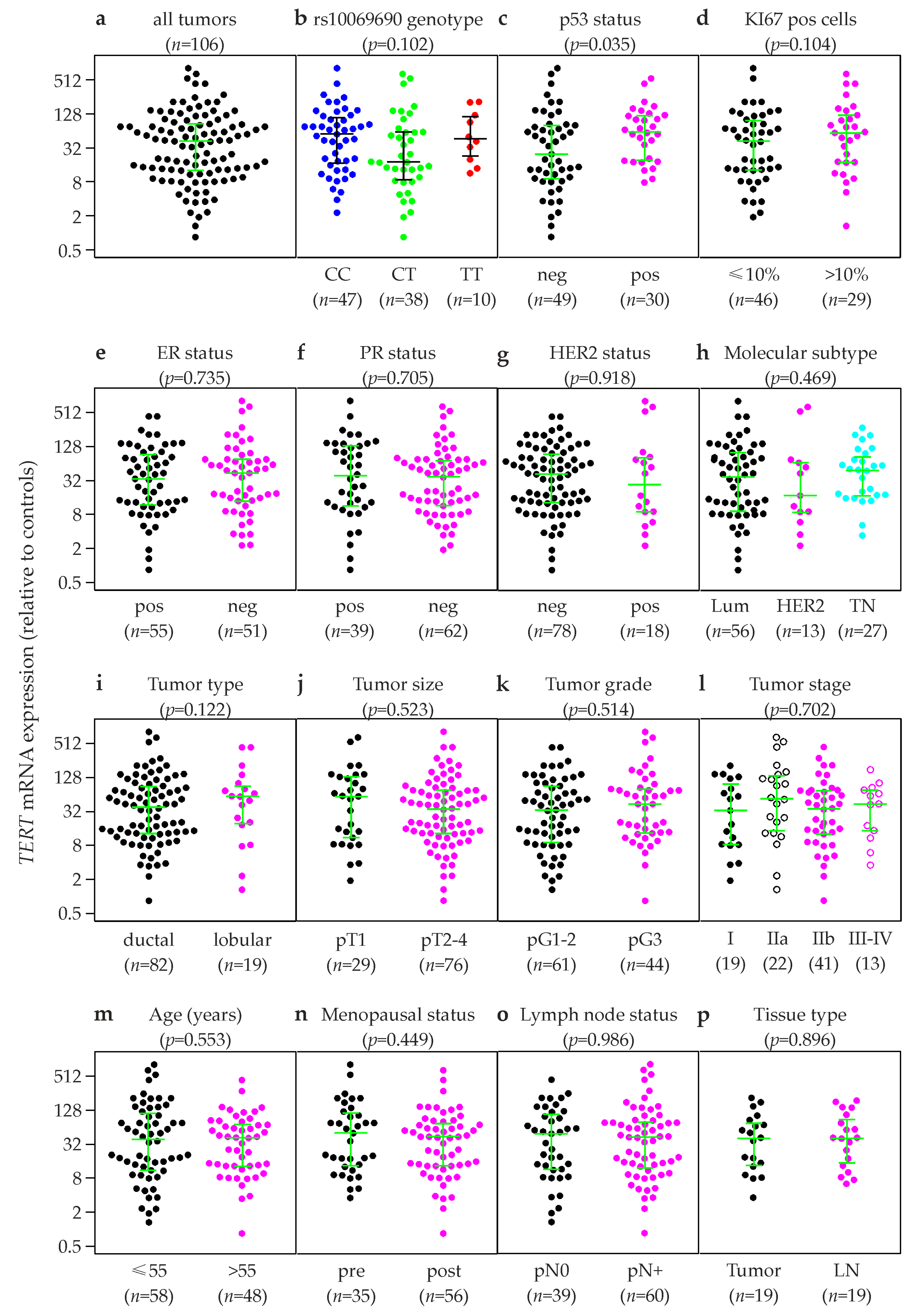
2.4. Association of TERT Expression in Primary Breast Tumors with Clinical and Histopathological Patient Characteristics
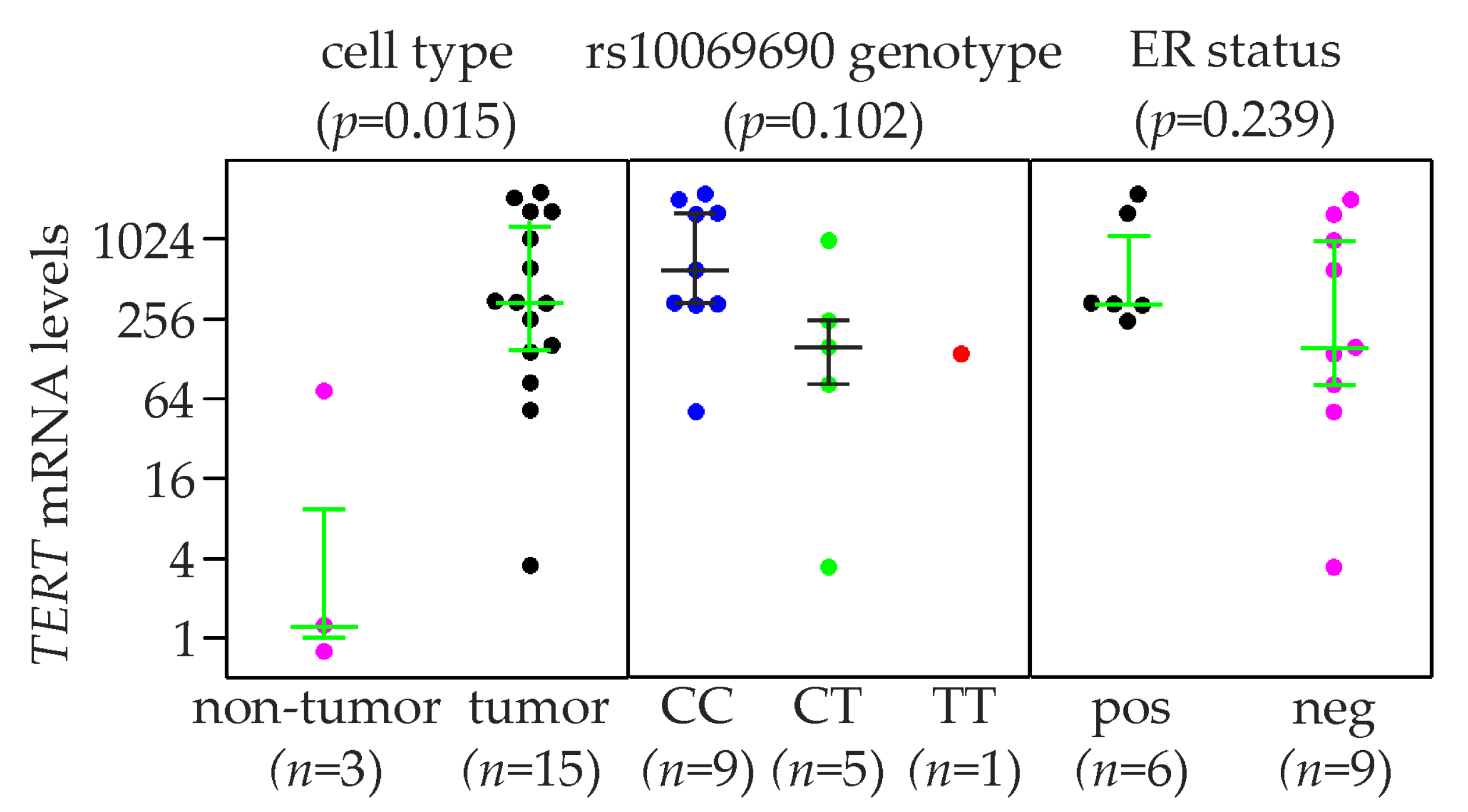
2.5. TERT Expression and rs10069690 Genotype in Breast Cancer Cell Lines
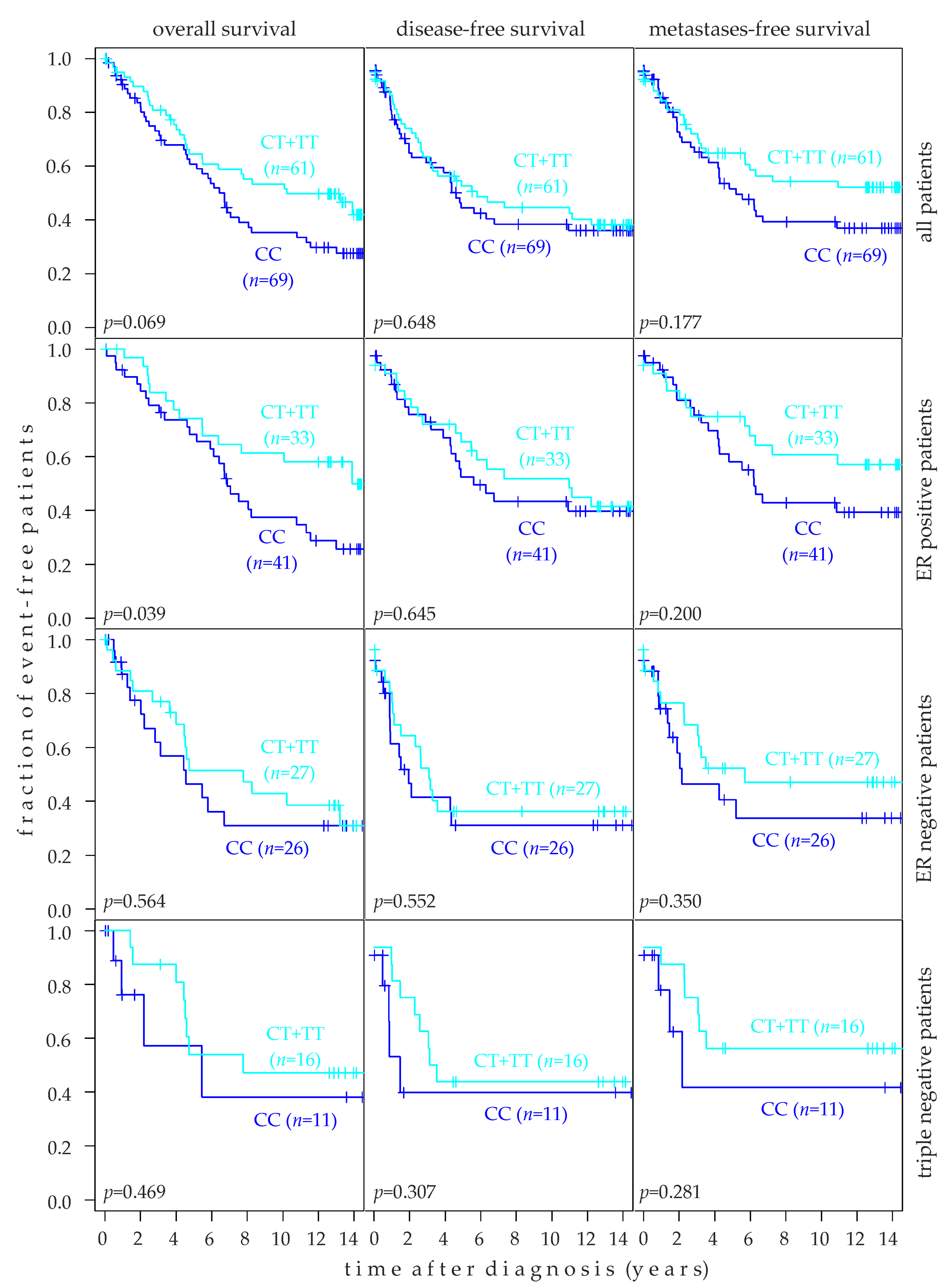
2.6. Association of rs10069690 Genotype with Breast Cancer Prognosis
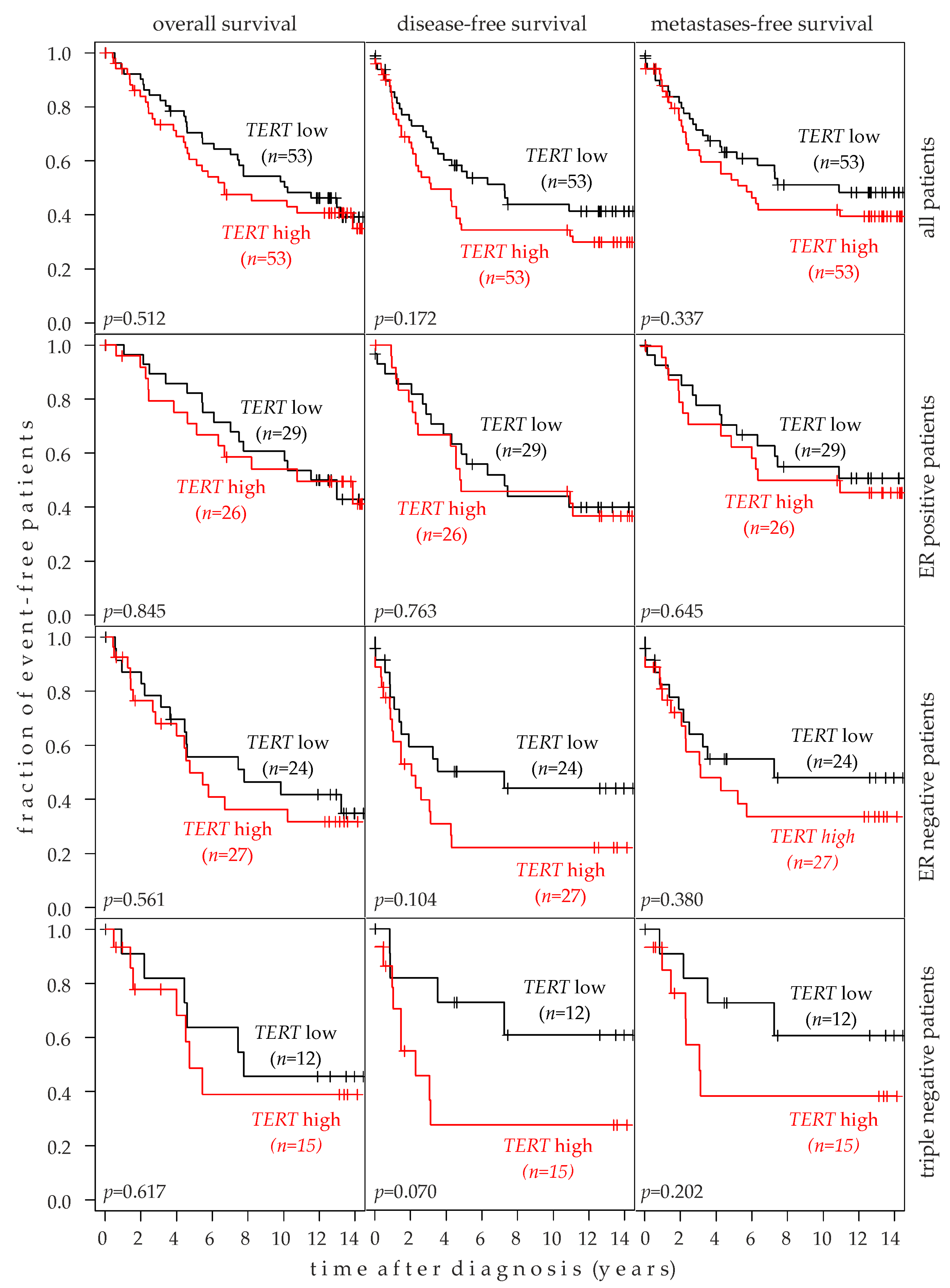
2.7. Association of TERT Expression with Breast Cancer Prognosis
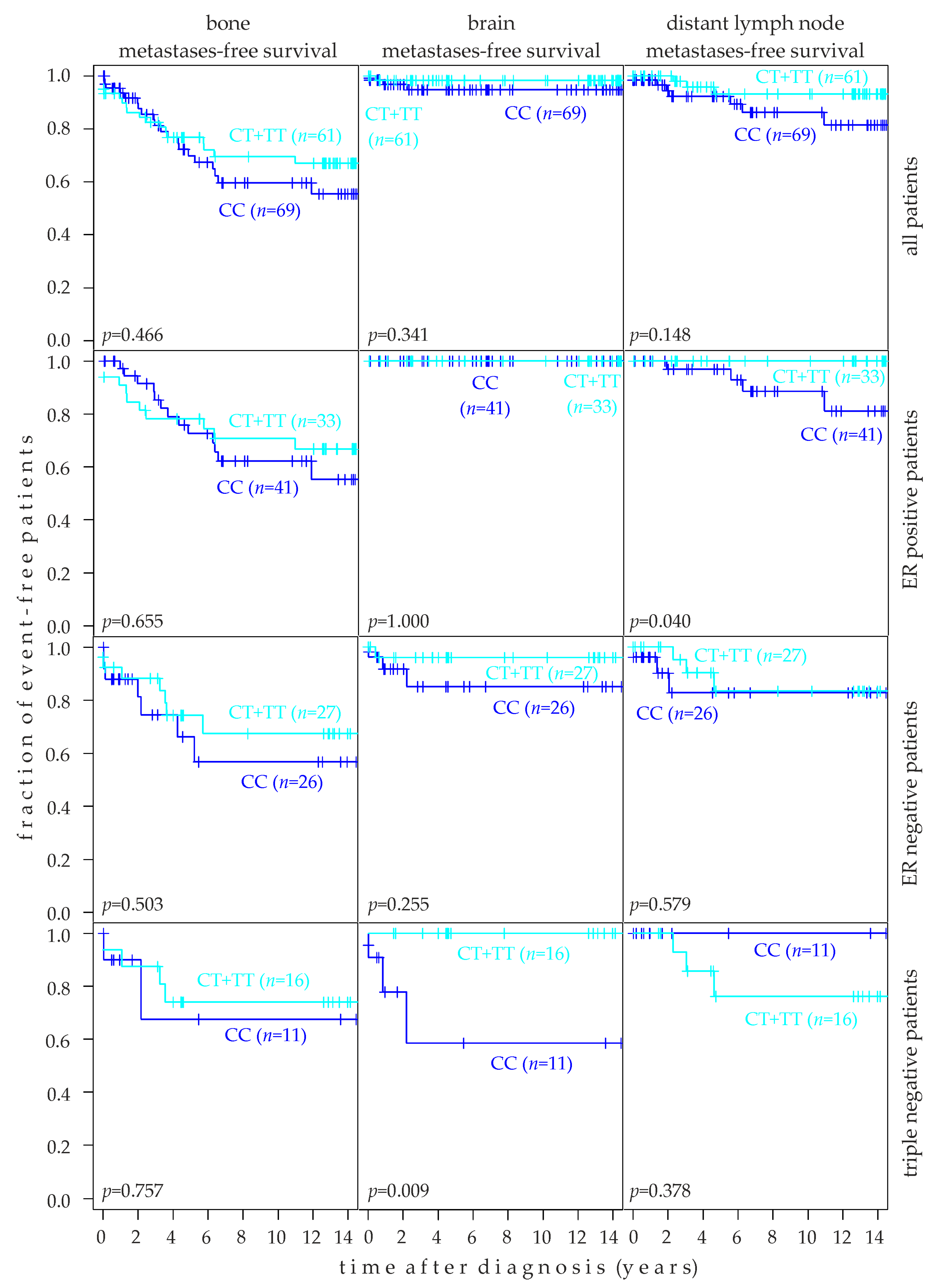
2.8. Association of rs10069690 Genotype with Target Tissue Specific Metastasis-Free Survival
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Cell Lines
4.3. DNA Isolation and SNV Genotyping
4.4. qRT-PCR Quantification of mRNA Expression Levels
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Sullivan, R.J.; Karlseder, J. Telomeres: Protecting chromosomes against genome instability. Nat. Rev. Mol. Cell Biol. 2010, 11, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lange, T. How Telomeres Solve the End-Protection Problem. Science 2009, 326, 948–952. [Google Scholar] [CrossRef] [Green Version]
