
Multifaceted Nature of DNA Polymerase θ


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Structure and Domain Organization of Pol θ
3. Pol θ as a Central Player in TMEJ
3.1. Discovery of TMEJ
3.2. Mechanism and Regulation of TMEJ in Cells
4. Pol θ Provides Genome (In)Stability
4.1. Templated Insertions (TINs) and Deletions—A Unique “Footprint” of Pol θ
4.2. Other Pol θ-Associated Mutations
4.3. Role of TMEJ in Genome Stability
5. Beyond TMEJ: DNA Translesion Synthesis
5.1. TLS Opposite Abasic Sites
5.2. TLS Opposite Oxidative DNA Lesions
5.3. TLS and Other Lesions
6. Beyond TMEJ: Base Excision Repair
7. Beyond TMEJ: RNA-Templated DNA Repair?
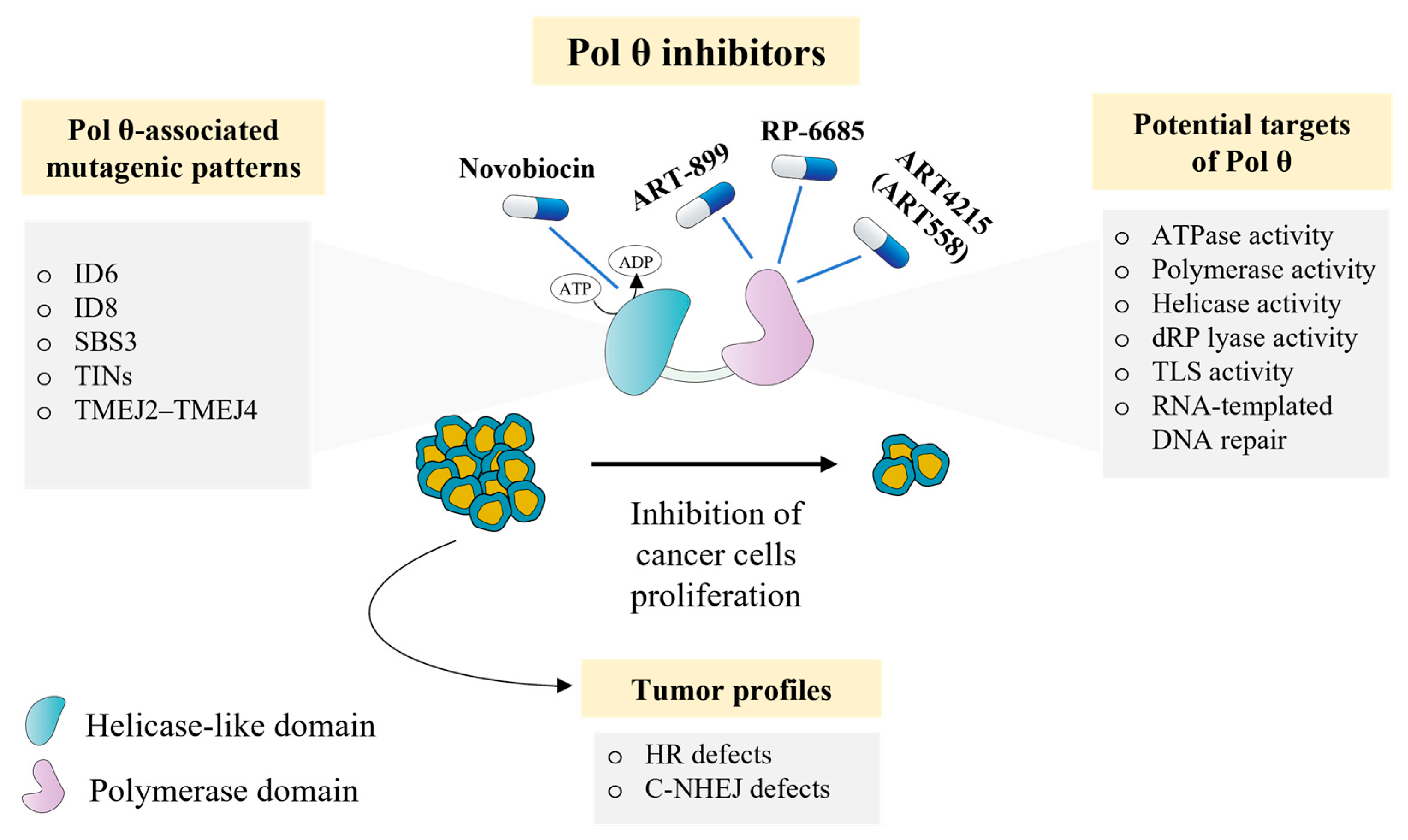
8. How Attractive Is Pol θ as a Target in Cancer Therapy?
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, W.; Gao, Y. Translesion and Repair DNA Polymerases: Diverse Structure and Mechanism. Annu. Rev. Biochem. 2018, 87, 239–261. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.V.; Mazina, O.M.; Leonhardt, E.A.; Case, R.B.; Boyd, J.B.; Burtis, K.C. Molecular Cloning of Drosophila Mus308, a Gene Involved in DNA Cross-Link Repair with Homology to Prokaryotic DNA Polymerase I Genes. Mol. Cell. Biol. 1996, 16, 5764–5771. [Google Scholar] [CrossRef] [PubMed]
- Shima, N.; Munroe, R.J.; Schimenti, J.C. The Mouse Genomic Instability Mutation Chaos1 Is an Allele of Polq That Exhibits Genetic Interaction with Atm. Mol. Cell. Biol. 2004, 24, 10381–10389. [Google Scholar] [CrossRef] [PubMed]
- Schrempf, A.; Slyskova, J.; Loizou, J.I. Targeting the DNA Repair Enzyme Polymerase θ in Cancer Therapy. Trends Cancer 2020, 7, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Wood, R.D.; Doublié, S. Genome Protection by DNA Polymerase θ. Annu. Rev. Genet. 2022, 56, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Zahn, K.E.; Jensen, R.B. Polymerase θ Coordinates Multiple Intrinsic Enzymatic Activities during DNA Repair. Genes 2021, 12, 1310. [Google Scholar] [CrossRef]
- Ramsden, D.A.; Carvajal-Garcia, J.; Gupta, G.P. Mechanism, Cellular Functions and Cancer Roles of Polymerase-Theta-Mediated DNA End Joining. Nat. Rev. Mol. Cell Biol. 2022, 23, 125–140. [Google Scholar] [CrossRef]
- Brambati, A.; Barry, R.M.; Sfeir, A. DNA Polymerase Theta (Polθ)—An Error-Prone Polymerase Necessary for Genome Stability. Curr. Opin. Genet. Dev. 2020, 60, 119–126. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Wood, R.D. DNA Polymerase POLQ and Cellular Defense against DNA Damage. DNA Repair (Amst) 2013, 12, 1–9. [Google Scholar] [CrossRef]
- Newman, J.A.; Cooper, C.D.O.; Aitkenhead, H.; Gileadi, O. Structure of the Helicase Domain of DNA Polymerase Theta Reveals a Possible Role in the Microhomology-Mediated End-Joining Pathway. Structure 2015, 23, 2319–2330. [Google Scholar] [CrossRef] [Green Version]
- Zahn, K.E.; Averill, A.M.; Aller, P.; Wood, R.D.; Doublié, S. Human DNA Polymerase θ Grasps the Primer Terminus to Mediate DNA Repair. Nat. Struct. Mol. Biol. 2015, 22, 304–311. [Google Scholar] [CrossRef]
- Seki, M.; Marini, F.; Wood, R.D. POLQ (Pol θ), a DNA Polymerase and DNA-Dependent ATPase in Human Cells. Nucleic Acids Res. 2003, 31, 6117–6126. [Google Scholar] [CrossRef]
- Li, C.; Zhu, H.; Jin, S.; Maksoud, L.M.; Jain, N.; Sun, J.; Gao, Y. Structural Basis of DNA Polymerase θ Mediated DNA End Joining. Nucleic Acids Res. 2023, 51, 463–474. [Google Scholar] [CrossRef]
- Mateos-Gomez, P.A.; Kent, T.; Deng, S.K.; Mcdevitt, S.; Kashkina, E.; Hoang, T.M.; Pomerantz, R.T.; Sfeir, A. The Helicase Domain of Polθ Counteracts RPA to Promote Alt-NHEJ. Nat. Struct. Mol. Biol. 2017, 24, 1116–1123. [Google Scholar] [CrossRef]
