
Effect of Antihypertensive Drug (Chlorothiazide) on Fibrillation of Lysozyme: A Combined Spectroscopy, Microscopy, and Computational Study

 , and
, and
Abstract
:1. Introduction

2. Materials and Methods
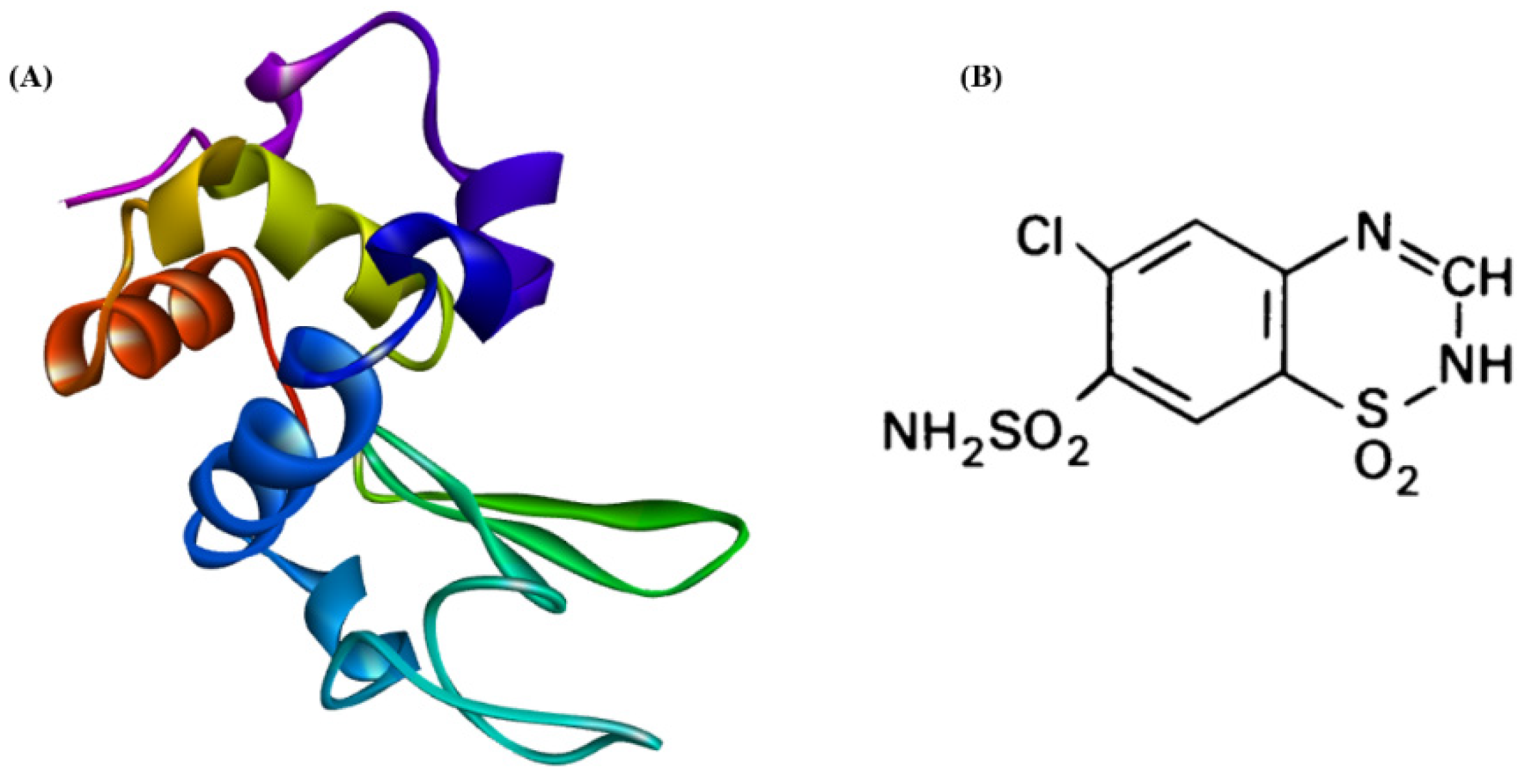
2.1. Materials
2.2. Preparation of HEWL Fibrils
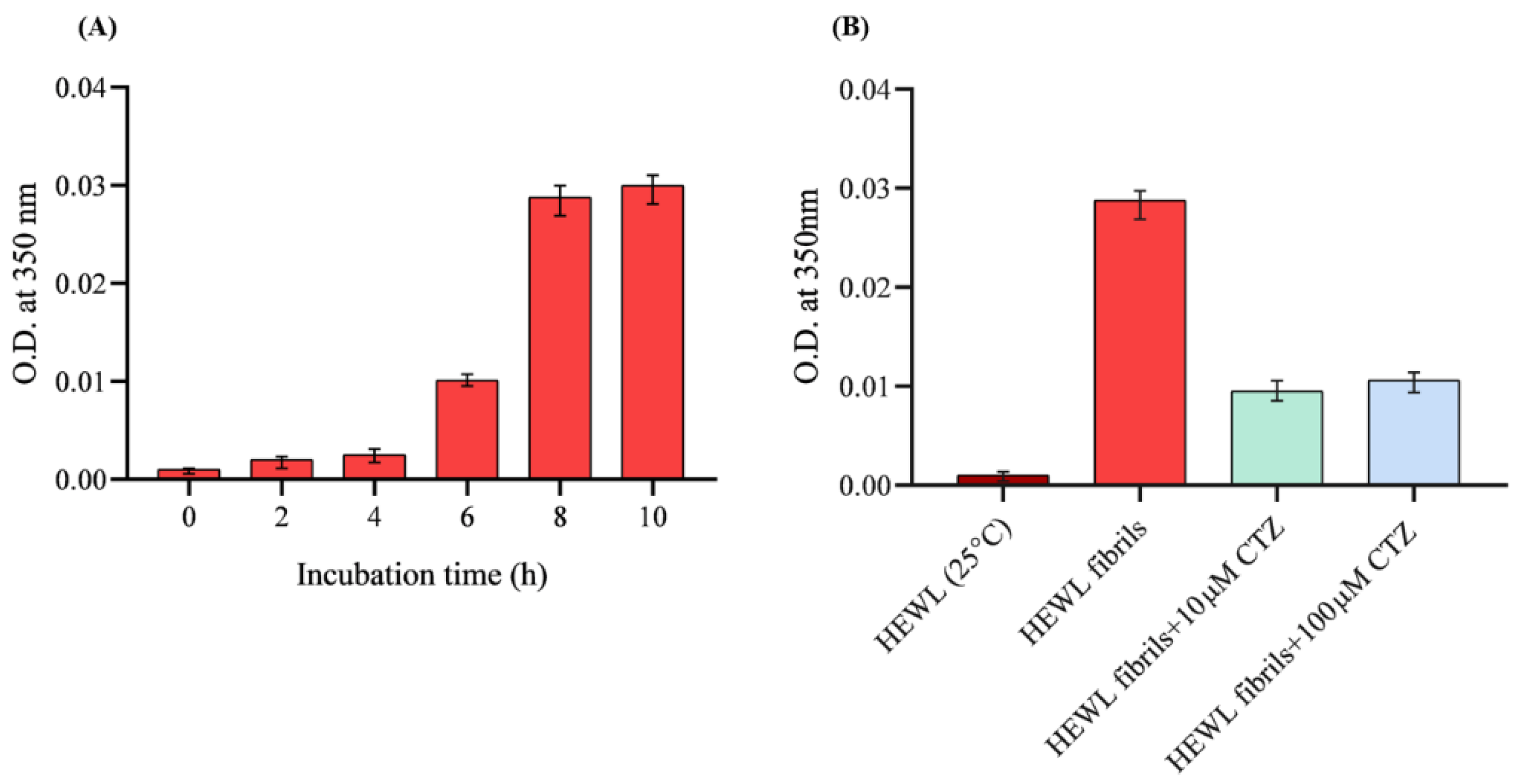
2.3. Turbidity Analysis
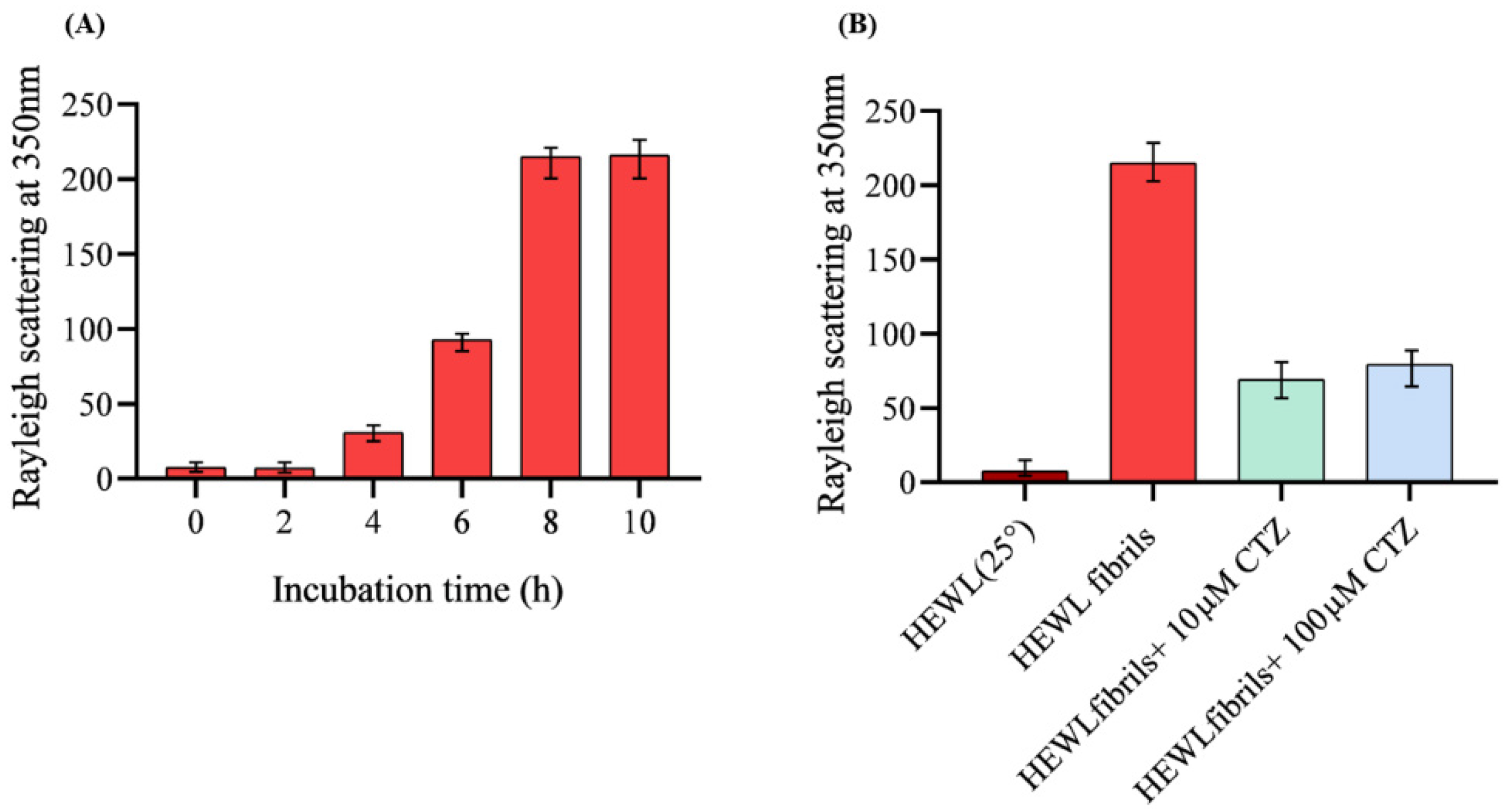
2.4. Rayleigh Light Scattering (RLS) Measurement
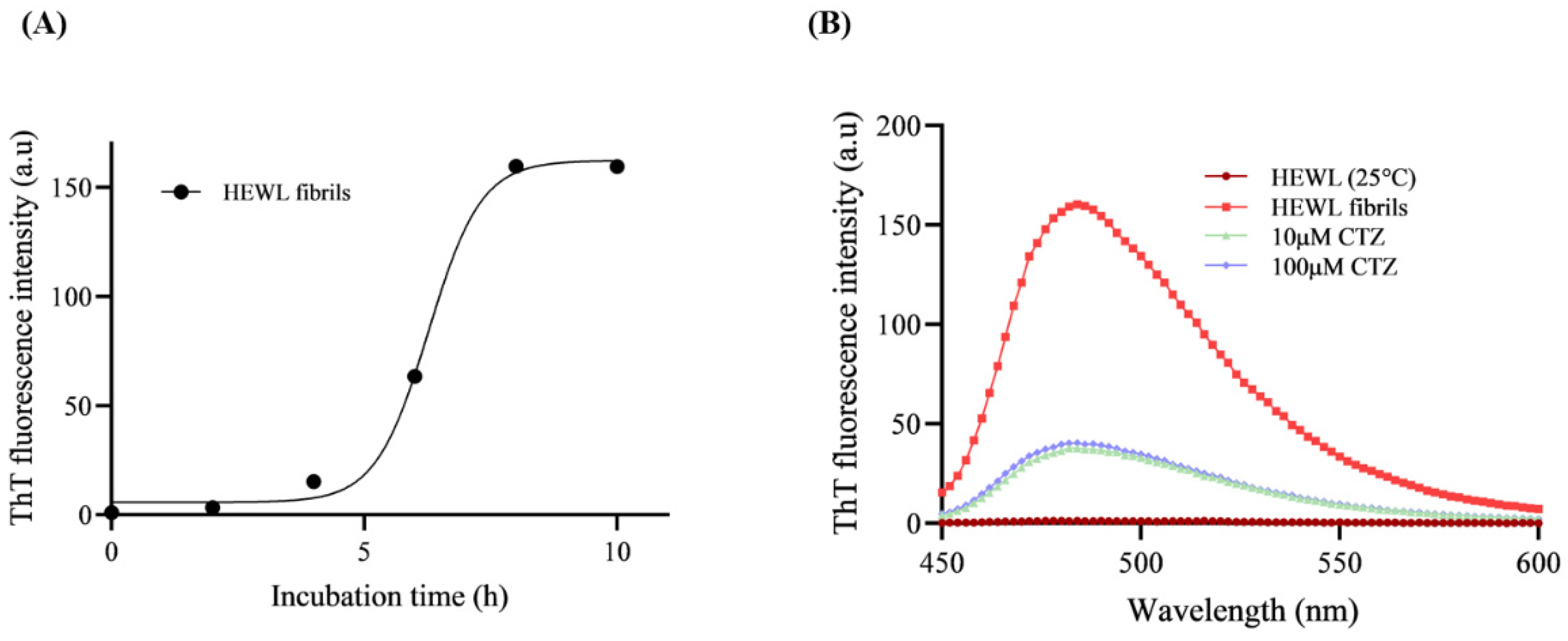
2.5. ThT Fluorescence Assay
2.6. ANS Binding Assay (Hydrophobicity Analysis)
2.7. Far-UV Circular Dichroism
2.8. Steady State Measurement
2.9. Transmission Electron Microscopy (TEM)
2.10. Molecular Modeling Studies
2.10.1. Molecular Docking
2.10.2. Molecular Dynamic Simulation
2.10.3. Statistical Analysis
3. Results and Discussion
3.1. Rayleigh Scattering Measurement (RLS)
3.2. Turbidity Measurement
3.3. ThT Fluorescence Assay
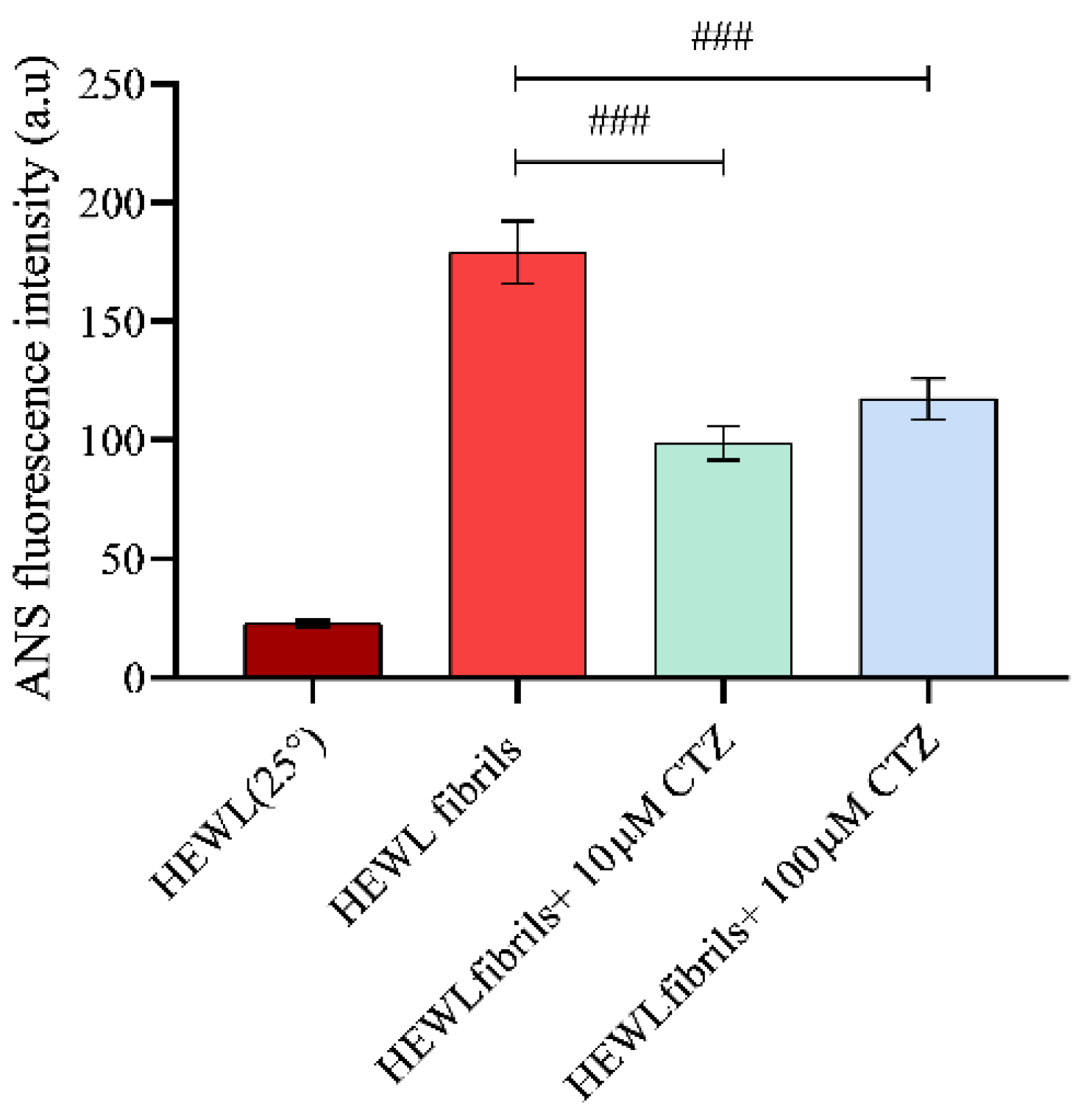
3.4. Effect of CTZ on Surface Hydrophobicity of HEWL
3.5. Circular Dichroism (CD) Measurement
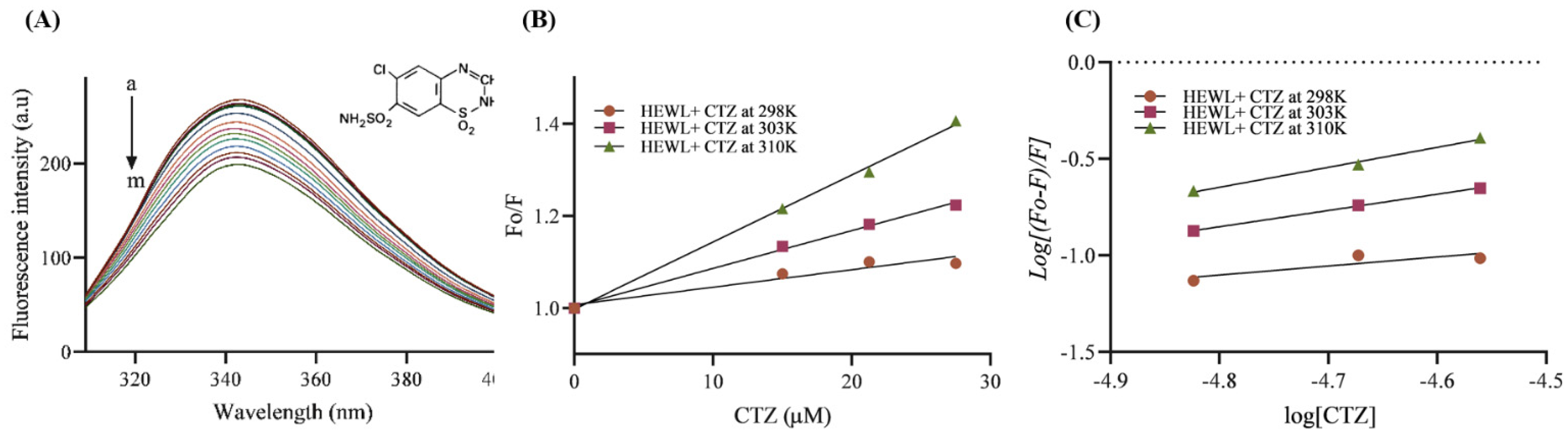
3.6. Fluorescence Quenching Analysis
3.6.1. Binding Affinity and Mechanism
3.6.2. Thermodynamic Parameters
3.7. Microscopy Imaging
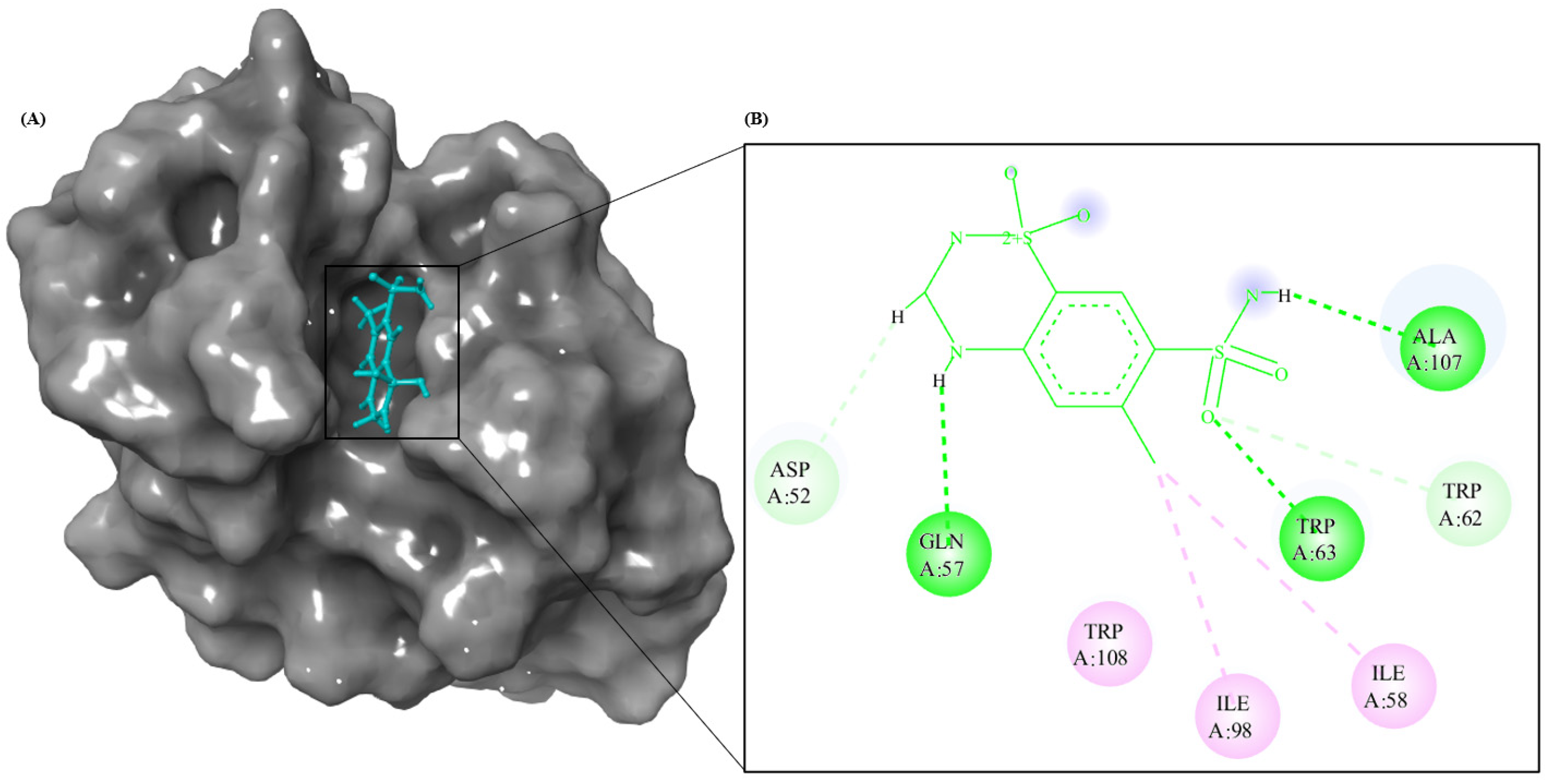
3.8. Molecular Modeling Studies
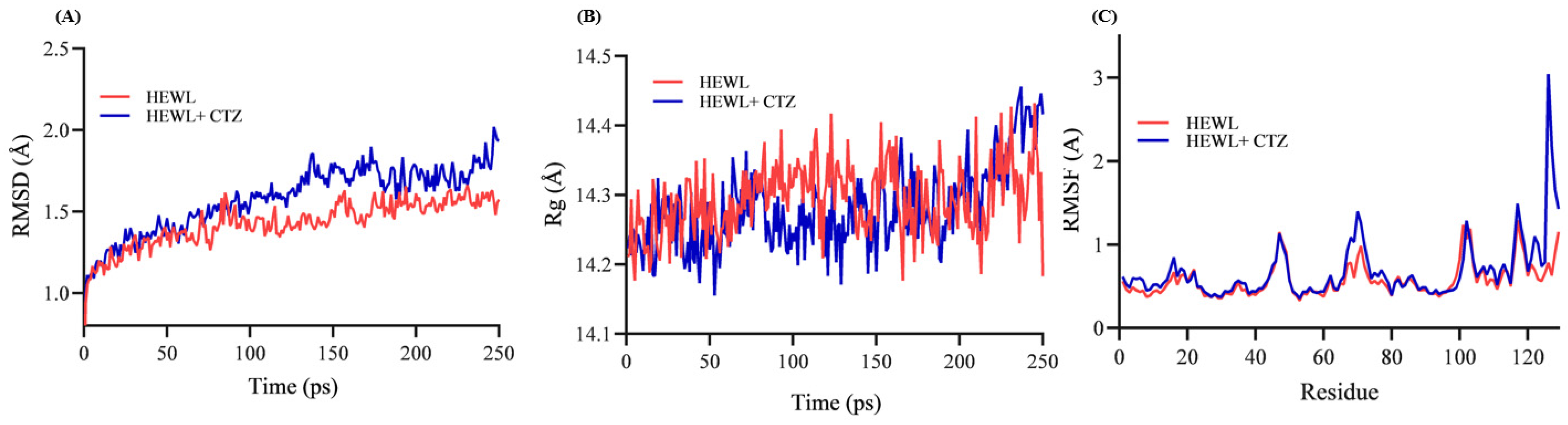
3.9. Structural Stability of HEWL–CTZ Complex
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chiti, F.; Dobson, C.M. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress Over the Last Decade. Annu. Rev. Biochem. 2017, 86, 27–68. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Siddiqi, M.K.; Majid, N.; Masroor, A.; Ali, S.M.; Khan, R.H. Unravelling the inhibitory and cytoprotective potential of diuretics towards amyloid fibrillation. Int. J. Biol. Macromol. 2019, 150, 1258–1271. [Google Scholar] [CrossRef] [PubMed]
- Tipping, K.W.; Karamanos, T.K.; Jakhria, T.; Iadanza, M.G.; Goodchild, S.C.; Tuma, R.; Ranson, N.A.; Hewitt, E.W.; Radford, S.E. pH-induced molecular shedding drives the formation of amyloid fibril-derived oligomers. Proc. Natl. Acad. Sci. USA 2015, 112, 5691–5696. [Google Scholar] [CrossRef] [PubMed]