- Olovnikov, A.M. A theory of marginotomy. The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon. J. Theor. Biol. 1973, 41, 181–190. [Google Scholar] [CrossRef]
- Barthel, F.P.; Wei, W.; Tang, M.; Martinez-Ledesma, E.; Hu, X.; Amin, S.B.; Akdemir, K.C.; Seth, S.; Song, X.; Wang, Q.; et al. Systematic analysis of telomere length and somatic alterations in 31 cancer types. Nat. Genet. 2017, 49, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, E.H. Telomeres and telomerase: Their mechanisms of action and the effects of altering their functions. FEBS Lett. 2005, 579, 859–862. [Google Scholar] [CrossRef] [Green Version]
- Robinson, N.J.; Schiemann, W.P. Telomerase in Cancer: Function, Regulation, and Clinical Translation. Cancers 2022, 14, 808. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.; Mitchell, J.R. Telomerase in the human organism. Oncogene 2002, 21, 564–579. [Google Scholar] [CrossRef] [Green Version]
- Greider, C.W.; Blackburn, E.H. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 1985, 43, 405–413. [Google Scholar] [CrossRef]
- Morales, C.P.; Holt, S.E.; Ouellette, M.; Kaur, K.J.; Yan, Y.; Wilson, K.S.; White, M.A.; Wright, W.E.; Shay, J.W. Absence of cancer-associated changes in human fibroblasts immortalized with telomerase. Nat. Genet. 1999, 21, 115–118. [Google Scholar] [CrossRef]
