
Anti-Inflammatory Activity and Mechanism of Sweet Corn Extract on Il-1β-Induced Inflammation in a Human Retinal Pigment Epithelial Cell Line (ARPE-19)

Abstract
:1. Introduction
2. Results
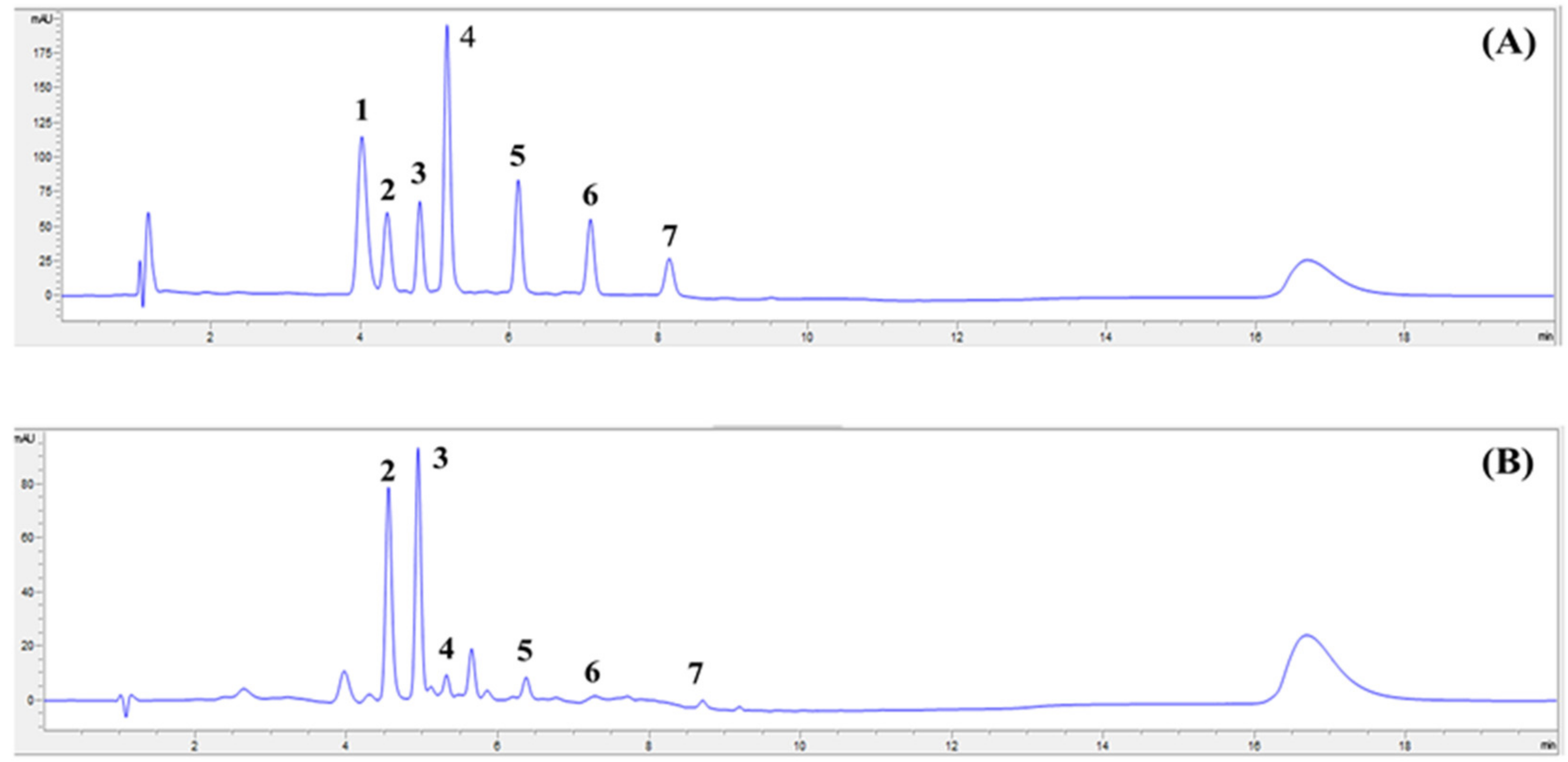
2.1. Sweet Corn Extract Profiled Using HPLC–DAD
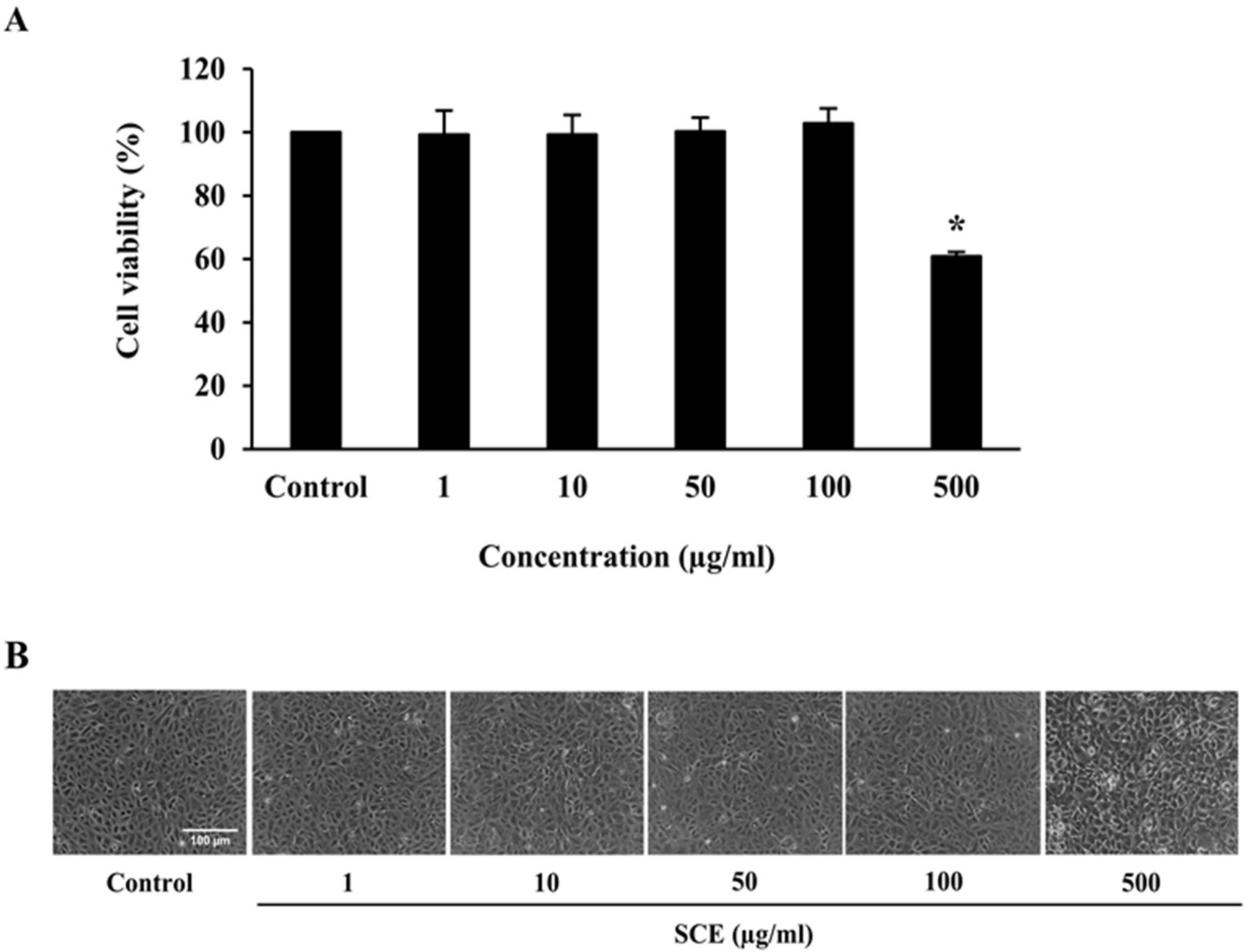
2.2. Effect of SCE on Cell Viability of ARPE-19 Cells
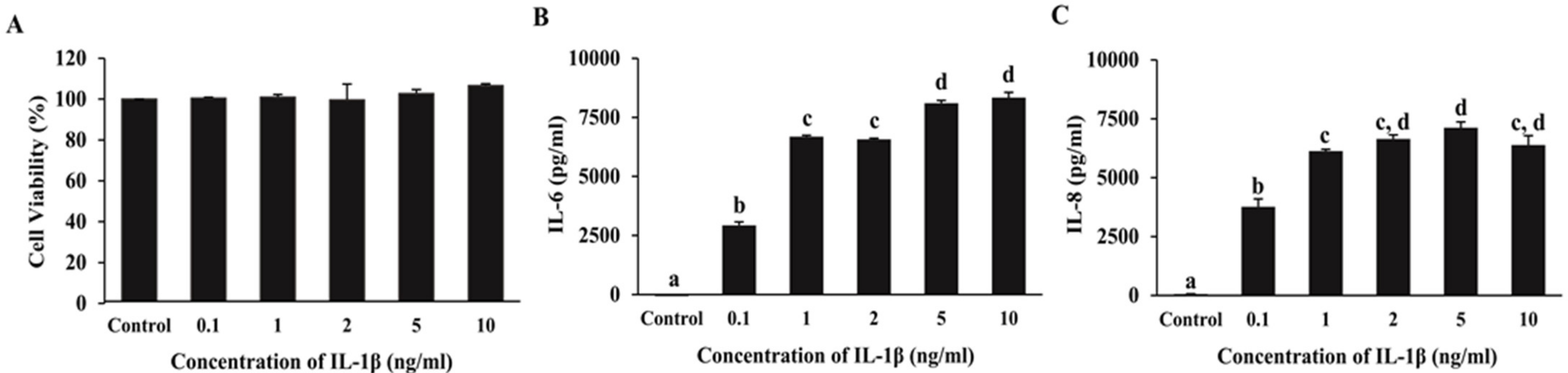
2.3. Effect of IL-1β on IL-6 and IL-8 Secretion and ARPE-19 Cell Viability
2.4. Effect of SCE on IL-6, IL-8, and MCP-1 Secretion in ARPE-19 Cells Induced with IL-1β
2.5. Effect of SCE on iNOS and ICAM-1 Expression in ARPE-19 Cells Induced with IL-1β
2.6. Effect of SCE on the MAPK Signaling Pathway in ARPE-19 Cells Induced with IL-1β
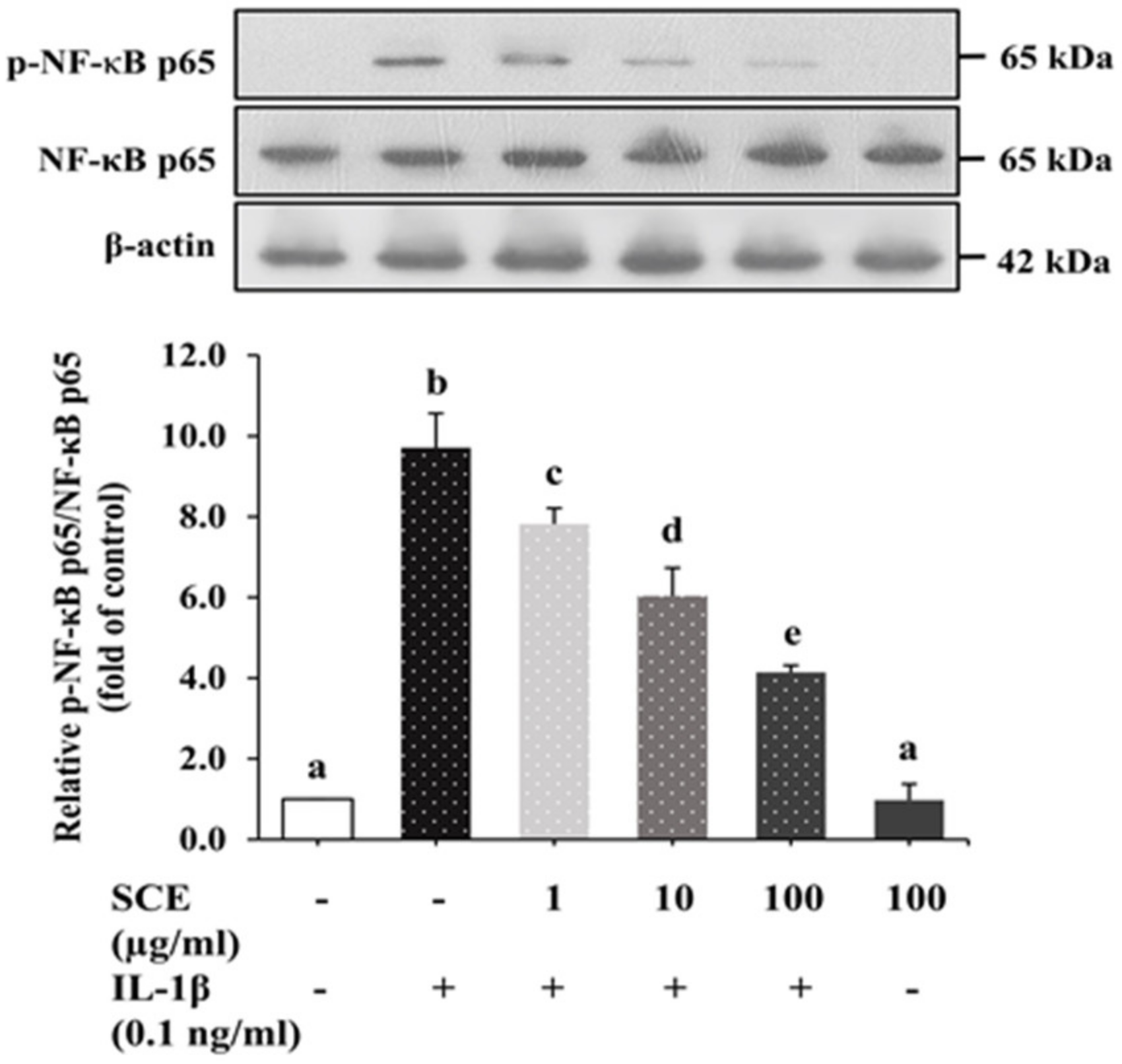
2.7. Effect of SCE on the NF-κB Signaling Pathway in ARPE-19 Cells Induced with IL-1β
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of Sweet Corn Extract (SCE)
4.3. Determination of Carotenoids in SCE Using HPLC–DAD
4.4. ARPE-19 Cells
4.5. Cell Viability
4.6. Evaluation of Cytotoxicity of SCE
4.7. Evaluation of a Suitable IL-β Concentration for Induction of Inflammation
4.8. Evaluation of the Protective Effect of SCE on IL-1β-Induced Inflammation in ARPE-19 Cells
4.9. ELISA Test
4.10. Preparation of Total Protein and Western Immunoblot Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhutto, I.A.; Lutty, G.A. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Asp. Med. 2012, 33, 295–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bressler, N. Age-Related Macular Degeneration Is the Leading Cause of Blindness. JAMA J. Am. Med. Assoc. 2004, 291, 1900–1901. [Google Scholar] [CrossRef] [PubMed]
- Bourne, R.R.; Steinmetz, J.D.; Saylan, M.; Mersha, A.M.; Weldemariam, A.H.; Wondmeneh, T.G.; Sreeramareddy, C.T.; Pinheiro, M.; Yaseri, M.; Yu, C.; et al. Causes of blindness and vision impairment in 2020 and trends over 30 years, and prevalence of avoidable blindness in relation to VISION 2020: The Right to Sight: An analysis for the Global Burden of Disease Study. Lancet Glob. Health 2021, 9, e144–e160. [Google Scholar]