- Moura, A.; Nocerino, P.; Gilbertson, J.A.; Rendell, N.B.; Mangione, P.P.; Verona, G.; Rowczenio, D.; Gillmore, J.D.; Taylor, G.W.; Bellotti, V.; et al. Lysozyme amyloid: Evidence for the W64R variant by proteomics in the absence of the wild type protein. Amyloid 2020, 27, 206–207. [Google Scholar] [CrossRef]
- Chen, X.; Deng, X.; Han, X.; Liang, Y.; Meng, Z.; Liu, R.; Su, W.; Zhu, H.; Fu, T. Inhibition of Lysozyme Amyloid Fibrillation by Silybin Diastereoisomers: The Effects of Stereochemistry. ACS Omega 2021, 6, 3307–3318. [Google Scholar] [CrossRef]
- Gupta, N.; Kaur, H.; Wajid, S. Renal amyloidosis: An update on diagnosis and pathogenesis. Protoplasma 2020, 257, 1259–1276. [Google Scholar] [CrossRef]
- Benyamine, A.; Bernard-Guervilly, F.; Tummino, C.; Macagno, N.; Daniel, L.; Valleix, S.; Granel, B. Hereditary lysozyme amyloidosis with sicca syndrome, digestive, arterial, and tracheobronchial involvement: Case-based review. Clin. Rheumatol. 2017, 36, 2623–2628. [Google Scholar] [CrossRef]
- Banerjee, S. Modification with N-benzylisatin restricts stress-induced aggregation of hen egg white lysozyme: Anti-amyloidogenic property of isatin derivative with possible clinical implications. Int. J. Biol. Macromol. 2021, 187, 341–349. [Google Scholar] [CrossRef]
- Trexler, A.; Nilsson, M. The Formation of Amyloid Fibrils from Proteins in the Lysozyme Family. Curr. Protein Pept. Sci. 2007, 8, 537–557. [Google Scholar] [CrossRef]
- Ghosh, S.; Pandey, N.K.; Banerjee, P.; Chaudhury, K.; Nagy, N.V.; Dasgupta, S. Copper(II) directs formation of toxic amorphous aggregates resulting in inhibition of hen egg white lysozyme fibrillation under alkaline salt-mediated conditions. J. Biomol. Struct. Dyn. 2015, 33, 991–1007. [Google Scholar] [CrossRef]
- Swaminathan, R.; Ravi, V.K.; Kumar, S.; Kumar, M.V.S.; Chandra, N. Lysozyme: A model protein for amyloid research. Adv. Protein Chem. Struct. Biol. 2011, 84, 63–111. [Google Scholar] [PubMed]
- Duarte, J.D.; Cooper-DeHoff, R.M. Mechanisms for blood pressure lowering and metabolic effects of thiazide and thiazide-like diuretics. Expert Rev. Cardiovasc. Ther. 2010, 8, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S. Antihypertensive drugs. Pharmacol. Res. 2017, 124, 116–125. [Google Scholar] [CrossRef]