- Shay, J.W.; Bacchetti, S. A survey of telomerase activity in human cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Zheng, C.; Lindvall, C.; Hou, M.; Ekedahl, J.; Lewensohn, R.; Yan, Z.; Yang, X.; Henriksson, M.; Blennow, E.; et al. Frequent amplification of the telomerase reverse transcriptase gene in human tumors. Cancer Res. 2000, 60, 6230–6235. [Google Scholar] [PubMed]
- Blackburn, E.H.; Greider, C.W.; Szostak, J.W. Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. Nat. Med. 2006, 12, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.M.; Morin, G.B.; Chapman, K.B.; Weinrich, S.L.; Andrews, W.H.; Lingner, J.; Harley, C.B.; Cech, T.R. Telomerase catalytic subunit homologs from fission yeast and human. Science 1997, 277, 955–959. [Google Scholar] [CrossRef]
- Savage, S.A.; Stewart, B.J.; Eckert, A.; Kiley, M.; Liao, J.S.; Chanock, S.J. Genetic variation, nucleotide diversity, and linkage disequilibrium in seven telomere stability genes suggest that these genes may be under constraint. Hum. Mutat. 2005, 26, 343–350. [Google Scholar] [CrossRef]
- Baird, D.M. Variation at the TERT locus and predisposition for cancer. Expert Rev. Mol. Med. 2010, 12, e16. [Google Scholar] [CrossRef]
- Bojesen, S.E.; Pooley, K.A.; Johnatty, S.E.; Beesley, J.; Michailidou, K.; Tyrer, J.P.; Edwards, S.L.; Pickett, H.A.; Shen, H.C.; Smart, C.E.; et al. Multiple independent variants at the TERT locus are associated with telomere length and risks of breast and ovarian cancer. Nat. Genet. 2013, 45, 371–384. [Google Scholar] [CrossRef]
- Haiman, C.A.; Chen, G.K.; Vachon, C.M.; Canzian, F.; Dunning, A.; Millikan, R.C.; Wang, X.; Ademuyiwa, F.; Ahmed, S.; Ambrosone, C.B.; et al. A common variant at the TERT-CLPTM1L locus is associated with estrogen receptor-negative breast cancer. Nat. Genet. 2011, 43, 1210–1214. [Google Scholar] [CrossRef]
- Rafnar, T.; Sulem, P.; Stacey, S.N.; Geller, F.; Gudmundsson, J.; Sigurdsson, A.; Jakobsdottir, M.; Helgadottir, H.; Thorlacius, S.; Aben, K.K.; et al. Sequence variants at the TERT-CLPTM1L locus associate with many cancer types. Nat. Genet. 2009, 41, 221–227. [Google Scholar] [CrossRef]
- Wolpin, B.M.; Rizzato, C.; Kraft, P.; Kooperberg, C.; Petersen, G.M.; Wang, Z.; Arslan, A.A.; Beane-Freeman, L.; Bracci, P.M.; Buring, J.; et al. Genome-wide association study identifies multiple susceptibility loci for pancreatic cancer. Nat. Genet. 2014, 46, 994–1000. [Google Scholar] [CrossRef]