- Jonas, J.B.; Cheung, C.M.G.; Panda-Jonas, S. Updates on the Epidemiology of Age-Related Macular Degeneration. Asia Pac. J. Ophthalmol. 2017, 6, 493–497. [Google Scholar]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106-16. [Google Scholar] [CrossRef] [Green Version]
- Cook, H.L.; Patel, P.J.; Tufail, A. Age-related macular degeneration: Diagnosis and management. Br. Med. Bull. 2008, 85, 127-49. [Google Scholar] [CrossRef]
- Moutray, T.; Chakravarthy, U. Age-related macular degeneration: Current treatment and future options. Ther. Adv. Chronic Dis. 2011, 2, 325-31. [Google Scholar] [CrossRef] [Green Version]
- Kauppinen, A.; Paterno, J.J.; Blasiak, J.; Salminen, A.; Kaarniranta, K. Inflammation and its role in age-related macular degeneration. Cell. Mol. Life Sci. 2016, 73, 1765–1786. [Google Scholar] [CrossRef] [Green Version]
- Dick, A.D. Doyne lecture 2016: Intraocular health and the many faces of inflammation. Eye 2017, 31, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Hartnett, M.E. Regulation of signaling events involved in the pathophysiology of neovascular AMD. Mol. Vis. 2016, 22, 189–202. [Google Scholar]
- Gupta, M.P.; Herzlich, A.A.; Sauer, T.; Chan, C.C. Retinal Anatomy and Pathology. Dev. Ophthalmol. 2016, 55, 7–17. [Google Scholar]
- Naylor, A.; Hopkins, A.; Hudson, N.; Campbell, M. Tight Junctions of the Outer Blood Retina Barrier. Int. J. Mol. Sci. 2019, 21, 211. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, G.M.; Kijlstra, A.; Peek, R.; de Vos, A.F. Retinal pigment epithelium-immune system interactions: Cytokine production and cytokine-induced changes. Prog. Retin Eye Res. 2001, 20, 29–48. [Google Scholar] [CrossRef]
- Jun, S.; Datta, S.; Wang, L.; Pegany, R.; Cano, M.; Handa, J.T. The impact of lipids, lipid oxidation, and inflammation on AMD, and the potential role of miRNAs on lipid metabolism in the RPE. Exp. Eye Res. 2019, 181, 346–355. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef]
- Devine, L.; Lightman, S.L.; Greenwood, J. Role of LFA-1, ICAM-1, VLA-4 and VCAM-1 in lymphocyte migration across retinal pigment epithelial monolayers in vitro. Immunology 1996, 88, 456-62. [Google Scholar] [CrossRef]
