

Cornelian Cherry (Cornus mas L.) Iridoid and Anthocyanin-Rich Extract Reduces Various Oxidation, Inflammation, and Adhesion Markers in a Cholesterol-Rich Diet Rabbit Model

 , , ,
, , ,  ,
,  , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Results
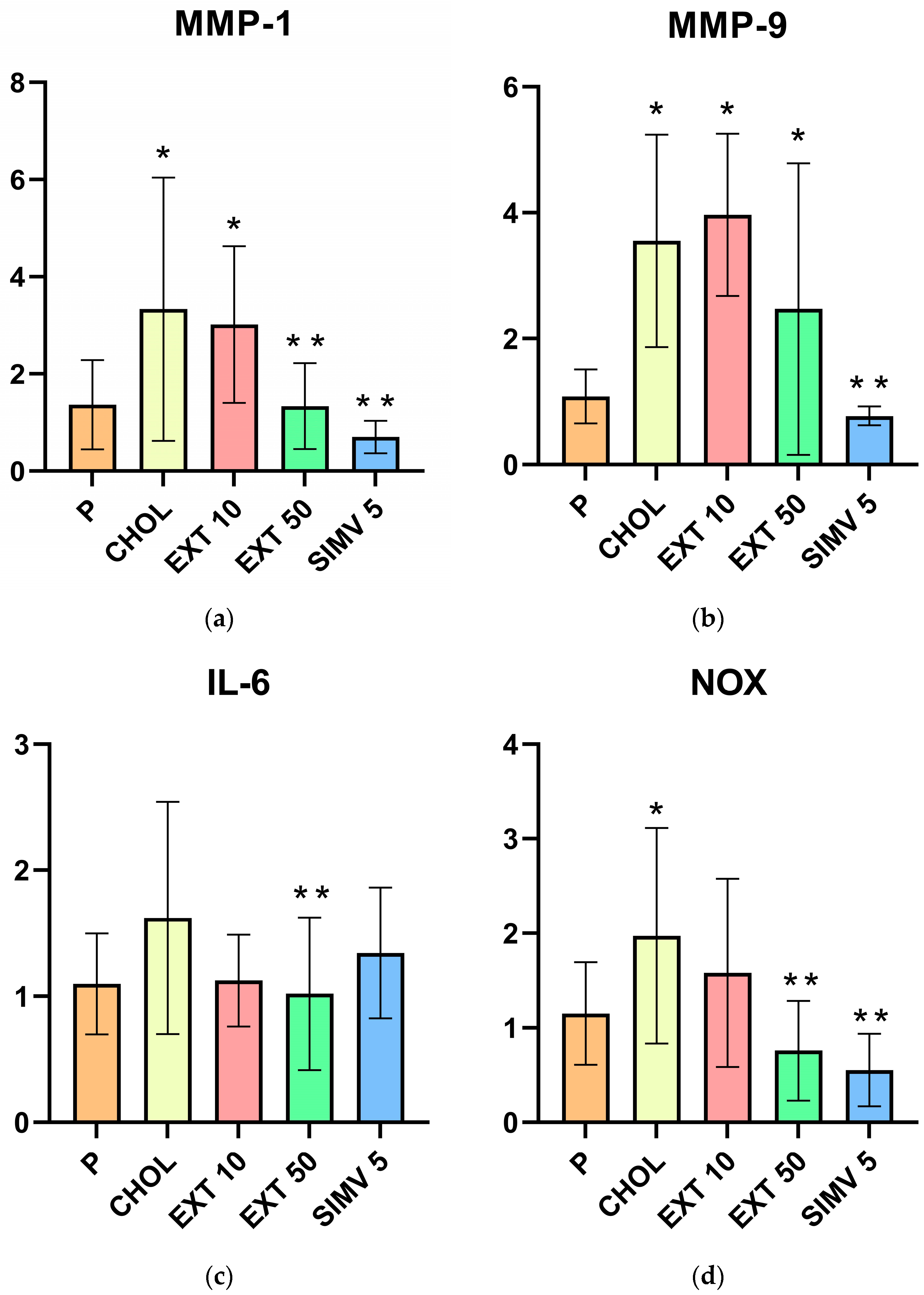
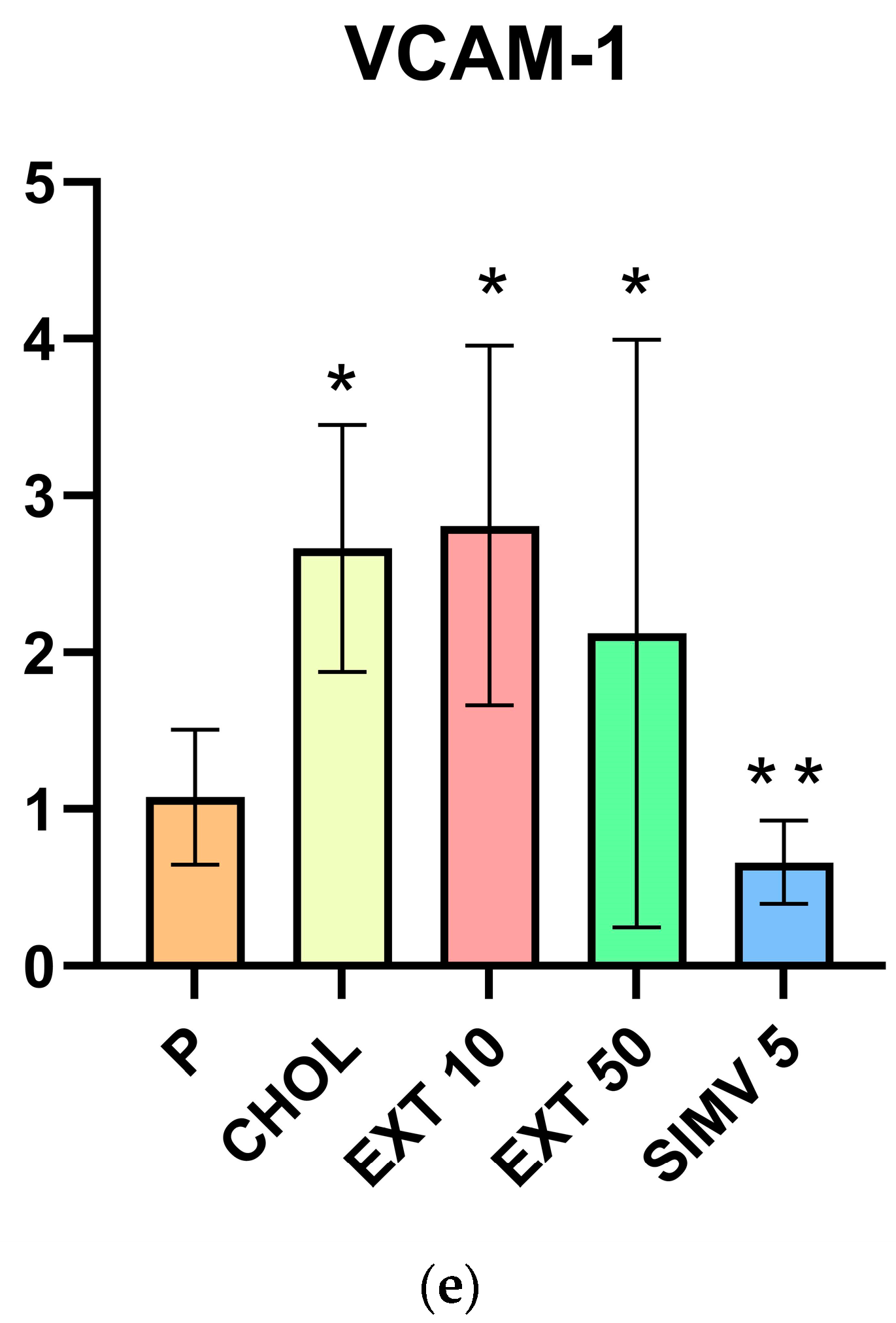
2.1. Assessment of MMP-1, MMP-9, IL-6, NOX, and VCAM-1 mRNA Expression in the Aorta by Real-Time PCR
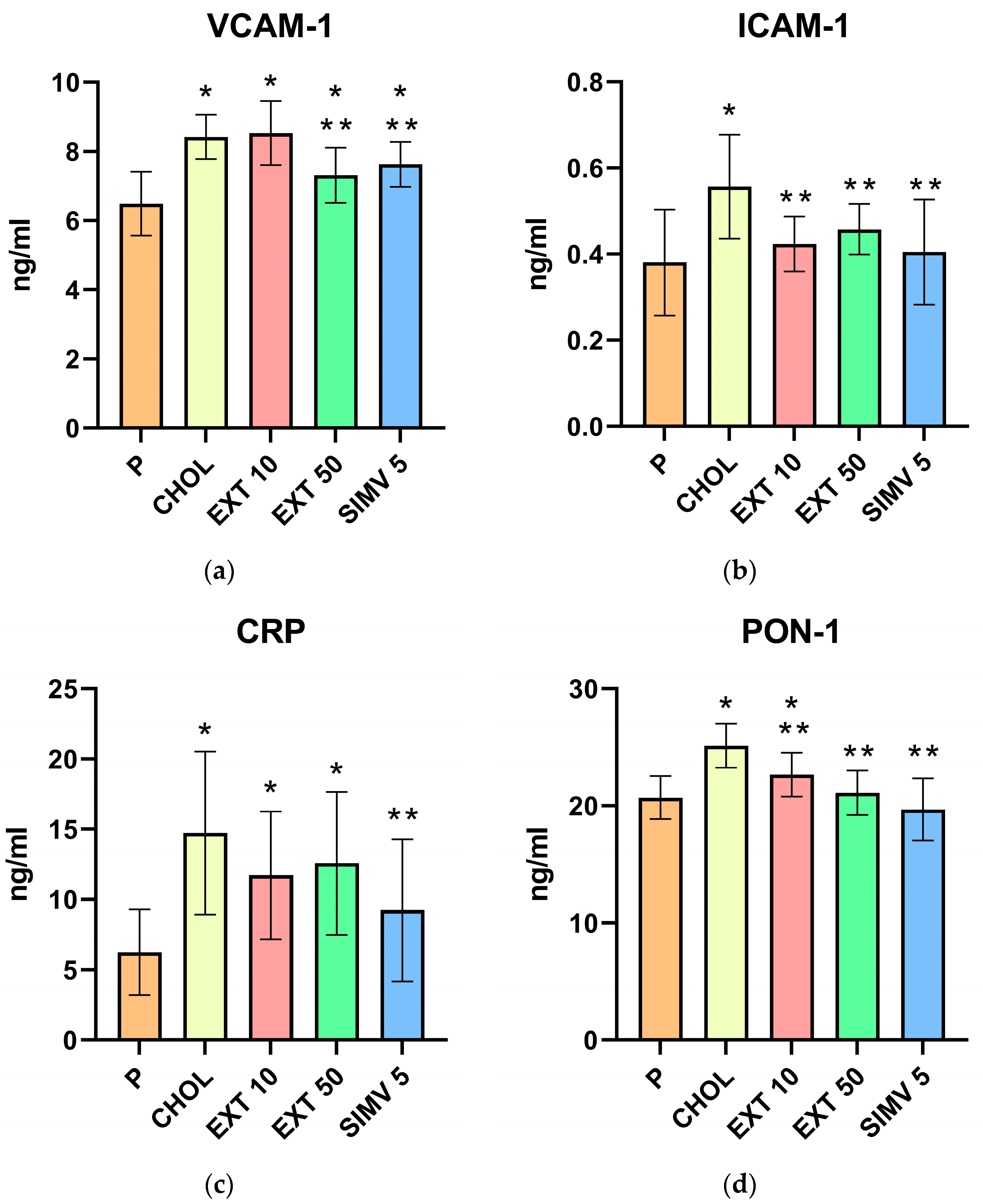
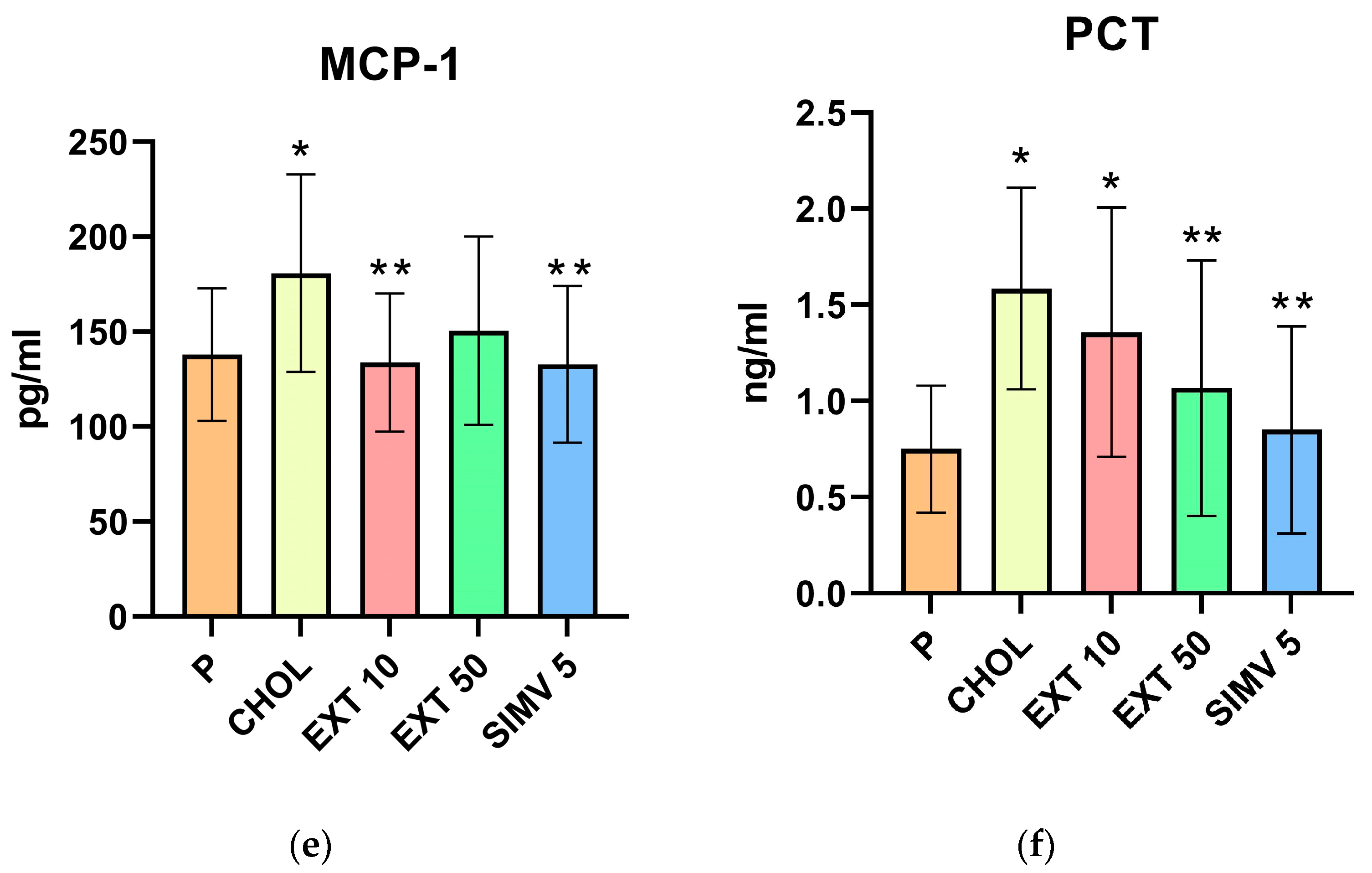
2.2. Assessment of VCAM-1, ICAM-1, CRP, PON-1, MCP-1, and PCT Serum Levels by ELISA
3. Discussion
4. Materials and Methods
4.1. Animal Model
4.2. Plant Materials and Preparation of Extract
4.3. RNA Isolation, Reverse Transcription, and Assessment of mRNA Expression of MMP-1, MMP-9, IL-6, NOX, and VCAM-1 in the Aorta by Real-Time PCR
4.4. Quantification of Serum Levels of VCAM-1, ICAM-1, CRP, PON-1, MCP-1, and PCT by Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wei, G.; Hao, L.; Li, X.; Xu, W.; Liu, F.; Peng, Q.; Lv, S. VCAM-1-targeted and PPARδ-agonist-loaded nanomicelles enhanced suppressing effects on apoptosis and migration of oxidized low-density lipoprotein-induced vascular smooth muscle cells. Biosci. Rep. 2020, 40, BSR20200559. [Google Scholar] [CrossRef] [Green Version]