- Ozdemir, A.Y.; Rusanov, T.; Kent, T.; Siddique, L.A.; Pomerantz, R.T. Polymerase θ-Helicase Efficiently Unwinds DNA and RNA-DNA Hybrids. J. Biol. Chem. 2018, 293, 5259–5269. [Google Scholar] [CrossRef]
- Hogg, M.; Seki, M.; Wood, R.D.; Doublié, S.; Wallace, S.S. Lesion Bypass Activity of DNA Polymerase θ (POLQ) Is an Intrinsic Property of the Pol Domain and Depends on Unique Sequence Inserts. J. Mol. Biol. 2011, 405, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Kent, T.; Chandramouly, G.; Mcdevitt, S.M.; Ozdemir, A.Y.; Pomerantz, R.T. Mechanism of Microhomology-Mediated End-Joining Promoted by Human DNA Polymerase θ. Nat. Struct. Mol. Biol. 2015, 22, 230–237. [Google Scholar] [CrossRef]
- Black, S.J.; Ozdemir, A.Y.; Kashkina, E.; Kent, T.; Rusanov, T.; Ristic, D.; Shin, Y.; Suma, A.; Hoang, T.; Chandramouly, G.; et al. Molecular Basis of Microhomology-Mediated End-Joining by Purified Full-Length Polθ. Nat. Commun. 2019, 10, 4423. [Google Scholar] [CrossRef]
- Ceccaldi, R.; Liu, J.C.; Amunugama, R.; Hajdu, I.; Primack, B.; Petalcorin, M.I.R.; O’Connor, K.W.; Konstantinopoulos, P.A.; Elledge, S.J.; Boulton, S.J.; et al. Homologous-Recombination-Deficient Tumours Are Dependent on Polθ-Mediated Repair. Nature 2015, 518, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Rothenberg, E.; Ramsden, D.A.; Lieber, M.R. The Molecular Basis and Disease Relevance of Non-Homologous DNA End Joining. Nat. Rev. Mol. Cell Biol. 2020, 21, 765–781. [Google Scholar] [PubMed]
- Goff, J.P.; Shields, D.S.; Seki, M.; Choi, S.; Epperly, M.W.; Wang, H.; Bakkenist, C.J.; Dertinger, S.D.; Dorothea, K.; Wittschieben, J.; et al. Lack of DNA Polymerase θ (POLQ) Radiosensitizes Bone Marrow Stromal Cells In Vitro and Increases Reticulocyte Micronuclei after Total-Body Irradiation. Radiat. Res. 2009, 172, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.H.; Yu, A.M.; McVey, M. Dual Roles for DNA Polymerase Theta in Alternative End-Joining Repair of Double-Strand Breaks in Drosophila. PLoS Genet. 2010, 6, e1001005. [Google Scholar] [CrossRef]
- Truong, L.N.; Li, Y.; Shi, L.Z.; Hwang, P.Y.H.; He, J.; Wang, H.; Razavian, N.; Berns, M.W.; Wu, X. Microhomology-Mediated End Joining and Homologous Recombination Share the Initial End Resection Step to Repair DNA Double-Strand Breaks in Mammalian Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 7720–7725. [Google Scholar] [CrossRef]
- Bothmer, A.; Robbiani, D.F.; Feldhahn, N.; Gazumyan, A.; Nussenzweig, A.; Nussenzweig, M.C. 53BP1 Regulates DNA Resection and the Choice between Classical and Alternative End Joining during Class Switch Recombination. J. Exp. Med. 2010, 207, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.S.; Balestrini, A.; Petrini, J.H.J. Functional Interplay of the Mre11 Nuclease and Ku in the Response to Replication-Associated DNA Damage. Mol. Cell Biol. 2011, 31, 4379–4389. [Google Scholar] [CrossRef]
- Deng, S.K.; Gibb, B.; De Almeida, M.J.; Greene, E.C.; Symington, L.S. RPA Antagonizes Microhomology-Mediated Repair of DNA Double-Strand Breaks. Nat. Struct. Mol. Biol. 2014, 21, 405–412. [Google Scholar] [CrossRef]
- Ahrabi, S.; Sarkar, S.; Pfister, S.X.; Pirovano, G.; Higgins, G.S.; Porter, A.C.G.; Humphrey, T.C. A Role for Human Homologous Recombination Factors in Suppressing Microhomology-Mediated End Joining. Nucleic Acids Res. 2016, 44, 5743–5757. [Google Scholar] [CrossRef]
- Wyatt, D.W.; Feng, W.; Conlin, M.P.; Yousefzadeh, M.J.; Roberts, S.A.; Mieczkowski, P.; Wood, R.D.; Gupta, G.P.; Ramsden, D.A. Essential Roles for Polymerase θ-Mediated End Joining in the Repair of Chromosome Breaks. Mol. Cell 2016, 63, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Lescale, C.; Babin, L.; Bedora-Faure, M.; Lenden-Hasse, H.; Baron, L.; Demangel, C.; Yelamos, J.; Brunet, E.; Deriano, L. Repair of G1 Induced DNA Double-Strand Breaks in S-G2/M by Alternative NHEJ. Nat. Commun. 2020, 11, 5239. [Google Scholar] [CrossRef] [PubMed]