- Knickelbein, J.E.; Chan, C.C.; Sen, H.N.; Ferris, F.L.; Nussenblatt, R.B. Inflammatory Mechanisms of Age-related Macular Degeneration. Int. Ophthalmol. Clin. 2015, 55, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Rock, K.L.; Lai, J.J.; Kono, H. Innate and adaptive immune responses to cell death. Immunol. Rev. 2011, 243, 191–205. [Google Scholar] [CrossRef] [Green Version]
- di Giovine, F.S.; Duff, G.W. Interleukin 1: The first interleukin. Immunol. Today 1990, 11, 13–20. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef] [Green Version]
- Henríquez-Olguín, C.; Altamirano, F.; Valladares, D.; López, J.R.; Allen, P.D.; Jaimovich, E. Altered ROS production, NF-κB activation and interleukin-6 gene expression induced by electrical stimulation in dystrophic mdx skeletal muscle cells. Biochim. Biophys. Acta 2015, 1852, 1410-9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siyuan, S.; Tong, L.; Liu, R. Corn phytochemicals and their health benefits. Food Sci. Hum. Wellness 2018, 7, 185–195. [Google Scholar] [CrossRef]
- Moros, E.E.; Darnoko, D.; Cheryan, M.; Perkins, E.G.; Jerrell, J. Analysis of xanthophylls in corn by HPLC. J. Agric. Food Chem. 2002, 50, 5787-90. [Google Scholar] [CrossRef] [PubMed]
- Bian, Q.; Qin, T.; Ren, Z.; Wu, D.; Shang, F. Lutein or Zeaxanthin Supplementation Suppresses Inflammatory Responses in Retinal Pigment Epithelial Cells and Macrophages. Adv. Exp. Med. Biol. 2012, 723, 43–50. [Google Scholar]
- Muangnoi, C.; Phumsuay, R.; Jongjitphisut, N.; Waikasikorn, P.; Sangsawat, M.; Rashatasakhon, P.; Paraoan, L.; Rojsitthisak, P. Protective Effects of a Lutein Ester Prodrug, Lutein Diglutaric Acid, against H2O2-Induced Oxidative Stress in Human Retinal Pigment Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 4722. [Google Scholar] [CrossRef]
- Izumi-Nagai, K.; Nagai, N.; Ohgami, K.; Satofuka, S.; Ozawa, Y.; Tsubota, K.; Umezawa, K.; Ohno, S.; Oike, Y.; Ishida, S. Macular pigment lutein is antiinflammatory in preventing choroidal neovascularization. Arterioscler. Thromb Vasc. Biol. 2007, 27, 2555–2562. [Google Scholar] [CrossRef]
- Dunn, K.C.; Aotaki-Keen, A.E.; Putkey, F.R.; Hjelmeland, L.M. ARPE-19, a human retinal pigment epithelial cell line with differentiated properties. Exp. Eye Res. 1996, 62, 155-70. [Google Scholar] [CrossRef]