- Bryk, D.; Olejarz, W.; Zapolska-Downar, D. The role of oxidative stress and NADPH oxidase in the pathogenesis of atherosclerosis. Postep. Hig. Med. Dosw. 2017, 71, 57–68. [Google Scholar] [CrossRef]
- Vermot, A.; Petit-Härtlein, I.; Smith, S.M.E.; Fieschi, F. NADPH Oxidases (NOX): An Overview from Discovery, Molecular Mechanisms to Physiology and Pathology. Antioxidants 2021, 10, 890. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Cross-Talk between NADPH Oxidase and Mitochondria: Role in ROS Signaling and Angiogenesis. Cells 2020, 9, 1849. [Google Scholar] [CrossRef]
- Singel, K.L.; Segal, B.H. NOX2-dependent regulation of inflammation. Clin. Sci. 2016, 130, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Malekmohammad, K.; Sewell, R.D.E.; Rafieian-Kopaei, M. Antioxidants, and Atherosclerosis: Mechanistic Aspects. Biomolecules 2019, 9, 301. [Google Scholar] [CrossRef] [Green Version]
- Mazzuca, M.Q.; Wlodek, M.E.; Dragomir, N.M.; Parkington, H.C.; Tare, M. Uteroplacental insufficiency programs regional vascular dysfunction and alters arterial stiffness in female offspring. J. Physiol. 2010, 588, 1997–2010. [Google Scholar] [CrossRef]