- Audebert, M.; Salles, B.; Weinfeld, M.; Calsou, P. Involvement of Polynucleotide Kinase in a Poly(ADP-Ribose) Polymerase-1-Dependent DNA Double-Strand Breaks Rejoining Pathway. J. Mol. Biol. 2006, 356, 257–265. [Google Scholar] [CrossRef]
- Mateos-Gomez, P.A.; Gong, F.; Nair, N.; Miller, K.M.; Lazzerini-Denchi, E.; Sfeir, A. Mammalian Polymerase θ Promotes Alternative NHEJ and Suppresses Recombination. Nature 2015, 518, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Luedeman, M.E.; Stroik, S.; Feng, W.; Luthman, A.J.; Gupta, G.P.; Ramsden, D.A. Poly(ADP) Ribose Polymerase Promotes DNA Polymerase Theta-Mediated End Joining by Activation of End Resection. Nat. Commun. 2022, 13, 4547. [Google Scholar] [CrossRef]
- Schaub, J.M.; Soniat, M.M.; Finkelstein, I.J. Polymerase Theta-Helicase Promotes End Joining by Stripping Single-Stranded DNA-Binding Proteins and Bridging DNA Ends. Nucleic Acids Res. 2022, 50, 3911–3921. [Google Scholar] [CrossRef]
- Carvajal-Garcia, J.; Cho, J.E.; Carvajal-Garcia, P.; Feng, W.; Wood, R.D.; Sekelsky, J.; Gupta, G.P.; Roberts, S.A.; Ramsden, D.A. Mechanistic Basis for Microhomology Identification and Genome Scarring by Polymerase Theta. Proc. Natl. Acad. Sci. USA 2020, 117, 8476–8485. [Google Scholar] [CrossRef]
- Zahn, K.E.; Jensen, R.B.; Wood, R.D.; Doublié, S. Human DNA Polymerase θ Harbors DNA End-Trimming Activity Critical for DNA Repair. Mol. Cell 2021, 81, 1534–1547.e4. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Robinson, A.R.; Duensing, A.; van Drunen, E.; Beverloo, H.B.; Weisberg, D.B.; Hasty, P.; Hoeijmakers, J.H.J.; Niedernhofer, L.J. ERCC1-XPF Endonuclease Facilitates DNA Double-Strand Break Repair. Mol. Cell. Biol. 2008, 28, 5082–5092. [Google Scholar] [CrossRef] [PubMed]
- Arana, M.E.; Seki, M.; Wood, R.D.; Rogozin, I.B.; Kunkel, T.A. Low-Fidelity DNA Synthesis by Human DNA Polymerase Theta. Nucleic Acids Res. 2008, 36, 3847–3856. [Google Scholar] [CrossRef] [PubMed]
- Masutani, C.; Rika Kusumoto, S.I. and F.H. Mechanisms of Accurate Translesion Synthesis by Human DNA Polymerase η. EMBO J. 2000, 19, 3100–3109. [Google Scholar] [CrossRef] [PubMed]
- Haracska, L.; Johnson, R.E.; Unk, I.; Phillips, B.B.; Hurwitz, J.; Prakash, L.; Prakash, S. Targeting of Human DNA Polymerase ι to the Replication Machinery via Interaction with PCNA. Proc. Natl. Acad. Sci. USA 2001, 98, 14256–14261. [Google Scholar] [CrossRef] [PubMed]
- van Schendel, R.; van Heteren, J.; Welten, R.; Tijsterman, M. Genomic Scars Generated by Polymerase Theta Reveal the Versatile Mechanism of Alternative End-Joining. PLoS Genet. 2016, 12, e1006368. [Google Scholar] [CrossRef] [Green Version]
- Meyer, D.; Fu, B.X.H.; Heyer, W.D. DNA Polymerases δ and λ Cooperate in Repairing Double-Strand Breaks by Microhomology-Mediated End-Joining in Saccharomyces Cerevisiae. Proc. Natl. Acad. Sci. USA 2015, 112, E6907–E6916. [Google Scholar] [CrossRef]
- Layer, J.V.; Debaize, L.; van Scoyk, A.; House, N.C.; Brown, A.J.; Liu, Y.; Stevenson, K.E.; Hemann, M.; Roberts, S.A.; Price, B.D.; et al. Polymerase δ Promotes Chromosomal Rearrangements and Imprecise Double-Strand Break Repair. Proc. Natl. Acad. Sci. USA 2020, 117, 27566–27577. [Google Scholar] [CrossRef]
- Ray, S.; Breuer, G.; DeVeaux, M.; Zelterman, D.; Bindra, R.; Sweasy, J.B. DNA Polymerase Beta Participates in DNA End-Joining. Nucleic Acids Res. 2018, 46, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Mengwasser, K.E.; Adeyemi, R.O.; Leng, Y.; Choi, M.Y.; Clairmont, C.; D’Andrea, A.D.; Elledge, S.J. Genetic Screens Reveal FEN1 and APEX2 as BRCA2 Synthetic Lethal Targets. Mol. Cell 2019, 73, 885–899.e6. [Google Scholar] [CrossRef]
