

The Influence of Solid and Liquid Systems In Vitro on the Growth and Biosynthetic Characteristics of Microshoot Culture of Spiraea betulifolia ssp. aemiliana
 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
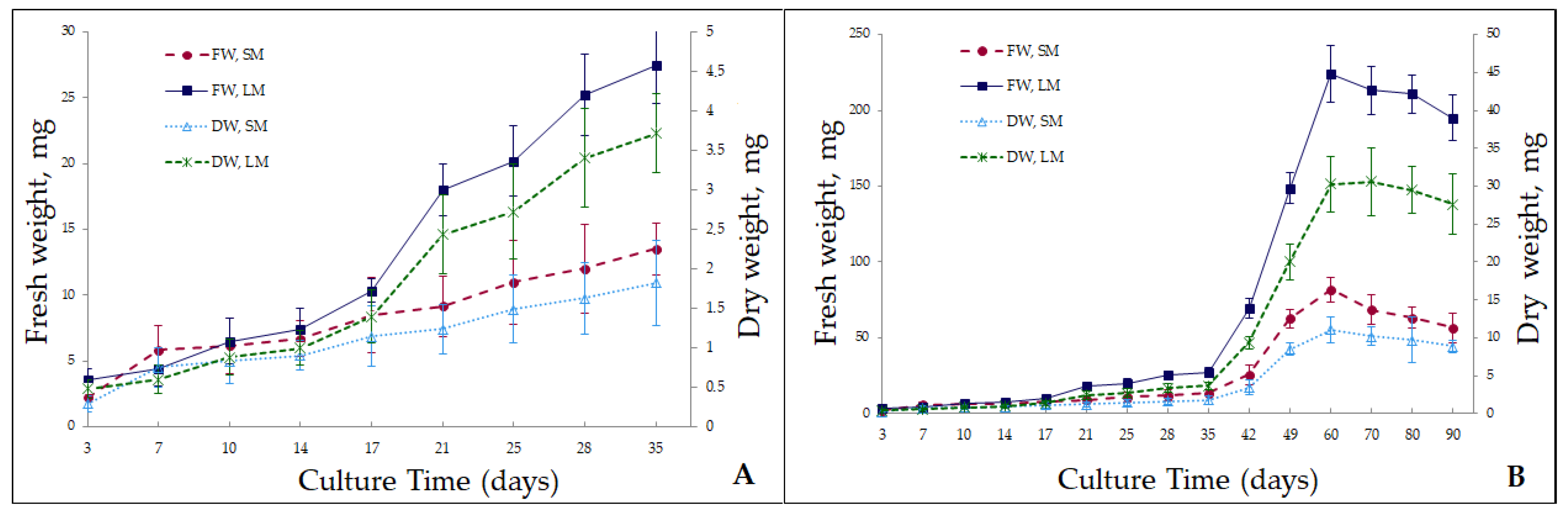
2.1. Microshoot Propagation in Liquid and Solid Media
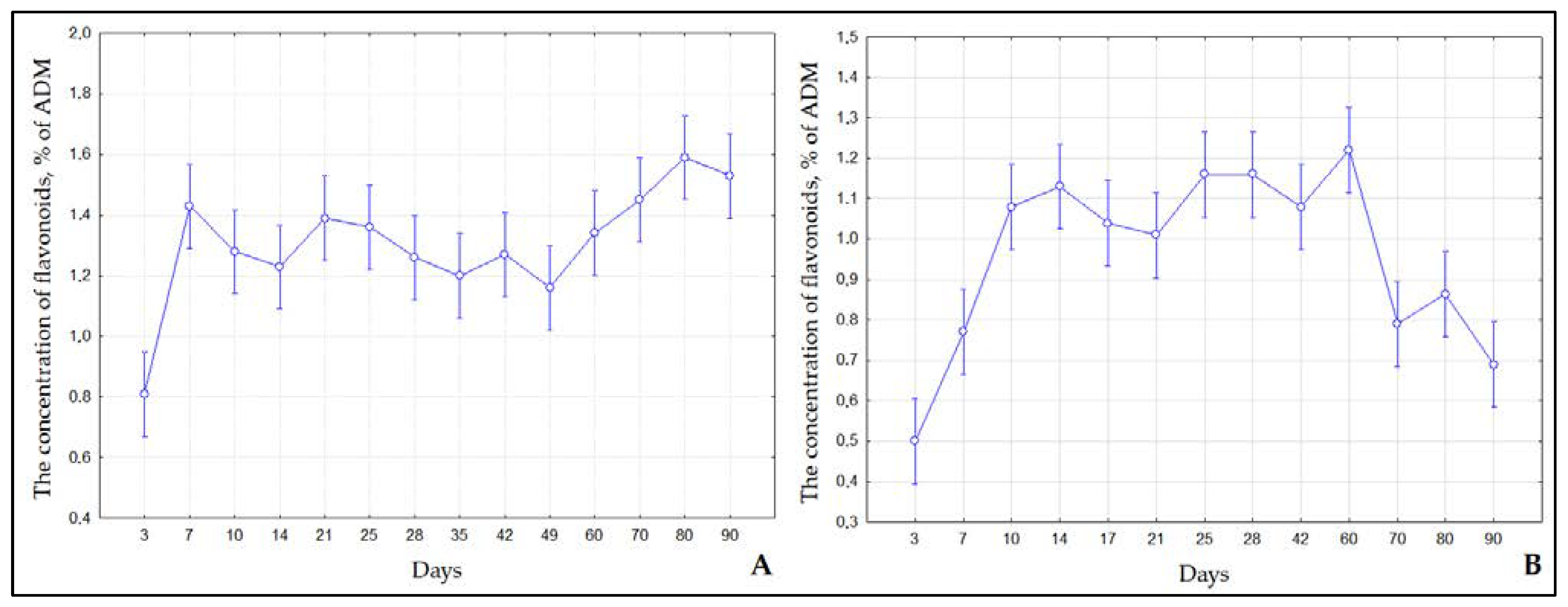
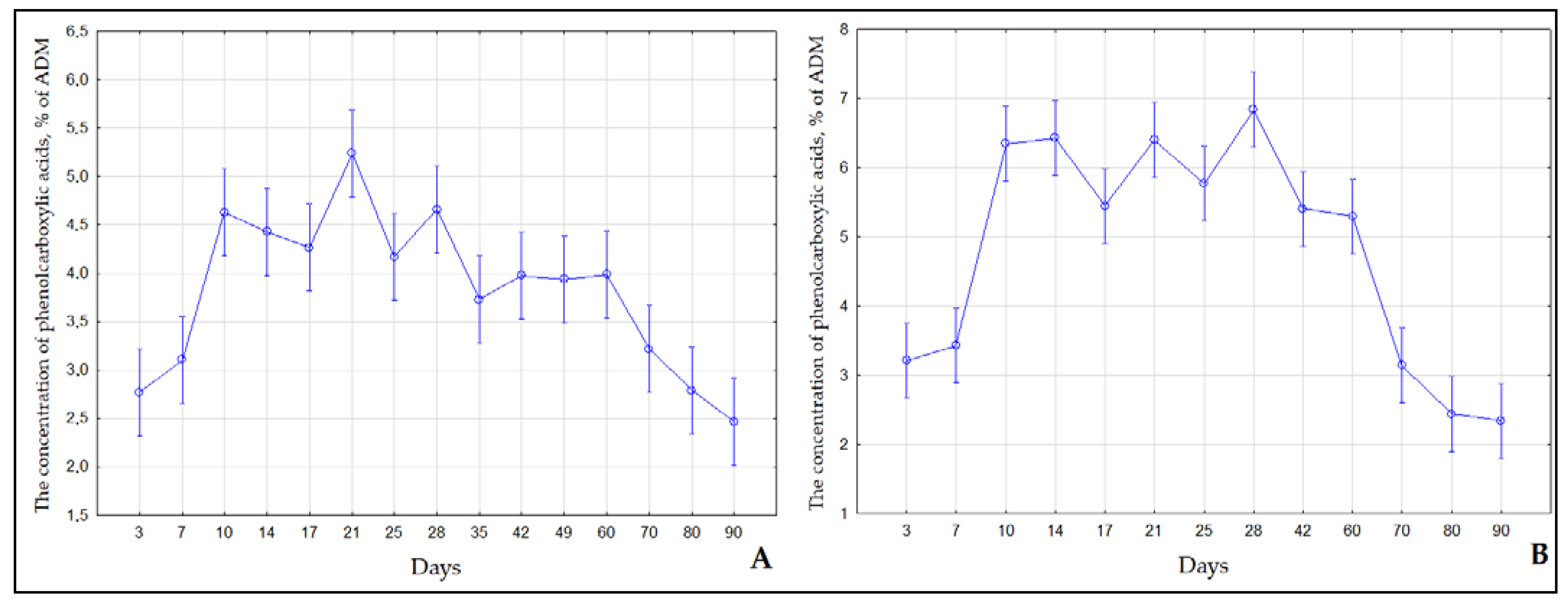
2.2. Contents of Flavonoids and Phenol Carboxylic Acids in S. Betulifolia Ssp. Aemiliana In Vitro and In Vivo
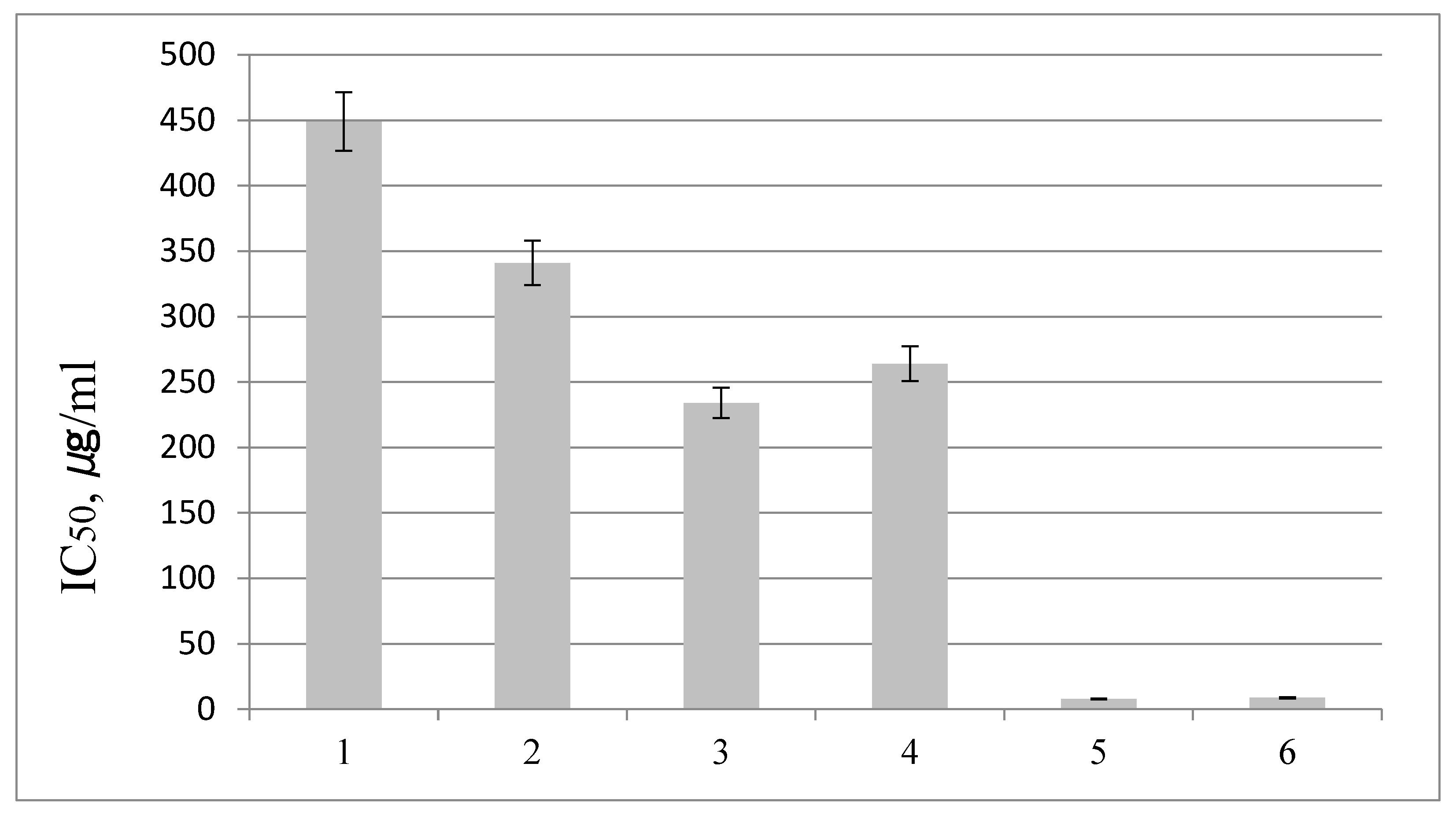
2.3. Antioxidant Activity of S. Betulifolia Ssp. Aemiliana In Vitro and In Vivo
3. Materials and Methods
3.1. Plant Material
3.2. Examination of Culture Growth Dynamics in Solid and Liquid Media
3.3. Plant Material for Phytochemical Assays
3.4. Extract Preparation
3.5. The Total Flavonoid Content
3.6. The Total Phenolic Acid Content
3.7. Quantitation of Individual Phenolic Compounds via HPLC
3.8. Estimation of Antiradical Activity
3.9. Chemicals
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dziggel, C.; Schafer, H.; Wink, M. Tools of pathway reconstruction and production of economically relevant plant secondary metabolites in recombinant microorganisms. Biotechnol. J. 2017, 12, 1600145. [Google Scholar] [CrossRef]
- Chen, S.L.; Yu, H.; Luo, H.M.; Wu, Q.; Li, C.F.; Steinmetz, A. Conservation and sustainable use of medicinal plants: Problems, progress, and prospects. Chin. Med. 2016, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.R.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar]
- Martin, K.P.; Joseph, D.; Madassery, J.; Philip, V.J. Direct shoot regeneration from lamina explants of two commercial cut flower cultivars of Anthurium andreanum. Hortic. Sci. 2003, 39, 500–504. [Google Scholar]
- Lai, R.; Lai, S. Role of Tissue culture in rapid clonal propagation and production of pathogen–free plants. In Crop Improvement Utilizing Biotechnology; CRC Press, Taylor & Francis: Boca Raton, FL, USA, 2019; pp. 73–116. [Google Scholar]
- Nigra, H.M.; Caso, O.H.; Giulietti, A.M. Production of solasodine by calli from different parts of Solanum eleagnifolium Cav. plants. Plant Cell Rep. 1987, 6, 135–137. [Google Scholar] [CrossRef]
- Zhao, J.; Zhu, W.-H.; Hu, Q.; Guo, Y.-Q. Compact callus cluster suspension cultures of Catharanthus roseus with enhanced indole alkaloid biosynthesis. In-Vitro Cell Dev Biol.-Plant 2001, 37, 68–72. [Google Scholar] [CrossRef]