- Santos, J.C.D.; Cruz, M.S.; Bortolin, R.H.; Oliveira, K.M.; Araújo, J.N.G.; Duarte, V.H.R.; Silva, A.M.G.D.; Santos, I.C.C.D.; Dantas, J.M.O.; Paiva, M.S.M.O.; et al. Relationship between circulating VCAM-1, ICAM-1, E-selectin and MMP9 and the extent of coronary lesions. Clinics 2018, 73, e203. [Google Scholar] [CrossRef]
- Blankenberg, S.; Barbaux, S.; Tiret, L. Adhesion molecules and atherosclerosis. Atherosclerosis 2003, 170, 191–203. [Google Scholar] [CrossRef]
- Amin, M.; Pushpakumar, S.; Muradashvili, N.; Kundu, S.; Tyagi, S.C.; Sen, U. Regulation and involvement of matrix metalloproteinases in vascular diseases. Front. Biosci. 2016, 21, 89–118. [Google Scholar]
- Macarie, R.D.; Vadana, M.; Ciortan, L.; Tucureanu, M.M.; Ciobanu, A.; Vinereanu, D.; Manduteanu, I.; Simionescu, M.; Butoi, E. The expression of MMP-1 and MMP-9 is up-regulated by smooth muscle cells after their cross-talk with macrophages in high glucose conditions. J. Cell. Mol. Med. 2018, 22, 4366–4376. [Google Scholar] [CrossRef]
- Dollery, C.M.; Libby, P. Atherosclerosis and proteinase activation. Cardiovasc. Res. 2006, 69, 625–635. [Google Scholar] [CrossRef] [Green Version]