- Launer, L.J.; Ross, G.W.; Petrovitch, H.; Masaki, K.; Foley, D.; White, L.R.; Havlik, R.J. Midlife blood pressure and dementia: The Honolulu-Asia aging study. Neurobiol. Aging 2000, 21, 49–55. [Google Scholar] [CrossRef]
- Chou, C.-L.; Yeh, H.-I. The Role of the Renin-Angiotensin System in Amyloid Metabolism of Alzheimer’s Disease. Acta Cardiol. Sin. 2014, 30, 114–118. [Google Scholar] [PubMed]
- Hajjar, I.; Rodgers, K. Do angiotensin receptor blockers prevent Alzheimer’s disease? Curr. Opin. Cardiol. 2013, 28, 417–425. [Google Scholar] [CrossRef] [PubMed]
- AbdAlla, S.; Langer, A.; Fu, X.; Quitterer, U. ACE inhibition with captopril retards the development of signs of neurodegeneration in an animal model of Alzheimer’s disease. Int. J. Mol. Sci. 2013, 14, 16917–16942. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, J.; Ho, L.; Ono, K.; Teplow, D.B.; Pasinetti, G.M. Identification of Antihypertensive Drugs Which Inhibit Amyloid-β Protein Oligomerization. J. Alzheimer’s Dis. 2009, 16, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Barthold, D.; Joyce, G.; Wharton, W.; Kehoe, P.; Zissimopoulos, J. The association of multiple anti-hypertensive medication classes with Alzheimer’s disease incidence across sex, race, and ethnicity. PLoS ONE 2018, 13, e0206705. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, Z.; Lin, E.; Zhao, W.; Qian, X.; Freire, D.; Bilski, A.E.; Cheng, A.; Vempati, P.; Ho, L.; et al. Unintended Effects of Cardiovascular Drugs on the Pathogenesis of Alzheimer’s Disease. PLoS ONE 2013, 8, e65232. [Google Scholar] [CrossRef]
- Rosenberg, P.; Mielke, M.; Tschanz, J.; Cook, L.; Corcoran, C.; Hayden, K.; Norton, M.; Rabins, P.; Green, R.; Welsh-Bohmer, K.; et al. Effects of Cardiovascular Medications on Rate of Functional Decline in Alzheimer Disease. Am. J. Geriatr. Psychiatry 2008, 16, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Yasar, S.; Lin, F.; Fried, L.P.; Kawas, C.H.; Sink, K.M.; DeKosky, S.T.; Carlson, M.C.; Ginkgo Evaluation of Memory (GEM). Study Investigators Diuretic use is associated with better learning and memory in older adults in the Ginkgo Evaluation of Memory study. Alzheimer’s Dement. 2012, 8, 188–195. [Google Scholar] [CrossRef]