- Whitcup, S.M.; Nussenblatt, R.B.; Lightman, S.L.; Hollander, D.A. Inflammation in retinal disease. Int. J. Inflam. 2013, 2013, 724648. [Google Scholar] [CrossRef]
- Detrick, B.; Hooks, J.J. The RPE Cell and the Immune System. In Retinal Pigment Epithelium in Health and Disease; Klettner, A., Dithmar, S., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef] [Green Version]
- Wooff, Y.; Man, S.M.; Aggio-Bruce, R.; Natoli, R.; Fernando, N. IL-1 Family Members Mediate Cell Death, Inflammation and Angiogenesis in Retinal Degenerative Diseases. Front. Immunol. 2019, 10, 1618. [Google Scholar] [CrossRef]
- Chung, R.W.S.; Leanderson, P.; Lundberg, A.K.; Jonasson, L. Lutein exerts anti-inflammatory effects in patients with coronary artery disease. Atherosclerosis 2017, 262, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Seddon, J.M.; Ajani, U.A.; Sperduto, R.D.; Hiller, R.; Blair, N.; Burton, T.C.; Farber, M.D.; Gragoudas, E.S.; Haller, J.; Miller, D.T.; et al. Dietary carotenoids, vitamins A, C, and E, and advanced age-related macular degeneration. Eye Disease Case-Control Study Group. JAMA 1994, 272, 1413–1420. [Google Scholar] [CrossRef]
- Huang, Y.M.; Dou, H.L.; Huang, F.F.; Xu, X.R.; Zou, Z.Y.; Lin, X.M. Effect of supplemental lutein and zeaxanthin on serum, macular pigmentation, and visual performance in patients with early age-related macular degeneration. Biomed. Res. Int. 2015, 2015, 564738. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Hu, J.; Yu, Y.; Li, G.; Guo, X.; Li, T.; Liu, R. Comparison of phenolics, flavonoids, and cellular antioxidant activities in ear sections of sweet corn (Zea mays L. saccharata Sturt): XXXX. J. Food Process. Preserv. 2018, 43, e13855. [Google Scholar] [CrossRef] [Green Version]
- Puangpraphant, S.; Cuevas-Rodríguez, E.-O.; Oseguera-Toledo, M. Chapter 9—Anti-inflammatory and antioxidant phenolic compounds. In Current Advances for Development of Functional Foods Modulating Inflammation and Oxidative Stress; Hernández-Ledesma, B., Martínez-Villaluenga, C., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 165–180. [Google Scholar]
- Scott, C.E.; Eldridge, A.L. Comparison of carotenoid content in fresh, frozen and canned corn. J. Food Compos. Anal. 2005, 18, 551–559. [Google Scholar] [CrossRef]