- Howard, S.M.; Yanez, D.A.; Stark, J.M. DNA Damage Response Factors from Diverse Pathways, Including DNA Crosslink Repair, Mediate Alternative End Joining. PLoS Genet. 2015, 11, e1004943. [Google Scholar] [CrossRef] [PubMed]
- Schimmel, J.; van Schendel, R.; den Dunnen, J.T.; Tijsterman, M. Templated Insertions: A Smoking Gun for Polymerase Theta-Mediated End Joining. Trends Genet. 2019, 35, 632–644. [Google Scholar] [CrossRef]
- Schimmel, J.; Kool, H.; Schendel, R.; Tijsterman, M. Mutational Signatures of Non-homologous and Polymerase Theta-mediated End-joining in Embryonic Stem Cells. EMBO J. 2017, 36, 3634–3649. [Google Scholar] [CrossRef] [PubMed]
- Khodaverdian, V.Y.; Hanscom, T.; Yu, A.M.; Yu, T.L.; Mak, V.; Brown, A.J.; Roberts, S.A.; McVey, M. Secondary Structure Forming Sequences Drive SD-MMEJ Repair of DNA Double-Strand Breaks. Nucleic Acids Res. 2017, 45, 12848–12861. [Google Scholar] [CrossRef]
- Feldman, T.; Bercovich, A.; Moskovitz, Y.; Chapal-Ilani, N.; Mitchell, A.; Medeiros, J.J.F.; Biezuner, T.; Kaushansky, N.; Minden, M.D.; Gupta, V.; et al. Recurrent Deletions in Clonal Hematopoiesis Are Driven by Microhomology-Mediated End Joining. Nat. Commun. 2021, 12, 2455. [Google Scholar] [CrossRef]
- Maga, G.; Shevelev, I.; Ramadan, K.; Spadari, S.; Hübscher, U. DNA Polymerase θ Purified from Human Cells Is a High-Fidelity Enzyme. J. Mol. Biol. 2002, 319, 359–369. [Google Scholar] [CrossRef]
- Seki, M.; Masutani, C.; Yang, L.W.; Schuffert, A.; Iwai, S.; Bahar, I.; Wood, R.D. High-Efficiency Bypass of DNA Damage by Human DNA Polymerase Q. EMBO J. 2004, 23, 4484–4494. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Gao, Y.; Yang, W. How a Homolog of High-Fidelity Replicases Conducts Mutagenic DNA Synthesis. Nat. Struct. Mol. Biol. 2015, 22, 298–303. [Google Scholar] [CrossRef]
- Zelensky, A.N.; Schimmel, J.; Kool, H.; Kanaar, R.; Tijsterman, M. Inactivation of Pol θ and C-NHEJ Eliminates off-Target Integration of Exogenous DNA. Nat. Commun. 2017, 8, 66. [Google Scholar] [CrossRef]
- Saito, S.; Maeda, R.; Adachi, N. Dual Loss of Human POLQ and LIG4 Abolishes Random Integration. Nat. Commun. 2017, 8, 16112. [Google Scholar] [CrossRef] [PubMed]
- Roerink, S.F.; Schendel, R.; Tijsterman, M. Polymerase Theta-Mediated End Joining of Replication-Associated DNA Breaks in C. Elegans. Genome Res. 2014, 24, 954–962. [Google Scholar] [CrossRef]
- van Schendel, R.; Roerink, S.F.; Portegijs, V.; van den Heuvel, S.; Tijsterman, M. Polymerase θ Is a Key Driver of Genome Evolution and of CRISPR/Cas9-Mediated Mutagenesis. Nat. Commun. 2015, 6, 7394. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Simpson, D.A.; Carvajal-Garcia, J.; Price, B.A.; Kumar, R.J.; Mose, L.E.; Wood, R.D.; Rashid, N.; Purvis, J.E.; Parker, J.S.; et al. Genetic Determinants of Cellular Addiction to DNA Polymerase Theta. Nat. Commun. 2019, 10, 4286. [Google Scholar] [CrossRef] [PubMed]
- Zatreanu, D.; Robinson, H.M.R.; Alkhatib, O.; Boursier, M.; Finch, H.; Geo, L.; Grande, D.; Grinkevich, V.; Heald, R.A.; Langdon, S.; et al. Polθ Inhibitors Elicit BRCA-Gene Synthetic Lethality and Target PARP Inhibitor Resistance. Nat. Commun. 2021, 12, 3636. [Google Scholar] [CrossRef] [PubMed]
- Mann, A.; Ramirez-Otero, M.A.; De Antoni, A.; Hanthi, Y.W.; Sannino, V.; Baldi, G.; Falbo, L.; Schrempf, A.; Bernardo, S.; Loizou, J.; et al. POLθ Prevents MRE11-NBS1-CtIP-Dependent Fork Breakage in the Absence of BRCA2/RAD51 by Filling Lagging-Strand Gaps. Mol. Cell 2022, 82, 4218–4231.e8. [Google Scholar] [CrossRef]