- Chaturvedi, S.K.; Zaidi, N.; Alam, P.; Khan, J.M.; Qadeer, A.; Siddique, I.A.; Asmat, S.; Zaidi, Y.; Khan, R.H. Unraveling Comparative Anti-Amyloidogenic Behavior of Pyrazinamide and D-Cycloserine: A Mechanistic Biophysical Insight. PLoS ONE 2015, 10, e0136528. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.R. Techniques to study amyloid fibril formation in vitro. Methods 2004, 34, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Bruggink, K.A.; Müller, M.; Kuiperij, H.B.; Verbeek, M.M. Methods for Analysis of Amyloid-β Aggregates. J. Alzheimer’s Dis. 2012, 28, 735–758. [Google Scholar] [CrossRef]
- Siposova, K.; Sedlak, E.; Kozar, T.; Nemergut, M.; Musatov, A. Dual effect of non-ionic detergent Triton X-100 on insulin amyloid formation. Colloids Surf. B Biointerfaces 2018, 173, 709–718. [Google Scholar] [CrossRef]
- So, M.; Kimura, Y.; Yamaguchi, K.; Sugiki, T.; Fujiwara, T.; Aguirre, C.; Ikenaka, K.; Mochizuki, H.; Kawata, Y.; Goto, Y. Polyphenol-solubility alters amyloid fibril formation of α-synuclein. Protein Sci. 2021, 30, 1701–1713. [Google Scholar] [CrossRef]
- Siddiquee, M.A.; ud din Parray, M.; Mehdi, S.H.; Alzahrani, K.A.; Alshehri, A.A.; Malik, M.A.; Patel, R. Green synthesis of silver nanoparticles from Delonix regia leaf extracts: In-vitro cytotoxicity and interaction studies with bovine serum albumin. Mater. Chem. Phys. 2020, 242, 122493. [Google Scholar] [CrossRef]
- Zaman, M.; Ehtram, A.; Chaturvedi, S.K.; Nusrat, S.; Khan, R.H. Amyloidogenic behavior of different intermediate state of stem bromelain: A biophysical insight. Int. J. Biol. Macromol. 2016, 91, 477–485. [Google Scholar] [CrossRef]
- Patel, P.; Parmar, K.; Patel, D.; Kumar, S.; Trivedi, M.; Das, M. Inhibition of amyloid fibril formation of lysozyme by ascorbic acid and a probable mechanism of action. Int. J. Biol. Macromol. 2018, 114, 666–678. [Google Scholar] [CrossRef]
- Zaman, M.; Chaturvedi, S.K.; Zaidi, N.; Qadeer, A.; Chandel, T.I.; Nusrat, S.; Alam, P.; Khan, R.H. DNA induced aggregation of stem bromelain; a mechanistic insight. RSC Adv. 2016, 6, 37591–37599. [Google Scholar] [CrossRef]
- Liu, F.; Wang, Y.; Sang, J.; Wei, W.; Zhao, W.; Chen, B.; Zhao, F.; Jia, L.; Lu, F. Brazilin Inhibits α-Synuclein Fibrillogenesis, Disrupts Mature Fibrils, and Protects against Amyloid-Induced Cytotoxicity. J. Agric. Food Chem. 2019, 67, 11769–11777. [Google Scholar] [CrossRef] [PubMed]