- Humphries, J.M.; Khachik, F. Distribution of lutein, zeaxanthin, and related geometrical isomers in fruit, vegetables, wheat, and pasta products. J. Agric. Food Chem. 2003, 51, 1322-7. [Google Scholar] [CrossRef]
- Ribaya-Mercado, J.; Blumberg, J. Lutein and Zeaxanthin and Their Potential Roles in Disease Prevention. J. Am. Coll. Nutr. 2005, 23, 567S–587S. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef]
- Qin, T.; Gao, S. Inhibition of Proteasome Activity Upregulates IL-6 Expression in RPE Cells through the Activation of P38 MAPKs. J. Ophthalmol. 2018, 2018, 5392432. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Takeda, A.; Hasegawa, E.; Jo, Y.J.; Arima, M.; Oshima, Y.; Ryoji, Y.; Nakazawa, T.; Yuzawa, M.; Nakashizuka, H.; et al. Interleukin-6 plays a crucial role in the development of subretinal fibrosis in a mouse model. Immunol. Med. 2018, 41, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Lechner, J.; Chen, M.; Hogg, R.E.; Toth, L.; Silvestri, G.; Chakravarthy, U.; Xu, H. Peripheral blood mononuclear cells from neovascular age-related macular degeneration patients produce higher levels of chemokines CCL2 (MCP-1) and CXCL8 (IL-8). J. Neuroinflamm. 2017, 14, 42. [Google Scholar] [CrossRef] [Green Version]
- Jonas, J.B.; Tao, Y.; Neumaier, M.; Findeisen, P. Monocyte Chemoattractant Protein 1, Intercellular Adhesion Molecule 1, and Vascular Cell Adhesion Molecule 1 in Exudative Age-Related Macular Degeneration. Arch. Ophthalmol. 2010, 128, 1281–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Othman, R.; Berbari, S.; Vaucher, E.; Couture, R. Differential Expression of Kinin Receptors in Human Wet and Dry Age-Related Macular Degeneration Retinae. Pharmaceuticals 2020, 13, 130. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.C.; Huang, W.C.; JH, S.P.; Wu, Y.H.; Cheng, C.Y. Quercetin Inhibits the Production of IL-1β-Induced Inflammatory Cytokines and Chemokines in ARPE-19 Cells via the MAPK and NF-κB Signaling Pathways. Int. J. Mol. Sci. 2019, 20, 2957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spekker-Bosker, K.; Ufermann, C.-M.; Oldenburg, M.; Däubener, W.; Eller, S.K. Interplay between IDO1 and iNOS in human retinal pigment epithelial cells. Med. Microbiol. Immunol. 2019, 208, 811–824. [Google Scholar] [CrossRef]