- Carvajal-Garcia, J.; Crown, K.N.; Ramsden, D.A.; Sekelsky, J. DNA Polymerase Theta Suppresses Mitotic Crossing Over. PLoS Genet. 2021, 17, e1009267. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.; Khoo, K.J.; Zhang, Y.; Maizels, N. POLQ Suppresses Interhomolog Recombination and Loss of Heterozygosity at Targeted DNA Breaks. Proc. Natl. Acad. Sci. USA 2020, 117, 22900–22909. [Google Scholar] [CrossRef]
- Wang, Z.; Song, Y.; Li, S.; Kurian, S.; Xiang, R.; Chiba, T.; Wu, X. DNA Polymerase (POLQ) Is Important for Repair of DNA Double-Strand Breaks Caused by Fork Collapse. J. Biol. Chem. 2019, 294, 3909–3919. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H. Error-Prone Replication through UV Lesions by DNA Polymerase θ Protects against Skin Cancers. Cell 2019, 176, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Koole, W.; Van Schendel, R.; Karambelas, A.E.; Van Heteren, J.T.; Okihara, K.L.; Tijsterman, M. A Polymerase Theta-Dependent Repair Pathway Suppresses Extensive Genomic Instability at Endogenous G4 DNA Sites. Nat. Commun. 2014, 5, 3216. [Google Scholar] [CrossRef] [PubMed]
- Ignatov, A.V.; Bondarenko, K.A.; Makarova, A.V. Non-Bulky Lesions in Human DNA: The Ways of Formation, Repair, and Replication. Acta Nat. 2017, 9, 12–26. [Google Scholar] [CrossRef]
- Laverty, D.J.; Averill, A.M.; Doublié, S.; Greenberg, M.M. The A-Rule and Deletion Formation during Abasic and Oxidized Abasic Site Bypass by DNA Polymerase θ. ACS Chem. Biol. 2017, 12, 1584–1592. [Google Scholar] [CrossRef]
- Yoon, J.H.; Johnson, R.E.; Prakash, L.; Prakash, S. Genetic Evidence for Reconfiguration of DNA Polymerase θ Active Site for Error-Free Translesion Synthesis in Human Cells. J. Biol. Chem. 2020, 295, 5918–5927. [Google Scholar] [CrossRef] [PubMed]
- Yudkina, A.V.; Shilkin, E.S.; Endutkin, A.V.; Makarova, A.V.; Zharkov, D.O. Reading and Misreading 8-Oxoguanine, a Paradigmatic Ambiguous Nucleobase. Crystals 2019, 9, 269. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, X.; Wang, Z.; Dai, X.; You, C. Next-Generation Sequencing-Based Analysis of the Roles of DNA Polymerases ν and θ in the Replicative Bypass of 8-Oxo-7,8-Dihydroguanine in Human Cells. ACS Chem. Biol. 2022, 17, 2315–2319. [Google Scholar] [CrossRef]
- Laverty, D.; Greenberg, M. In Vitro Bypass of Thymidine Glycol by DNA Polymerase Theta Forms Sequence-Dependent Frameshift Mutations. Biochemistry 2017, 56, 6726–6733. [Google Scholar] [CrossRef]
- Yoon, J.H.; Choudhury, J.R.; Park, J.; Prakash, S.; Prakash, L. A Role for DNA Polymerase θ in Promoting Replication through Oxidative DNA Lesion, Thymine Glycol, in Human Cells. J. Biol. Chem. 2014, 289, 13177–13185. [Google Scholar] [CrossRef]
- Rioux, K.L.; Delaney, S. 1,N6-Ethenoadenine: From Molecular to Biological Consequences. Chem. Res. Toxicol. 2020, 33, 2688–2698. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Johnson, R.E.; Prakash, L.; Prakash, S. DNA Polymerase θ Accomplishes Translesion Synthesis Opposite 1,N6-Ethenodeoxyadenosine with a Remarkably High Fidelity in Human Cells. Genes Dev. 2019, 34, 146. [Google Scholar] [CrossRef]
- Du, H.; Wang, P.; Wu, J.; He, X.; Wang, Y.; Sung, P. The Roles of Polymerases ν and θ in Replicative Bypass of O6- And N2-Alkyl-2′-Deoxyguanosine Lesions in Human Cells. J. Biol. Chem. 2020, 295, 4556–4562. [Google Scholar] [CrossRef]
- Yoon, J.H.; Choudhury, J.R.; Park, J.; Prakash, S.; Prakash, L. Translesion Synthesis DNA Polymerases Promote Error-Free Replication through the Minor-Groove DNA Adduct 3-Deaza-3-Methyladenine. J. Biol. Chem. 2017, 292, 18682–18688. [Google Scholar] [CrossRef] [PubMed]
- Conde, J.; Yoon, J.H.; Choudhury, J.R.; Prakash, L.; Prakash, S. Genetic Control of Replication through N1-Methyladenine in Human Cells. J. Biol. Chem. 2015, 290, 29794–29800. [Google Scholar] [CrossRef] [PubMed]