- Jha, S.; Sahu, N.P.; Mahato, S.B. Production of the alkaloids emetine and cephaeline in callus cultures of Cephaelis ipecacuanha. Planta Med. 1988, 54, 504–506. [Google Scholar] [CrossRef]
- Kreis, W. In-Vitro Culturing Techniques of Medicinal Plants. In Medicinal Plant Biotechnology. From Basic Research to Industrial Applications; Kayser, O., Quax, W.J., Eds.; Wiley-VCH: Weinheim, Germany, 2007; pp. 159–185. [Google Scholar]
- Toppel, G.; Witte, L.; Riebesehl, B.; von Borstel, K.; Hartmann, T. Alkaloid patterns and biosynthetic capacity of root cultures from some pyrrolizidine alkaloid-producing Senecio species. Plant Cell Rep. 1987, 6, 466–469. [Google Scholar] [CrossRef]
- Srividya, N.B.; Sridevi, P.; Satyanaraya, P. Azadirachtin and nimbin content in in vitro cultured shoots and roots of Azadirachta indica A Juss. Indian J. Plant Physiol. 1998, 3, 128–129. [Google Scholar]
- Muraseva, D.; Kostikova, V. In vitro and ex vitro rooting of Spiraea betulifolia subsp. aemiliana (Rosaceae), an ornamental shrub. In Proceedings of the Plant Diversity: Status, Trends, Conservation Concept: BIO Web of Conferences, Novosibirsk, Russia, 30 September–3 October 2020; Volume 24, p. 00058. [Google Scholar]
- Muraseva, D.S.; Kostikova, V.A. In vitro propagation of Spiraea betulifolia subsp. aemiliana (Rosaceae) and comparative analysis of phenolic compounds of microclones and intact plants. Plant Cell Tissue Organ Cult. 2021, 144, 493–504. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Yoshida, K.; Hishida, A.; Iida, O.; Hosokawa, K.; Kawabata, J. Flavonol caffeoylglycosides as alpha-glucosidase inhibitors from Spiraea cantoniensis flower. J. Agric. Food Chem. 2008, 56, 4367–4371. [Google Scholar] [CrossRef] [PubMed]
- Kashchenko, N.I.; Chirikova, N.K.; Olennikov, D.N. Acylated flavonoids from Spiraea genus as inhibitors of α-amylase. Russ. J. Bioorg. Chem. 2018, 44, 876–886. [Google Scholar] [CrossRef]
- Kostikova, V.A.; Petrova, N.V. Phytoconstituents and Bioactivity of Plants of the Genus Spiraea L. (Rosaceae): A Review. Int. J. Mol. Sci. 2021, 22, 11163. [Google Scholar] [CrossRef] [PubMed]
- Storozhenko, N.D. Polyphenol Compounds of Spiraea hypericifolia L. Ph.D. Thesis, Irkutsk Institute of Organic Chemistry SB of RAS, Irkutsk, Russia, 1977. (In Russian). [Google Scholar]