- During, A.; Doraiswamy, S.; Harrison, E.H. Xanthophylls are preferentially taken up compared with beta-carotene by retinal cells via a SRBI-dependent mechanism. J. Lipid Res. 2008, 49, 1715-24. [Google Scholar] [CrossRef] [Green Version]
- Bian, Q.; Gao, S.; Zhou, J.; Qin, J.; Taylor, A.; Johnson, E.J.; Tang, G.; Sparrow, J.R.; Gierhart, D.; Shang, F. Lutein and zeaxanthin supplementation reduces photooxidative damage and modulates the expression of inflammation-related genes in retinal pigment epithelial cells. Free Radic. Biol. Med. 2012, 53, 1298-307. [Google Scholar] [CrossRef] [Green Version]
- Bhosale, P.; Larson, A.J.; Frederick, J.M.; Southwick, K.; Thulin, C.D.; Bernstein, P.S. Identification and characterization of a Pi isoform of glutathione S-transferase (GSTP1) as a zeaxanthin-binding protein in the macula of the human eye. J. Biol. Chem. 2004, 279, 49447-54. [Google Scholar] [CrossRef] [Green Version]
- Silván, J.M.; Reguero, M.; de Pascual-Teresa, S. A protective effect of anthocyanins and xanthophylls on UVB-induced damage in retinal pigment epithelial cells. Food Funct. 2016, 7, 1067–1076. [Google Scholar] [CrossRef]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy--from molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta 2005, 1754, 253–262. [Google Scholar] [CrossRef]
- Chen, X.; Han, R.; Hao, P.; Wang, L.; Liu, M.; Jin, M.; Kong, D.; Li, X. Nepetin inhibits IL-1β induced inflammation via NF-κB and MAPKs signaling pathways in ARPE-19 cells. Biomed. Pharmacother. 2018, 101, 87–93. [Google Scholar] [CrossRef]
- Kyosseva, S.V. Targeting MAPK Signaling in Age-Related Macular Degeneration. Ophthalmol. Eye Dis. 2016, 8, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Pugazhenthi, S.; Zhang, Y.; Bouchard, R.; Mahaffey, G. Induction of an inflammatory loop by interleukin-1β and tumor necrosis factor-α involves NF-kB and STAT-1 in differentiated human neuroprogenitor cells. PLoS ONE 2013, 8, e69585. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Zhang, W.; Cui, Z.-M.; Cui, S.-Y.; Fan, J.-B.; Zhu, X.-H.; Liu, W. Resveratrol alleviates the interleukin-1β-induced chondrocytes injury through the NF-κB signaling pathway. J. Orthop. Surg. Res. 2020, 15, 424. [Google Scholar] [CrossRef]