- Seki Mineaki, R.W. DNA Polymerase θ (POLQ) Can Extend from Mismatches and from Bases Opposite a (6–4) Photoproduct. DNA Repair 2008, 7, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, M.; Kohzaki, M.; Nakamura, J.; Asagoshi, K.; Hou, E.; Prasad, R.; Wilson, S.H.; Tano, K.; Yasui, A.; Seki, M.; et al. Vertebrate POLQ and POLβ Cooperate in Base Excision Repair of Oxidative DNA Damage. Mol. Cell 2006, 24, 115–125. [Google Scholar] [CrossRef]
- Prasad, R.; Longley, M.J.; Sharief, F.S.; Hou, E.W.; Copeland, W.C.; Wilson, S.H. Human DNA Polymerase θ Possesses 5′-DRP Lyase Activity and Functions in Single-Nucleotide Base Excision Repair in Vitro. Nucleic Acids Res. 2009, 37, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Laverty, D.J.; Mortimer, I.P.; Greenberg, M.M. Mechanistic Insight Through Irreversible Inhibition: DNA Polymerase θ Uses a Common Active Site for Polymerase and Lyase Activities. J. Am. Chem. Soc. 2018, 140, 9034–9037. [Google Scholar] [CrossRef] [PubMed]
- Laverty, D.J.; Greenberg, M.M. Expanded Substrate Scope of DNA Polymerase Theta and DNA Polymerase Beta: Lyase Activity on 5′-Overhangs and Clustered Lesions. Biochemistry 2018, 57, 6119–6127. [Google Scholar] [CrossRef]
- Chan, K.Y.; Li, X.; Ortega, J.; Gu, L.; Li, G. DNA Polymerase θ Promotes CAG CTG Repeat Expansions in Huntington ’ s Disease via Insertion Sequences of Its Catalytic Domain. J. Biol. Chem. 2021, 297, 101144. [Google Scholar] [CrossRef] [PubMed]
- Storici, F.; Bebenek, K.; Kunkel, T.A.; Gordenin, D.A.; Resnick, M.A. RNA-Templated DNA Repair. Nature 2007, 447, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Keskin, H.; Meers, C.; Storici, F. Transcript RNA Supports Precise Repair of Its Own DNA Gene. RNA Biol. 2016, 13, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Meers, C.; Keskin, H.; Storici, F. DNA Repair by RNA: Templated, or Not Templated, That Is the Question. DNA Repair 2016, 44, 17–21. [Google Scholar] [CrossRef]
- Chandramouly, G.; Zhao, J.; McDevitt, S.; Rusanov, T.; Hoang, T.; Borisonnik, N.; Treddinick, T.; Lopezcolorado, F.W.; Kent, T.; Siddique, L.A.; et al. Polθ Reverse Transcribes RNA and Promotes RNA-Templated DNA Repair. Sci. Adv. 2021, 7, eabf1771. [Google Scholar] [CrossRef]
- Su, Y.; Ghodke, P.P.; Egli, M.; Li, L.; Wang, Y.; Peter Guengerich, F. Human DNA Polymerase η Has Reverse Transcriptase Activity in Cellular Environments. J. Biol. Chem. 2019, 294, 6073–6081. [Google Scholar] [CrossRef]
- Su, Y.; Egli, M.; Guengerich, F.P.; Sung, P. Human DNA Polymerase η Accommodates RNA for Strand Extension. J. Biol. Chem. 2017, 292, 18044–18051. [Google Scholar] [CrossRef]
- Nick McElhinny, S.A.; Watts, B.E.; Kumar, D.; Watt, D.L.; Lundström, E.B.; Burgers, P.M.J.; Johansson, E.; Chabes, A.; Kunkel, T.A. Abundant Ribonucleotide Incorporation into DNA by Yeast Replicative Polymerases. Proc. Natl. Acad. Sci. USA 2010, 107, 4949–4954. [Google Scholar] [CrossRef]
- Balachander, S.; Gombolay, A.L.; Yang, T.; Xu, P.; Newnam, G.; Keskin, H.; El-Sayed, W.M.M.; Bryksin, A.V.; Tao, S.; Bowen, N.E.; et al. Ribonucleotide Incorporation in Yeast Genomic DNA Shows Preference for Cytosine and Guanosine Preceded by Deoxyadenosine. Nat. Commun. 2020, 11, 2447. [Google Scholar] [CrossRef]
- Li, Y.; Breaker, R.R. Kinetics of RNA Degradation by Specific Base Catalysis of Transesterification Involving the 2γ-Hydroxyl Group. J. Am. Chem. Soc. 1999, 121, 5364–5372. [Google Scholar] [CrossRef]
- Kellner, V.; Luke, B. Molecular and Physiological Consequences of Faulty Eukaryotic Ribonucleotide Excision Repair. EMBO J. 2020, 39, e102309. [Google Scholar] [CrossRef] [PubMed]
- Reijns, M.A.M.; Rabe, B.; Rigby, R.E.; Mill, P.; Astell, K.R.; Lettice, L.A.; Boyle, S.; Leitch, A.; Keighren, M.; Kilanowski, F.; et al. Enzymatic Removal of Ribonucleotides from DNA Is Essential for Mammalian Genome Integrity and Development. Cell 2012, 149, 1008–1022. [Google Scholar] [CrossRef]