- Mirovich, V.M.; Krivosheev, I.M.; Gordeeva, V.V.; Tsyrenzhapov, A.V. Method for Producing Agent Possessing Anti-Inflammatory, Diuretic and Antioxidant Activity. Russian Patent RU 2542493C1, 8 November 2013. (In Russian). [Google Scholar]
- Kostikova, V.A.; Zarubaev, V.V.; Esaulkova, I.L.; Sinegubova, E.O.; Kadyrova, R.A.; Shaldaeva, T.M.; Veklich, T.N.; Kuznetsov, A.A. The antiviral, antiradical, and phytochemical potential of dry extracts from Spiraea hypericifolia, S. media, and S. salicifolia (Rosaceae). S. Afr. J. Bot. 2022, 147, 215–222. [Google Scholar] [CrossRef]
- Park, J.H.; Ahn, E.K.; Ko, H.J.; Hwang, M.H.; Cho, Y.R.; Lee, D.R.; Choi, B.K.; Seo, D.W.; Oh, J.S. Spiraea prunifolia leaves extract inhibits adipogenesis and lipogenesis by promoting β-oxidation in high fat diet-induced obese mice. Biomed. Pharmacother. 2022, 149, 112889. [Google Scholar]
- Densco, I. Factors influencing vitrification of carnation and conifers. Acta Hortic. 1987, 212, 167–176. [Google Scholar]
- Kevers, C.; Prat, R.; Gaspar, T.H. Vitrification of carnation in vitro: Changes in the cell wall mechanical properties, cellulose and lignin content. Plant Growth Regul. 1987, 5, 59–66. [Google Scholar] [CrossRef]
- Böttcher, I.; Zoglauer, K.; Goring, H. Induction and reversion ofvitrification of plants cultured in vitro. Physiol. Plant. 1988, 72, 560–654. [Google Scholar]
- Skidmore, D.I.; Simons, A.J.; Bedi, S. In vitro culture of shoots of Pinus caribaea on a liquid medium. Plant Cell Tissue Organ Cult. 1988, 14, 129–136. [Google Scholar] [CrossRef]
- Dillen, W.; Buysens, S.A. Simple technique to overcome vitrification in Gypsophila paniculata L. Plant Cell Tissue Organ Cult. 1989, 19, 181–188. [Google Scholar] [CrossRef]
- Alvard, D.; Cote, F.; Teisson, C. Comparison of methods of liquid medium culture for banana micropropagation: Effects of temporary immersion of explants. Plant Cell Tissue Organ Cult. 1993, 32, 55–60. [Google Scholar] [CrossRef]
- Levin, R.; Stav, R.; Alper, Y.; Watad, A.A. A technique for repeated axenic subculture of plant tissues in a biroeactor on liquid medium containing sucrose. Plant Tissue Cult. Biotechnol. 1997, 3, 41–45. [Google Scholar]