- Reber, L.; Vermeulen, L.; Haegeman, G.; Frossard, N. Ser276 phosphorylation of NF-kB p65 by MSK1 controls SCF expression in inflammation. PLoS ONE 2009, 4, e4393. [Google Scholar] [CrossRef]
- Terasaki, H.; Kase, S.; Shirasawa, M.; Otsuka, H.; Hisatomi, T.; Sonoda, S.; Ishida, S.; Ishibashi, T.; Sakamoto, T. TNF-α Decreases VEGF Secretion in Highly Polarized RPE Cells but Increases It in Non-Polarized RPE Cells Related to Crosstalk between JNK and NF-κB Pathways. PLoS ONE 2013, 8, e69994. [Google Scholar] [CrossRef]
- Fang, I.M.; Yang, C.H.; Yang, C.M.; Chen, M.S. Linoleic acid-induced expression of inducible nitric oxide synthase and cyclooxygenase II via p42/44 mitogen-activated protein kinase and nuclear factor-kappaB pathway in retinal pigment epithelial cells. Exp. Eye Res. 2007, 85, 667–677. [Google Scholar] [CrossRef]
- Yolanda, S.-M.; Carolina, G.-S.; José, L.R.-D.; Alemán-de la Torre, I. Phenolic Compounds in Maize Grains and Its Nixtamalized Products. In Phenolic Compounds; Marcos, S.-H., Mariana, P.-T., Maria del Rosario, G.-M., Eds.; IntechOpen: Rijeka, Croatia, 2017; p. Ch. 8. [Google Scholar]
- Kohno, M.; Musashi, K.; Ikeda, H.O.; Horibe, T.; Matsumoto, A.; Kawakami, K. Oral administration of ferulic acid or ethyl ferulate attenuates retinal damage in sodium iodate-induced retinal degeneration mice. Sci. Rep. 2020, 10, 8688. [Google Scholar] [CrossRef]
- Sun, X.; Ma, L.; Li, X.; Wang, J.; Li, Y.; Huang, Z. Ferulic acid alleviates retinal neovascularization by modulating microglia/macrophage polarization through the ROS/NF-κB axis. Front. Immunol. 2022, 13, 976729. [Google Scholar] [CrossRef]
- Coleman, H.R.; Chan, C.C.; Ferris, F.L., 3rd; Chew, E.Y. Age-related macular degeneration. Lancet 2008, 372, 1835-45. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, J. Anti-Inflammatory Effects of p-coumaric Acid in LPS-Stimulated RAW264.7 Cells: Involvement of NF-κB and MAPKs Pathways. Med. Chem. 2016, 6, 327–330. [Google Scholar] [CrossRef]