- Gasymov, O.K.; Glasgow, B.J. ANS fluorescence: Potential to augment the identification of the external binding sites of proteins. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2007, 1774, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Nusrat, S.; Masroor, A.; Zaman, M.; Siddiqi, M.K.; Ajmal, M.R.; Zaidi, N.; Abdelhameed, A.S.; Khan, R.H. Interaction of catecholamine precursor l-Dopa with lysozyme: A biophysical insight. Int. J. Biol. Macromol. 2018, 109, 1132–1139. [Google Scholar] [CrossRef]
- Manzo, G.; Scorciapino, M.A.; Wadhwani, P.; Bürck, J.; Montaldo, N.P.; Pintus, M.; Sanna, R.; Casu, M.; Giuliani, A.; Pirri, G.; et al. Enhanced Amphiphilic Profile of a Short β-Stranded Peptide Improves Its Antimicrobial Activity. PLoS ONE 2015, 10, e0116379. [Google Scholar] [CrossRef]
- Anand, U.; Jash, C.; Boddepalli, R.K.; Shrivastava, A.; Mukherjee, S. Exploring the Mechanism of Fluorescence Quenching in Proteins Induced by Tetracycline. J. Phys. Chem. B 2011, 115, 6312–6320. [Google Scholar] [CrossRef]
- Li, D.; Ji, B.; Jin, J. Spectrophotometric studies on the binding of Vitamin C to lysozyme and bovine liver catalase. J. Lumin. 2008, 128, 1399–1406. [Google Scholar] [CrossRef]
- Gálvez-Iriqui, A.C.; Plascencia-Jatomea, M.; Bautista-Baños, S. Lysozymes: Characteristics, mechanism of action and technological applications on the control of pathogenic microorganisms. Rev. Mex. Fitopatol./Mex. J. Phytopathol. 2020, 38, 360–383. [Google Scholar] [CrossRef]
- Patel, P.; Parmar, K.; Das, M. Inhibition of insulin amyloid fibrillation by Morin hydrate. Int. J. Biol. Macromol. 2018, 108, 225–239. [Google Scholar] [CrossRef]
- Narang, S.S.; Shuaib, S.; Goyal, B. Molecular insights into the inhibitory mechanism of rifamycin SV against β2–microglobulin aggregation: A molecular dynamics simulation study. Int. J. Biol. Macromol. 2017, 102, 1025–1034. [Google Scholar] [CrossRef]
- Kelutur, F.J.; Mustarichie, R. Molecular docking of the potential compound from cocoa shells (Theobroma cacao L.) against androgen receptor as anti-alopecia. J. Glob. Pharma Technol. 2020, 12, 52–60. [Google Scholar]