- Huo, D.; Feng, Y.; Haddad, S.; Zheng, Y.; Yao, S.; Han, Y.J.; Ogundiran, T.O.; Adebamowo, C.; Ojengbede, O.; Falusi, A.G.; et al. Genome-wide association studies in women of African ancestry identified 3q26.21 as a novel susceptibility locus for oestrogen receptor negative breast cancer. Hum. Mol. Genet. 2016, 25, 4835–4846. [Google Scholar] [CrossRef] [Green Version]
- Michailidou, K.; Beesley, J.; Lindstrom, S.; Canisius, S.; Dennis, J.; Lush, M.J.; Maranian, M.J.; Bolla, M.K.; Wang, Q.; Shah, M.; et al. Genome-wide association analysis of more than 120,000 individuals identifies 15 new susceptibility loci for breast cancer. Nat. Genet. 2015, 47, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Michailidou, K.; Lindstrom, S.; Dennis, J.; Beesley, J.; Hui, S.; Kar, S.; Lemacon, A.; Soucy, P.; Glubb, D.; Rostamianfar, A.; et al. Association analysis identifies 65 new breast cancer risk loci. Nature 2017, 551, 92–94. [Google Scholar] [CrossRef] [Green Version]
- He, G.; Song, T.; Zhang, Y.; Chen, X.; Xiong, W.; Chen, H.; Sun, C.; Zhao, C.; Chen, Y.; Wu, H. TERT rs10069690 polymorphism and cancers risk: A meta-analysis. Mol. Genet. Genom. Med. 2019, 7, e00903. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.Y.; Dong, Y.L.; Feng, Y.; Zhang, Z.; Cao, X.Z. Polymorphisms in the telomerase reverse transcriptase promoter are associated with risk of breast cancer: A meta-analysis. J. Cancer Res. Ther. 2016, 12, 1040–1044. [Google Scholar] [CrossRef]
- Lilyquist, J.; Ruddy, K.J.; Vachon, C.M.; Couch, F.J. Common Genetic Variation and Breast Cancer Risk-Past, Present, and Future. Cancer Epidemiol. Biomark. Prev. 2018, 27, 380–394. [Google Scholar] [CrossRef] [Green Version]
- Killedar, A.; Stutz, M.D.; Sobinoff, A.P.; Tomlinson, C.G.; Bryan, T.M.; Beesley, J.; Chenevix-Trench, G.; Reddel, R.R.; Pickett, H.A. A Common Cancer Risk-Associated Allele in the hTERT Locus Encodes a Dominant Negative Inhibitor of Telomerase. PLoS Genet. 2015, 11, e1005286. [Google Scholar] [CrossRef] [PubMed]
- Phan, L.; Jin, Y.; Zhang, H.; Qiang, W.; Shekhtman, E.; Shao, D.; Revoe, D.; Villamarin, R.; Ivanchenko, E.; Kimura, M.; et al. ALFA: Allele Frequency Aggregator. Available online: www.ncbi.nlm.nih.gov/snp/docs/gsr/alfa/ (accessed on 14 June 2021).