- Ziv, M.; Ronen, G.; Raviv, M. Proliferation of meristematic clusters in disposable presterlized plastic bioreactors for large scale micropropagation of plants. In Vitro Cell Dev. Biol. Plant 1998, 34, 152–158. [Google Scholar] [CrossRef]
- Ascough, G.D.; Fennell, C.W. The regulation of plant growth and development in liquid culture. S. Afr. J. Bot. 2004, 70, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.Y.; Knight, S.L.; Smith, M.A.L. Effect of liquid culture on the growth and development of minature rose (Rosa chinensis Jacq. ‘Minima’). Plant Cell Tissue Organ Cult. 1993, 32, 329–334. [Google Scholar] [CrossRef]
- Akin, M.; Eyduran, E.; Reed, B.M. Use of RSM and CHAID data mining algorithm for predicting mineral nutrition of hazelnut. Plant Cell Tissue Organ Cult. 2017, 128, 303–316. [Google Scholar] [CrossRef]
- Akin, M.; Eyduran, S.P.; Eyduran, E.; Reed, B.M. Analysis of macro nutrient related growth responses using multivariate adaptive regression splines. Plant Cell Tissue Organ Cult. 2020, 140, 661–670. [Google Scholar] [CrossRef]
- Barz, W.H.; Hoesel, W. Metabolism and Degradation of Phenolic Compounds in Plants. In Biochemistry of Plant Phenolics. Recent Advances in Phytochemistry; Swain, T., Harbone, J.B., van Sumere, C.F., Eds.; Springer: Boston, MA, USA, 1979; Volume 12. [Google Scholar]
- Antonova, G.F.; Zheleznichenko, T.V.; Stasova, V.V. Changes in Content and Composition of Phenolic Acids during Growth of Xylem Cells of Scots Pine. Russ. J. Dev. Biol. 2011, 42, 238–246. [Google Scholar] [CrossRef]
- Zubova, M.; Nechaeva, T.; Kartashov, A.; Zagoskina, N. Regulation of the phenolic compounds accumulation in the tea-plant callus culture with a separate and combined effect of light and cadmium ions. Biol. Bull. 2020, 47, 593–604. [Google Scholar] [CrossRef]
- Goncharuk, E.A.; Zubova, M.Y.; Nechaeva, T.L.; Kazantseva, V.V.; Gulevich, A.A.; Baranova, E.N.; Lapshin, P.V.; Katanskaya, V.M.; Aksenova, M.A.; Zagoskina, N.V. Effects of Hydrogen Peroxide on In Vitro Cultures of Tea (Camellia sinensis L.) Grown in the Dark and in the Light: Morphology, Content of Malondialdehyde, and Accumulation of Various Polyphenols. Molecules 2022, 27, 6674. [Google Scholar]