- Julie, M.; Prabhu, T.; ASİF, F.B.; Muthu, S. In silico drug evaluation and drug research of bioactive molecule methyl 4-bromo-2-fluorobenzoate. Ank. Univ. Eczaci. Fak. Derg. 2021, 45, 10. [Google Scholar] [CrossRef]
- Akaho, E. Epigenetic Drugs, and Their Virtual Screening Study Retrieved from ZINC Database Along with an AutoDock Study of the Best Inhibitor. Curr. Asp. Pharm. Res. Dev. 2022, 7, 29–44. [Google Scholar] [CrossRef]
- Chaudhary, A.P.; Vispute, N.H.; Shukla, V.K.; Ahmad, B. A comparative study of fibrillation kinetics of two homologous proteins under identical solution condition. Biochimie 2017, 132, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Y.; Lu, T.; Li, F. The effects of organic solvents on the membrane-induced fibrillation of human islet amyloid polypeptide and on the inhibition of the fibrillation. Biochim. Biophys. Acta (BBA)—Biomembr. 2014, 1838, 3162–3170. [Google Scholar] [CrossRef]
- Odigboegwu, O.; Pan, L.J.; Chatterjee, P. Use of Antihypertensive Drugs During Preeclampsia. Front. Cardiovasc. Med. 2018, 5, 50. [Google Scholar] [CrossRef]
- Zhan, M.; Guo, M.; Jiang, Y.; Wang, X. Characterization of the Interaction between Gallic Acid and Lysozyme by Molecular Dynamics Simulation and Optical Spectroscopy. Int. J. Mol. Sci. 2015, 16, 14786–14807. [Google Scholar] [CrossRef]
- Ganesan, A.; Coote, M.L.; Barakat, K. Molecular dynamics-driven drug discovery: Leaping forward with confidence. Drug Discov. Today 2017, 22, 249–269. [Google Scholar] [CrossRef]
- Shuaib, S.; Goyal, B. Scrutiny of the mechanism of small molecule inhibitor preventing conformational transition of amyloid-β42 monomer: Insights from molecular dynamics simulations. J. Biomol. Struct. Dyn. 2017, 36, 663–678. [Google Scholar] [CrossRef]
- Halgren, T.A. Identifying and Characterizing Binding Sites and Assessing Druggability. J. Chem. Inf. Model. 2009, 49, 377–389. [Google Scholar] [CrossRef]
- Chiti, F.; Dobson, C.M. Protein Misfolding, Functional Amyloid, and Human Disease. Annu. Rev. Biochem. 2006, 75, 333–366. [Google Scholar] [CrossRef] [Green Version]
- Chun, B.J.; Choi, J.I.; Jang, S.S. Molecular dynamics simulation study of sodium dodecyl sulfate micelle: Water penetration and sodium dodecyl sulfate dissociation. Colloids Surf. A Physicochem. Eng. Asp. 2015, 474, 36–43. [Google Scholar] [CrossRef]