- Shay, J.W. Role of Telomeres and Telomerase in Aging and Cancer. Cancer Discov. 2016, 6, 584–593. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, M.; Leclercq, G. Relevance of breast cancer cell lines as models for breast tumours: An update. Breast Cancer Res. Treat. 2004, 83, 249–289. [Google Scholar] [CrossRef]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.P.; Tong, F.; et al. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef]
- Dratwa, M.; Wysoczanska, B.; Lacina, P.; Kubik, T.; Bogunia-Kubik, K. TERT-Regulation and Roles in Cancer Formation. Front. Immunol. 2020, 11, 589929. [Google Scholar] [CrossRef] [PubMed]
- Darlix, A.; Louvel, G.; Fraisse, J.; Jacot, W.; Brain, E.; Debled, M.; Mouret-Reynier, M.A.; Goncalves, A.; Dalenc, F.; Delaloge, S.; et al. Impact of breast cancer molecular subtypes on the incidence, kinetics and prognosis of central nervous system metastases in a large multicentre real-life cohort. Br. J. Cancer 2019, 121, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wang, R.L.; Liu, J.J.; Zhou, J.; Li, X.; Hu, W.W.; Jiang, W.J.; Hao, N.B. The prognostic significance of hTERT overexpression in cancers: A systematic review and meta-analysis. Medicine 2018, 97, e11794. [Google Scholar] [CrossRef]
- Shay, J.W.; Wright, W.E. Telomeres and telomerase: Three decades of progress. Nat. Rev. Genet. 2019, 20, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Dratwa, M.; Wysoczanska, B.; Brankiewicz, W.; Stachowicz-Suhs, M.; Wietrzyk, J.; Matkowski, R.; Ekiert, M.; Szelachowska, J.; Maciejczyk, A.; Szajewski, M.; et al. Relationship between Telomere Length, TERT Genetic Variability and TERT, TP53, SP1, MYC Gene Co-Expression in the Clinicopathological Profile of Breast Cancer. Int. J. Mol. Sci. 2022, 23, 5164. [Google Scholar] [CrossRef]
- Vogelstein, B.; Lane, D.; Levine, A.J. Surfing the p53 network. Nature 2000, 408, 307–310. [Google Scholar] [CrossRef]
- Bender, R.; Lange, S. Adjusting for multiple testing—When and how? J. Clin. Epidemiol. 2001, 54, 343–349. [Google Scholar] [CrossRef]
- Stampfer, M.R.; Bartley, J.C. Human mammary epithelial cells in culture: Differentiation and transformation. Cancer Treat. Res. 1988, 40, 1–24. [Google Scholar]
- Pacher, M.; Seewald, M.J.; Mikula, M.; Oehler, S.; Mogg, M.; Vinatzer, U.; Eger, A.; Schweifer, N.; Varecka, R.; Sommergruber, W.; et al. Impact of constitutive IGF1/IGF2 stimulation on the transcriptional program of human breast cancer cells. Carcinogenesis 2007, 28, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Friesenhengst, A.; Pribitzer-Winner, T.; Schreiber, M. Association of the G473A polymorphism and expression of lysyl oxidase with breast cancer risk and survival in European women: A hospital-based case-control study. PLoS ONE 2014, 9, e105579. [Google Scholar] [CrossRef]
- Proestling, K.; Hebar, A.; Pruckner, N.; Marton, E.; Vinatzer, U.; Schreiber, M. The Pro Allele of the p53 Codon 72 Polymorphism Is Associated with Decreased Intratumoral Expression of BAX and p21, and Increased Breast Cancer Risk. PLoS ONE 2012, 7, e47325. [Google Scholar] [CrossRef] [PubMed]
- Taubenschuß, E.; Marton, E.; Mogg, M.; Frech, B.; Ehart, L.; Muin, D.; Schreiber, M. The L10P Polymorphism and Serum Levels of Transforming Growth Factor β1 in Human Breast Cancer. Int. J. Mol. Sci. 2013, 14, 15376–15385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesenhengst, A.; Pribitzer-Winner, T.; Miedl, H.; Pröstling, K.; Schreiber, M. Elevated Aromatase (CYP19A1) Expression Is Associated with a Poor Survival of Patients with Estrogen Receptor Positive Breast Cancer. Horm. Cancer 2018, 9, 128–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miedl, H.; Dietrich, B.; Kaserer, K.; Schreiber, M. The 40bp Indel Polymorphism rs150550023 in the MDM2 Promoter is Associated with Intriguing Shifts in Gene Expression in the p53-MDM2 Regulatory Hub. Cancers 2020, 12, 3363. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009. [Google Scholar]