- Varani, J.; Perone, P.; Fligiel, S.E.; Fisher, G.J.; Voorhees, J.J. Inhibition of type I procollagen production in photodamage: Correlation between presence of high molecular weight collagen fragments and reduced procollagen synthesis. J. Investig. Dermatol. 2002, 119, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Newby, A.C. Dual role of matrix metalloproteinases (matrixins) in intimal thickening and atherosclerotic plaque rupture. Physiol. Rev. 2005, 85, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Bayrak, A.; Bayrak, T.; Tokgözoglu, S.L.; Volkan-Salanci, B.; Deniz, A.; Yavuz, B.; Alikasifoglu, M.; Demirpençe, E. Serum PON-1 activity but not Q192R polymorphism is related to the extent of atherosclerosis. J. Atheroscler. Thromb. 2012, 19, 376–384. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Mazhar, Z.; Alsayrafi, D.; Garelnabi, M. p-Cymene Modulate Oxidative Stress and Inflammation in Murine Macrophages: Potential Implication in Atherosclerosis. Cardiovasc. Hematol. Agents Med. Chem. 2020, 18, 151–157. [Google Scholar] [CrossRef]
- Sozański, T.; Kucharska, A.Z.; Rapak, A.; Szumny, D.; Trocha, M.; Merwid-Ląd, A.; Dzimira, S.; Piasecki, T.; Piórecki, N.; Magdalan, J.; et al. Iridoid-loganic acid versus anthocyanins