- Stevens, M.J.; Hoh, J.H.; Woolf, T.B. Insights into the Molecular Mechanism of Membrane Fusion from Simulation: Evidence for the Association of Splayed Tails. Phys. Rev. Lett. 2003, 91, 188102. [Google Scholar] [CrossRef] [PubMed]
- Hashemzadeh, H.; Javadi, H.; Darvishi, M.H. Study of Structural stability and formation mechanisms in DSPC and DPSM liposomes: A coarse-grained molecular dynamics simulation. Sci. Rep. 2020, 10, 1837. [Google Scholar] [CrossRef] [PubMed]
- Sawada, M.; Yamaguchi, K.; Hirano, M.; Noji, M.; So, M.; Otzen, D.E.; Kawata, Y.; Goto, Y. Amyloid Formation of α-Synuclein Based on the Solubility- and Supersaturation-Dependent Mechanism. Langmuir 2020, 36, 4671–4681. [Google Scholar] [CrossRef]
- Lee, D.; Kim, S.M.; Kim, H.Y.; Kim, Y. Fluorescence Chemicals To Detect Insoluble and Soluble Amyloid-β Aggregates. ACS Chem. Neurosci. 2019, 10, 2647–2657. [Google Scholar] [CrossRef]
- Kremer, J.J.; Pallitto, M.M.; Sklansky, D.J.; Murphy, R.M. Correlation of β-Amyloid Aggregate Size and Hydrophobicity with Decreased Bilayer Fluidity of Model Membranes. Biochemistry 2000, 39, 10309–10318. [Google Scholar] [CrossRef] [PubMed]
- Gao, W. Inhibition behavior of Sennoside A and Sennoside C on amyloid fibrillation of human lysozyme and its possible mechanism. Int. J. Biol. Macromol. 2021, 178, 424–433. [Google Scholar] [CrossRef]
- Zaidi, F.K.; Bhat, R. Two polyphenols with diverse mechanisms towards amyloidosis: Differential modulation of the fibrillation pathway of human lysozyme by curcumin and EGCG. J. Biomol. Struct. Dyn. 2020, 40, 4593–4611. [Google Scholar] [CrossRef] [PubMed]
- Brudar, S.; Hribar-Lee, B. The Role of Buffers in Wild-Type HEWL Amyloid Fibril Formation Mechanism. Biomolecules 2019, 9, 65. [Google Scholar] [CrossRef]
- Chaari, A.; Fahy, C.; Chevillot-Biraud, A.; Rholam, M. Insights into Kinetics of Agitation-Induced Aggregation of Hen Lysozyme under Heat and Acidic Conditions from Various Spectroscopic Methods. PLoS ONE 2015, 10, e0142095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Li, Y.; Hao, W.; Hu, X.; Ma, G. Parallel β-Sheet Fibril and Antiparallel β-Sheet Oligomer: New Insights into Amyloid Formation of Hen Egg White Lysozyme under Heat and Acidic Condition from FTIR Spectroscopy. J. Phys. Chem. B 2013, 117, 4003–4013. [Google Scholar] [CrossRef] [PubMed]