- Skała, E.; Olszewska, M.A.; Makowczyńska, J.; Kicel, A. Effect of Sucrose Concentration on Rhaponticum carthamoides (Willd.) Iljin Transformed Root Biomass, Caffeoylquinic Acid Derivative, and Flavonoid Production. Int. J. Mol. Sci. 2022, 23, 13848. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Kumar, S.; Abbat, S.; Nikhil, K.; Sondhi, S.M.; Bharatam, P.V.; Roy, P.; Pruthi, V. Ferulic acid amide derivatives as anticancer and antioxidant agents: Synthesis, thermal, biological and computational studies. Med. Chem. Res. 2016, 25, 1175–1192. [Google Scholar]
- Ali, G.; Neda, G. Flavonoids and phenolic acids: Role and biochemical activity in plants and human. J. Med. Plants Res. 2011, 5, 6697–6703. [Google Scholar]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Kostikova, V.A.; Kuznetsov, A.A. Changes in the sets and levels of flavonoids and Phenolcarboxylic Acids in the Leaves of Spiraea betulifolia subsp. aemiliana (Rosaceae) during Introduction into Novosibirsk Conditions. Chem. Sustain. Dev. 2021, 29, 40–50. [Google Scholar]
- Zaprometov, M. Specialized functions of phenolic compounds in plants. Russ. J. Plant Physiol. 1993, 40, 921–931. [Google Scholar]
- Ryan, K.G.; Swinny, E.E.; Markham, K.R.; Winefield, C. Flavonoid gene expression and UV photoprotection in transgenic and mutant Petunia leaves. Phytochemistry 2002, 59, 23–32. [Google Scholar]
- Iwashina, T.; Murai, Y. Distribution of kaempferol glycosides and their function in plants. In Kaempferol: Chemistry, Natural Occurrences and Health Benefits; Villers, G., Fougere, Y., Eds.; Nova Science Publishers: New York, NY, USA, 2013; pp. 63–98. [Google Scholar]
- Saito, N.; Tatsuzawa, F.; Toki, K.; Shinoda, K.; Shigihara, A.; Honda, T. The blue anthocyanin pigments from the blue flowers of Heliophila coronopifolia L. (Brassicaceae). Phytochemistry 2011, 72, 2219–2229. [Google Scholar]
- Hashiba, K.; Iwashina, T.; Matsumoto, S. Variation in the quality and quantity of flavonoids in the leaves of coastal and inland Campanula punctata. Biochem. Syst. Ecol. 2006, 34, 854–861. [Google Scholar] [CrossRef]
- Olsson, L.C.; Veit, M.; Weissenböck, G.; Bornman, J.F. Differential flavonoid response to enhanced UV-B radiation in Brassica napus. Phytochemistry 1998, 49, 1021–1028. [Google Scholar] [CrossRef]
- Lu, C.-M.; Yang, J.-J.; Wang, P.-Y.; Lin, C.-C. A new acylated flavonol glycoside and antioxidant effects of Hedyotis diffusa. Planta Med. 2000, 66, 374–377. [Google Scholar] [CrossRef]
- Picman, A.K.; Schneider, E.F.; Picman, J. Effect of flavonoids on mycelial growth of Verticillium albo-atrum. Biochem. Syst. Ecol. 1995, 23, 683–693. [Google Scholar] [CrossRef]