- Lee, H.W.; Zhang, H.; Liang, X.; Ong, C.N. Simultaneous determination of carotenoids, tocopherols and phylloquinone in 12 Brassicaceae vegetables. LWT 2020, 130, 109649. [Google Scholar] [CrossRef]
- Gupta, P.; Sreelakshmi, Y.; Sharma, R. A rapid and sensitive method for determination of carotenoids in plant tissues by high performance liquid chromatography. Plant Methods 2015, 11, 5. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lutein | Zeaxanthin | β-Cryptoxanthin | α-Carotene | β-Carotene |
|---|---|---|---|---|
| 1427.3 ± 2.3 | 2017.8 ± 4.6 | 119.9 ± 0.2 | 88.0 ± 1.2 | 126.3 ± 1.7 |
| Time (min) | %A | %B | %C |
|---|---|---|---|
| 0 | 4 | 81 | 15 |
| 1.5 | 1 | 69 | 30 |
| 5.0 | 0 | 70 | 30 |
| 10.0 | 0 | 0 | 100 |
| 13.0 | 0 | 0 | 100 |
| 13.1 | 4 | 81 | 15 |
| 20.0 | 4 | 81 | 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koraneeyakijkulchai, I.; Phumsuay, R.; Thiyajai, P.; Tuntipopipat, S.; Muangnoi, C. Anti-Inflammatory Activity and Mechanism of Sweet Corn Extract on Il-1β-Induced Inflammation in a Human Retinal Pigment Epithelial Cell Line (ARPE-19). Int. J. Mol. Sci. 2023, 24, 2462. https://doi.org/10.3390/ijms24032462
Koraneeyakijkulchai I, Phumsuay R, Thiyajai P, Tuntipopipat S, Muangnoi C. Anti-Inflammatory Activity and Mechanism of Sweet Corn Extract on Il-1β-Induced Inflammation in a Human Retinal Pigment Epithelial Cell Line (ARPE-19). International Journal of Molecular Sciences. 2023; 24(3):2462. https://doi.org/10.3390/ijms24032462
Chicago/Turabian StyleKoraneeyakijkulchai, Inthra, Rianthong Phumsuay, Parunya Thiyajai, Siriporn Tuntipopipat, and Chawanphat Muangnoi. 2023. "Anti-Inflammatory Activity and Mechanism of Sweet Corn Extract on Il-1β-Induced Inflammation in a Human Retinal Pigment Epithelial Cell Line (ARPE-19)" International Journal of Molecular Sciences 24, no. 3: 2462. https://doi.org/10.3390/ijms24032462
APA StyleKoraneeyakijkulchai, I., Phumsuay, R., Thiyajai, P., Tuntipopipat, S., & Muangnoi, C. (2023). Anti-Inflammatory Activity and Mechanism of Sweet Corn Extract on Il-1β-Induced Inflammation in a Human Retinal Pigment Epithelial Cell Line (ARPE-19). International Journal of Molecular Sciences, 24(3), 2462. https://doi.org/10.3390/ijms24032462