- Harrington, D.P.; Fleming, T.R. A Class of Rank Test Procedures for Censored Survival-Data. Biometrika 1982, 69, 553–566. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes/ Alleles | Unadjusted | Adjusted for Age | ||
|---|---|---|---|---|
| OR (95% CI) | p-Value | OR (95% CI) | p-Value | |
| TT vs. CC | 0.92 (0.51–1.67) | 0.793 | 0.99 (0.51–1.91) | 0.957 |
| TT vs. CT | 0.93 (0.51–1.69) | 0.801 | 0.95 (0.48–1.89) | 0.885 |
| TT vs. CT + CC | 0.92 (0.52–1.64) | 0.788 | 0.97 (0.51–1.84) | 0.929 |
| CT vs. CC | 1.00 (0.72–1.39) | 0.990 | 1.04 (0.72–1.51) | 0.833 |
| TT + CT vs. CC | 0.99 (0.72–1.35) | 0.927 | 1.03 (0.72–1.47) | 0.867 |
| T vs. C | 0.98 (0.76–1.25) | 0.851 | 1.01 (0.77–1.34) | 0.927 |
| Subgroup | Patient No. (%) | TT vs. CC | T vs. C | |||
|---|---|---|---|---|---|---|
| OR (95% CI) | p | OR (95% CI) | p | |||
| age (years) | <55 | 171 (42.4%) | 1.13 (0.56–2.27) | 0.659 | 1.00 (0.74–1.35) | 0.993 |
| ≥55 | 232 (57.6%) | 0.76 (0.38–1.55) | 0.425 | 0.96 (0.72–1.28) | 0.775 | |
| tumor type | ductal | 245 (75.4%) | 1.00 (0.52–1.92) | 0.868 | 1.00 (0.76–1.31) | 0.977 |
| lobular | 80 (24.6%) | 0.61 (0.22–1.71) | 0.413 | 0.73 (0.48–1.10) | 0.129 | |
| lymph node status | pN0 | 193 (59.4%) | 1.20 (0.61–2.35) | 0.548 | 1.08 (0.81–1.44) | 0.618 |
| pN+ | 132 (40.6%) | 0.48 (0.19–1.25) | 0.112 | 0.78 (0.55–1.11) | 0.165 | |
| ER status | pos | 270 (69.8%) | 0.66 (0.33–1.32) | 0.257 | 0.88 (0.66–1.15) | 0.345 |
| neg | 117 (30.2%) | 1.63 (0.78–3.40) | 0.208 | 1.20 (0.86–1.67) | 0.275 | |
| PR status | pos | 186 (48.9%) | 0.64 (0.29–1.39) | 0.301 | 0.84 (0.62–1.14) | 0.256 |
| neg | 194 (51.1%) | 1.22 (0.62–2.41) | 0.546 | 1.11 (0.83–1.48) | 0.485 | |
| HER2 status | pos | 74 (20.2%) | 1.08 (0.43–2.73) | 0.723 | 0.99 (0.66–1.48) | 0.960 |
| neg | 293 (79.8%) | 0.94 (0.49–1.77) | 0.808 | 1.00 (0.77–1.31) | 0.976 | |
| Ki67 pos cells | <50% | 278 (90.8%) | 0.76 (0.39–1.48) | 0.449 | 0.90 (0.69–1.18) | 0.445 |
| ≥50% | 28 (9.2%) | 2.66 (0.84–8.44) | 0.110 | 1.56 (0.88–2.75) | 0.133 | |
| molecular subtype | luminal | 292 (75.8%) | 0.73 (0.37–1.41) | 0.354 | 0.89 (0.68–1.17) | 0.416 |
| HER2 type | 33 (8.6%) | 1.46 (0.45–4.81) | 0.414 | 1.13 (0.65–1.95) | 0.673 | |
| triple neg | 60 (15.6%) | 1.87 (0.75–4.71) | 0.155 | 1.35 (0.88–2.05) | 0.167 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zins, K.; Peka, E.; Miedl, H.; Ecker, S.; Abraham, D.; Schreiber, M. Association of the Telomerase Reverse Transcriptase rs10069690 Polymorphism with the Risk, Age at Onset and Prognosis of Triple Negative Breast Cancer. Int. J. Mol. Sci. 2023, 24, 1825. https://doi.org/10.3390/ijms24031825
Zins K, Peka E, Miedl H, Ecker S, Abraham D, Schreiber M. Association of the Telomerase Reverse Transcriptase rs10069690 Polymorphism with the Risk, Age at Onset and Prognosis of Triple Negative Breast Cancer. International Journal of Molecular Sciences. 2023; 24(3):1825. https://doi.org/10.3390/ijms24031825
Chicago/Turabian StyleZins, Karin, Elisabeth Peka, Heidi Miedl, Stefanie Ecker, Dietmar Abraham, and Martin Schreiber. 2023. "Association of the Telomerase Reverse Transcriptase rs10069690 Polymorphism with the Risk, Age at Onset and Prognosis of Triple Negative Breast Cancer" International Journal of Molecular Sciences 24, no. 3: 1825. https://doi.org/10.3390/ijms24031825
APA StyleZins, K., Peka, E., Miedl, H., Ecker, S., Abraham, D., & Schreiber, M. (2023). Association of the Telomerase Reverse Transcriptase rs10069690 Polymorphism with the Risk, Age at Onset and Prognosis of Triple Negative Breast Cancer. International Journal of Molecular Sciences, 24(3), 1825. https://doi.org/10.3390/ijms24031825