- El-Gammal, A.A.; Mansour, R.M.A. Antimicrobial activities of some flavonoid compounds. Zentralbl. Mikrobiol. 1986, 141, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Bassam, B.J.; Djordjevic, M.A.; Redmond, J.W.; Batley, M.; Rolfe, B.G. Identification of a nodD-dependent locus in the Rhizobium strain NGR234 activated by phenolic factors secreted by soybeans and other legumes. Mol. Plant-Microbe Interact. 1988, 1, 161–168. [Google Scholar] [CrossRef]
- Vogt, T.; Pollak, P.; Tarlyn, N.; Taylor, L.P. Pollination- or wound-induced kaempferol accumulation in Petunia stigmas enhances seed production. Plant Cell 1994, 6, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Ringli, C.; Bigler, L.; Kuhn, B.M.; Leiber, R.-M.; Diet, A.; Santelia, D.; Frey, B.; Pollmann, S.; Klein, M. The modified flavonol glycosylation profile in the Arabidopsis rol1 mutants results in alterations in plant growth and cell shape formation. Plant Cell 2008, 20, 1470–1481. [Google Scholar] [CrossRef] [Green Version]
- Cid-Ortega, S.; Monroy-Rivera, J.A. Extraction of kaempferol and its glycosides using supercritical fluids from plant sources: A review. Food Technol. Biotechnol. 2018, 56, 480–493. [Google Scholar] [CrossRef]
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, antioxidant and anti-inflammatory activities of kaempferol and its corresponding glycosides and the enzymatic preparation of kaempferol. PLoS ONE 2018, 13, e0197563. [Google Scholar] [CrossRef]
- Ahmadian, R.; Rahimi, R.; Bahramsoltani, R. Kaempferol: An encouraging flavonoid for COVID-19. Bol. Latinoam. Caribe Plantas Med. Aromat. 2020, 19, 492–494. [Google Scholar] [CrossRef]
- Firoz, A.; Talwar, P. COVID-19 and retinaCOVID-19 and retinal degenerative diseases: Promising link “Kaempferol”. Curr. Opin. Pharmacol. 2022, 64, 102231. [Google Scholar] [CrossRef] [PubMed]
- Vijendra, P.D.; Jayanna, S.G.; Kumar, V.; Sannabommaji, T.; Rajashekar, J.; Gajula, H. Product enhancement of triterpenoid saponins in cell suspension cultures of Leucas aspera Spreng. Ind. Crops Prod. 2020, 156, 112857. [Google Scholar] [CrossRef]
- Brighente, I.M.C.; Dias, M.; Verdi, L.G.; Pizzolatti, M.G. Antioxidant activity and total phenolic content of some Brazilian species. Pharm. Biol. 2007, 45, 156–161. [Google Scholar] [CrossRef]