- Kawamura, K.; Bahar, R.; Seimiya, M.; Chiyo, M.; Wada, A.; Okada, S.; Hatano, M.; Tokuhisa, T.; Kimura, H.; Watanabe, S.; et al. DNA Polymerase θ Is Preferentially Expressed in Lymphoid Tissues and Upregulated in Human Cancers. Int. J. Cancer 2004, 109, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Brandalize, A.P.C.; Schüler-Faccini, L.; Hoffmann, J.S.; Caleffi, M.; Cazaux, C.; Ashton-Prolla, P. A DNA Repair Variant in POLQ (c.-1060A > G) Is Associated to Hereditary Breast Cancer Patients: A Case-Control Study. BMC Cancer 2014, 14, 850. [Google Scholar] [CrossRef] [PubMed]
- Smolinska, A.; Singer, K.; Golchert, J.; Smyczynska, U.; Fendler, W.; Sendler, M.; van den Brandt, J.; Singer, S.; Homuth, G.; Lerch, M.M.; et al. DNA Polymerase Theta Plays a Critical Role in Pancreatic Cancer Development and Metastasis. Cancers 2022, 14, 4077. [Google Scholar] [CrossRef]
- Prodhomme, M.K.; Péricart, S.; Pommier, R.M.; Morel, A.P.; Brunac, A.C.; Franchet, C.; Moyret-Lalle, C.; Brousset, P.; Puisieux, A.; Hoffmann, J.S.; et al. Opposite Roles for ZEB1 and TMEJ in the Regulation of Breast Cancer Genome Stability. Front. Cell Dev. Biol. 2021, 9, 727429. [Google Scholar] [CrossRef]
- Higgins, G.S.; Harris, A.L.; Prevo, R.; Helleday, T.; McKenna, W.G.; Buffa, F.M. Overexpression of POLQ Confers a Poor Prognosis in Early Breast Cancer Patients. Oncotarget 2010, 1, 175–184. [Google Scholar] [CrossRef]
- Lemée, F.; Bergoglio, V.; Fernandez-Vidal, A.; Machado-Silva, A.; Pillaire, M.J.; Bieth, A.; Gentil, C.; Baker, L.; Martin, A.L.; Leduc, C.; et al. DNA Polymerase θ Up-Regulation Is Associated with Poor Survival in Breast Cancer, Perturbs DNA Replication, and Promotes Genetic Instability. Proc. Natl. Acad. Sci. USA 2010, 107, 13390–13395. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Wang, L.; Liu, Y.; Li, M.; Zhang, Y.; Peng, W.; Deng, T.; Peng, M.L.; Jiang, J.Q.; Tang, J.; et al. Knockdown of POLQ Interferes the Development and Progression of Hepatocellular Carcinoma through Regulating Cell Proliferation, Apoptosis and Migration. Cancer Cell Int. 2021, 21, 482. [Google Scholar] [CrossRef]
- Kuei, C.H.; Lin, H.Y.; Lin, M.H.; Lee, H.H.; Lin, C.H.; Lee, W.J.; Chen, Y.L.; Lu, L.S.; Zheng, J.Q.; Hung, R.C.; et al. DNA Polymerase Theta Repression Enhances the Docetaxel Responsiveness in Metastatic Castration-Resistant Prostate Cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165954. [Google Scholar] [CrossRef] [PubMed]
- Allera-Moreau, C.; Rouquette, I.; Lepage, B.; Oumouhou, N.; Walschaerts, M.; Leconte, E.; Schilling, V.; Gordien, K.; Brouchet, L.; Delisle, M.B.; et al. DNA Replication Stress Response Involving PLK1, CDC6, POLQ, RAD51 and CLASPIN Upregulation Prognoses the Outcome of Early/Mid-Stage Non-Small Cell Lung Cancer Patients. Oncogenesis 2012, 1, e30. [Google Scholar] [CrossRef] [PubMed]
- Prodhomme, M.K.; Pommier, R.M.; Franchet, C.; Fauvet, F.; Bergoglio, V.; Brousset, P.; Morel, A.-P.; Brunac, A.-C.; Devouassoux-Shisheboran, M.; Petrilli, V.; et al. EMT Transcription Factor ZEB1 Represses the Mutagenic POLθ-Mediated End-Joining Pathway in Breast Cancers. Cancer Res. 2020, 81, 1595–1606. [Google Scholar] [CrossRef]
- Gellert, M.; O’Dea, M.H.; Itoh, T.; Tomizawa, J.I. Novobiocin and Coumermycin Inhibit DNA Supercoiling Catalyzed by DNA Gyrase. Proc. Natl. Acad. Sci. USA 1976, 73, 4474–4478. [Google Scholar] [CrossRef]