- Jolles, P. Lysozymes: Model Enzymes in Biochemistry and Biology; Experientia Supplementum; Birkhäuser: Basel, Switzerland, 1996; Volume 75. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
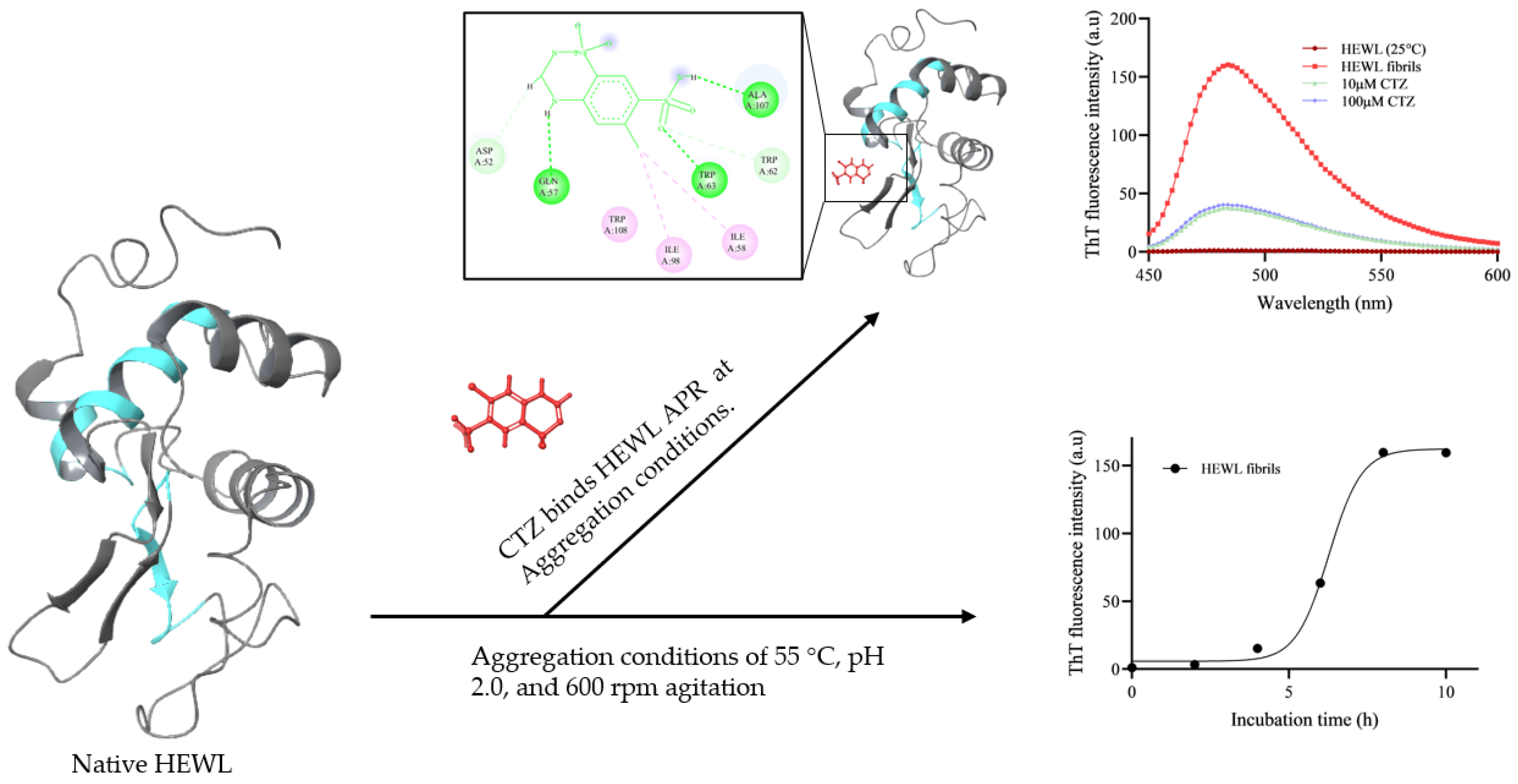{kind=link}
| T(K) | Ksv (L mol−1) | Kq (L mol−1 s−1) | Kb (M−1) | ΔH° (Kcal mol−1) | ΔS° (Kcal mol−1 K−1) | ΔG° (Kcal mol−1) |
|---|---|---|---|---|---|---|
| 298.15 | 3.80 × 109 ± 0.05 | 3.80 × 108 ± 0.05 | 1.51 × 108 ± 0.07 | 36.41 | 0.12 | −0.27 |
| 303.15 | 8.20 × 109 ± 0.0 | 8.20 × 108 ± 0.09 | 4.62 × 108 ± 0.11 | −0.88 | ||
| 310.15 | 14.56 × 109 ± 0.17 | 14.56 × 108 ± 0.17 | 16.46 × 108 ± 0.13 | −1.74 |
| Amino Acid Residues | Interactions Involved | Binding Energy (kcal mol−1) |
|---|---|---|
| ASP52 | Hydrogen Bonding | −6.58 |
| GLN57 | ||
| TRP62 | ||
| TRP63 | ||
| ALA107 | ||
| ILE58 | Hydrophobic Interactions | |
| ILE98 | ||
| TRP108 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altwaijry, N.; Almutairi, G.S.; Khan, M.S.; Shaik, G.M.; Alokail, M.S. Effect of Antihypertensive Drug (Chlorothiazide) on Fibrillation of Lysozyme: A Combined Spectroscopy, Microscopy, and Computational Study. Int. J. Mol. Sci. 2023, 24, 3112. https://doi.org/10.3390/ijms24043112
Altwaijry N, Almutairi GS, Khan MS, Shaik GM, Alokail MS. Effect of Antihypertensive Drug (Chlorothiazide) on Fibrillation of Lysozyme: A Combined Spectroscopy, Microscopy, and Computational Study. International Journal of Molecular Sciences. 2023; 24(4):3112. https://doi.org/10.3390/ijms24043112
Chicago/Turabian StyleAltwaijry, Nojood, Ghaliah S. Almutairi, Mohd Shahnawaz Khan, Gouse M. Shaik, and Majed S. Alokail. 2023. "Effect of Antihypertensive Drug (Chlorothiazide) on Fibrillation of Lysozyme: A Combined Spectroscopy, Microscopy, and Computational Study" International Journal of Molecular Sciences 24, no. 4: 3112. https://doi.org/10.3390/ijms24043112
APA StyleAltwaijry, N., Almutairi, G. S., Khan, M. S., Shaik, G. M., & Alokail, M. S. (2023). Effect of Antihypertensive Drug (Chlorothiazide) on Fibrillation of Lysozyme: A Combined Spectroscopy, Microscopy, and Computational Study. International Journal of Molecular Sciences, 24(4), 3112. https://doi.org/10.3390/ijms24043112