- van Beek, T.A. Chemical analysis of Ginkgo biloba leaves and extracts. J. Chromatogr. A 2002, 967, 21–55. [Google Scholar] [CrossRef]
- Kumarasamy, Y.; Byres, M.; Cox, P.J.; Jaspars, M.; Nahar, L.; Sarker, S.D. Screening seeds of some Scottish plants for free radical scavenging activity. Phytother. Res. 2007, 21, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Gawron-Gzella, A.; Witkowska-Banaszczak, E.; Bylka, W.; Dudek-Makuch, M.; Odwrot, A.; Skrodzka, N. Chemical composition, antioxidant and antimicrobial activities of Sanguisorba officinalis L. extracts. Pharm. Chem. J. 2016, 50, 244–249. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Name | Detailed Information (Date of Sampling) | Content of Substances, % of ADM | |
|---|---|---|---|
| Flavonoids | Phenol Carboxylic Acids | ||
| IPs | Intact plants brought from Kunashir Island that have been introduced into the experimental field of CSBG SB RAS since 2016 (June 2021) | 3.80 ± 0.31 | 6.47 ± 0.58 |
| APs | Ex vitro–acclimated plants: plants adapted from in vitro culture and grown in open ground that have been introduced into the experimental field of CSBG SB RAS since 2020 (June 2021) | 3.81 ± 0.34 | 7.50 ± 0.67 |
| Peak No. | Compound | Spectral Characteristics (λmax, nm) | Retention Time (tR), min | Content, mg/g of Absolutely Dry Mass | |||
|---|---|---|---|---|---|---|---|
| In Vitro | In Vivo | ||||||
| MshS | MshL | IPs | APs | ||||
| Extracts (solvent system I) | |||||||
| A1 | chlorogenic acid | 244, 300 sh., 330 | 3.2 | 0.32 ± 0.01 b | 0.46 ±0.01 a | 0.46 ± 0.01 a | 0.38 ± 0.02 b |
| A2 | p-coumaric acid | 226, 293, 320 | 7.9 | 0.04 ± 0.02 c | 0.12 ± 0.03 c | 0.42 ± 0.04 b | 0.66 ± 0.04 a |
| A3 | cinnamic acid | 216, 270 | 35.9 | – | – | 0.43 ± 0.08 a | 0.42 ± 0.05 a |
| A4 | phenolic acid 4 | 255, 265 sh., 315 | 43.1 | 0.75 ± 0.07 b | 0.73 ± 0.03 b | 2.76 ± 0.13 a | 2.77 ± 0.10 a |
| Peak No. | Compound | Spectral Characteristics (λmax, nm) | Retention Time (tR), min | Content, mg/g of Absolutely Dry Mass | |||
|---|---|---|---|---|---|---|---|
| In Vitro | In Vivo | ||||||
| MshS | MshL | IPs | APs | ||||
| Extracts (solvent system I) | |||||||
| F1 | taxifolin | 290 | 8.5 | – | 0.12 ± 0.01 b | 0.50 ± 0.07 a | 0.65 ± 0.08 a |
| F2 | hyperoside | 225, 268 sh., 355 | 18.0 | – | – | 1.38 ± 0,14 a | 1.59 ± 0,07 a |