- Zhou, J.; Gelot, C.; Pantelidou, C.; Li, A.; Yücel, H.; Davis, R.E.; Färkkilä, A.; Kochupurakkal, B.; Syed, A.; Shapiro, G.I.; et al. A First-in-Class Polymerase Theta Inhibitor Selectively Targets Homologous-Recombination-Deficient Tumors. Nat. Cancer 2021, 2, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Bubenik, M.; Mader, P.; Mochirian, P.; Clark, J.; Truchon, J.; Perryman, A.L.; Pau, V.; Kurinov, I.; Zahn, K.E.; Leclaire, M.; et al. Identification of RP-6685, an Orally Bioavailable Compound That Inhibits the DNA Polymerase Activity of Polθ. J. Med. Chem. 2022, 65, 13198–13215. [Google Scholar] [CrossRef] [PubMed]
- Stockley, M.L.; Ferdinand, A.; Benedetti, G.; Blencowe, P.; Boyd, S.M.; Calder, M.; Charles, M.D.; Edwardes, L.V.; Ekwuru, T.; Finch, H.; et al. Discovery, Characterization, and Structure-Based Optimization of Small-Molecule In Vitro and In Vivo Probes for Human DNA Polymerase Theta. J. Med. Chem. 2022, 65, 13879–13891. [Google Scholar] [CrossRef]
- Rodriguez-Berriguete, G.; Ranzani, M.; Prevo, R.; Puliyadi, R.; Bolland, H.R.; Millar, V.; Ebner, D.; Boursier, M.; Cerutti, A.; Cicconi, A.; et al. Small-Molecule Polθ Inhibitors Provide Safe and Effective Tumor Radiosensitization in 1 Preclinical Models 2. Clin. Cancer Res. 2023, 44, 617311. [Google Scholar] [CrossRef]
- Baxter, J.S.; Zatreanu, D.; Pettitt, S.J.; Lord, C.J. Resistance to DNA Repair Inhibitors in Cancer. Mol. Oncol. 2022, 16, 3811–3827. [Google Scholar] [CrossRef]
- Liddiard, K.; Aston-Evans, A.N.; Cleal, K.; Hendrickson, E.A.; Baird, D.M. POLQ Suppresses Genome Instability and Alterations in DNA Repeat Tract Lengths. NAR Cancer 2022, 4, zcac020. [Google Scholar] [CrossRef]
- Chen, X.S.; Pomerantz, R.T. Dna Polymerase θ: A Cancer Drug Target with Reverse Transcriptase Activity. Genes 2021, 12, 1146. [Google Scholar] [CrossRef]
- Dai, C.H.; Chen, P.; Li, J.; Lan, T.; Chen, Y.C.; Qian, H.; Chen, K.; Li, M.Y. Co-Inhibition of Pol θ and HR Genes Efficiently Synergize with Cisplatin to Suppress Cisplatin-Resistant Lung Cancer Cells Survival. Oncotarget 2016, 7, 65157–65170. [Google Scholar] [CrossRef] [PubMed]
- Chandramouly, G.; Liao, S.; Rusanov, T.; Borisonnik, N.; Calbert, M.L.; Kent, T.; Sullivan-Reed, K.; Vekariya, U.; Kashkina, E.; Skorski, T.; et al. Polθ Promotes the Repair of 5′-DNA-Protein Crosslinks by Microhomology-Mediated End-Joining. Cell Rep. 2021, 34, 108820. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Kim, J.; Haradhvala, N.J.; Huang, M.N.; Tian Ng, A.W.; Wu, Y.; Boot, A.; Covington, K.R.; Gordenin, D.A.; Bergstrom, E.N.; et al. The Repertoire of Mutational Signatures in Human Cancer. Nature 2020, 578, 94–101. [Google Scholar] [CrossRef]
- Hwang, T.; Reh, S.; Dunbayev, Y.; Zhong, Y.; Takata, Y.; Shen, J.; McBride, K.M.; Murnane, J.P.; Bhak, J.; Lee, S.; et al. Defining the Mutation Signatures of DNA Polymerase θ in Cancer Genomes. NAR Cancer 2020, 2, zcaa017. [Google Scholar] [CrossRef] [PubMed]
- Moore, G.; Majumdar, R.; Powell, S.N.; Khan, A.J.; Weinhold, N.; Yin, S.; Higginson, D.S. Templated Insertions Are Associated Specifically with BRCA2 Deficiency and Overall Survival in Advanced Ovarian Cancer. Mol. Cancer Res. 2022, 20, 1061–1070. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kruchinin, A.A.; Makarova, A.V. Multifaceted Nature of DNA Polymerase θ. Int. J. Mol. Sci. 2023, 24, 3619. https://doi.org/10.3390/ijms24043619
Kruchinin AA, Makarova AV. Multifaceted Nature of DNA Polymerase θ. International Journal of Molecular Sciences. 2023; 24(4):3619. https://doi.org/10.3390/ijms24043619
Chicago/Turabian StyleKruchinin, Alexander A., and Alena V. Makarova. 2023. "Multifaceted Nature of DNA Polymerase θ" International Journal of Molecular Sciences 24, no. 4: 3619. https://doi.org/10.3390/ijms24043619
APA StyleKruchinin, A. A., & Makarova, A. V. (2023). Multifaceted Nature of DNA Polymerase θ. International Journal of Molecular Sciences, 24(4), 3619. https://doi.org/10.3390/ijms24043619