from the Cornus mas fruits (cornelian cherry): Common and different effects on diet-induced atherosclerosis, PPARs expression and inflammation. Atherosclerosis 2016, 254, 151–160. [Google Scholar] [CrossRef]
- Sozański, T.; Kucharska, A.Z.; Szumny, D.; Magdalan, J.; Merwid-Ląd, A.; Nowak, B.; Piórecki, N.; Dzimira, S.; Jodkowska, A.; Szeląg, A.; et al. Cornelian cherry consumption increases the L-arginine/ADMA ratio, lowers ADMA and SDMA levels in the plasma, and enhances the aorta glutathione level in rabbits fed a high-cholesterol diet. J. Funct. Food. 2017, 34, 189–196. [Google Scholar] [CrossRef]
- Sozański, T.; Kucharska, A.Z.; Dzimira, S.; Magdalan, J.; Szumny, D.; Matuszewska, A.; Nowak, B.; Piórecki, N.; Szeląg, A.; Trocha, M. Loganic acid and anthocyanins from cornelian cherry (Cornus mas L.) fruits modulate diet-induced atherosclerosis and redox status in rabbits. Adv. Clin. Exp. Med. 2018, 27, 1505–1513. [Google Scholar] [CrossRef] [Green Version]
- Sozański, T.; Kucharska, A.Z.; Wiśniewski, J.; Fleszar, M.G.; Rapak, A.; Gomułkiewicz, A.; Dzięgiel, P.; Magdalan, J.; Nowak, B.; Szumny, D.; et al. The iridoid loganic acid and anthocyanins from the cornelian cherry (Cornus mas L.) fruit increase the plasma l-arginine/ADMA ratio and decrease levels of ADMA in rabbits fed a high-cholesterol diet. Phytomedicine 2019, 52, 1–11. [Google Scholar] [CrossRef]