| F3 | isoquercitrin | 259, 266 sh., 358 | 19.3 | – | – | 0.42 ± 0.01 b | 0.81 ± 0.09 a |
| F4 | rutin | 256, 358 | 20.0 | – | – | 0.28 ± 0.01 a | 0.33 ± 0.04 a |
| F5 | flavone 5 | 250, 340 | 23.8 | – | 0.06 ± 0.01 b | 1.09 ± 0.13 a | 0.83 ± 0.18 a |
| F6 | astragalin | 265, 300 sh., 350 | 32.5 | – | 0.12 ± 0.01 b | 0.57 ± 0.09 a | 0.65 ± 0.07 a |
| F7 | quercetin | 255, 372 | 40.6 | 0.13 ± 0.04 b | 0.18 ± 0.01 b | 2.55 ± 0.15 a | 2.98 ± 0.13 a |
| F8 | kaempferol | 266, 370 | 46.9 | 0.31 ± 0.01 a | 0.25 ± 0.01 b | 0.12 ± 0.02 c | 0.14 ± 0.01 c |
| Hydrolysates of extracts (solvent system II) | |||||||
| Aglycone 1 | Quercetin * | 255, 372 | 6.4 | 1.38 ± 0.07 b | 1.24 ± 0.06 b | 9.23 ± 0.19 a | 8.83 ± 0.18 a |
| Aglycone 2 | Kaempferol * | 266, 370 | 11.0 | 1.33 ± 0.04 a | 1.06 ± 0.08 b | 0.78 ± 0.04 c | 0.88 ± 0.06 b,c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheleznichenko, T.V.; Muraseva, D.S.; Erst, A.S.; Kuznetsov, A.A.; Kulikovskiy, M.S.; Kostikova, V.A. The Influence of Solid and Liquid Systems In Vitro on the Growth and Biosynthetic Characteristics of Microshoot Culture of Spiraea betulifolia ssp. aemiliana. Int. J. Mol. Sci. 2023, 24, 2362. https://doi.org/10.3390/ijms24032362
Zheleznichenko TV, Muraseva DS, Erst AS, Kuznetsov AA, Kulikovskiy MS, Kostikova VA. The Influence of Solid and Liquid Systems In Vitro on the Growth and Biosynthetic Characteristics of Microshoot Culture of Spiraea betulifolia ssp. aemiliana. International Journal of Molecular Sciences. 2023; 24(3):2362. https://doi.org/10.3390/ijms24032362
Chicago/Turabian StyleZheleznichenko, Tatiana V., Dinara S. Muraseva, Andrey S. Erst, Alexander A. Kuznetsov, Maxim S. Kulikovskiy, and Vera A. Kostikova. 2023. "The Influence of Solid and Liquid Systems In Vitro on the Growth and Biosynthetic Characteristics of Microshoot Culture of Spiraea betulifolia ssp. aemiliana" International Journal of Molecular Sciences 24, no. 3: 2362. https://doi.org/10.3390/ijms24032362
APA StyleZheleznichenko, T. V., Muraseva, D. S., Erst, A. S., Kuznetsov, A. A., Kulikovskiy, M. S., & Kostikova, V. A. (2023). The Influence of Solid and Liquid Systems In Vitro on the Growth and Biosynthetic Characteristics of Microshoot Culture of Spiraea betulifolia ssp. aemiliana. International Journal of Molecular Sciences, 24(3), 2362. https://doi.org/10.3390/ijms24032362