- Nowak, B.; Matuszewska, A.; Tomanik, M.; Filipiak, J.; Kucharska, A.Z.; Piórecki, N.; Jędrzejuk, D.; Zduniak, K.; Trocha, M.; Bolanowski, M.; et al. Cornelian cherry extract ameliorates osteoporosis associated with hypercholesterolemia in New Zealand rabbits. Adv. Clin. Exp. Med. 2020, 29, 1389–1397. [Google Scholar] [CrossRef]
- Nowak, B.; Matuszewska, A.; Szeląg, A.; Danielewski, M.; Dziewiszek, W.; Nikodem, A.; Filipiak, J.; Jędrzejuk, J.; Bolanowski, M.; Kucharska, A.; et al. Cornelian cherry (Cornus mas L.) extract reduces cardiovascular risk and prevents bone loss in ovariectomized Wistar rats. J. Funct. Food. 2022, 90, 104974. [Google Scholar] [CrossRef]
- Szandruk-Bender, M.; Rutkowska, M.; Merwid-Ląd, A.; Wiatrak, B.; Szeląg, A.; Dzimira, S.; Sobieszczańska, B.; Krzystek-Korpacka, M.; Kucharska, A.Z.; Matuszewska, A.; et al. Cornelian Cherry Iridoid-Polyphenolic Extract Improves Mucosal Epithelial Barrier Integrity in Rat Experimental Colitis and Exerts Antimicrobial and Antiadhesive Activities in vitro. Oxid. Med. Cell. Longev. 2020, 2020, 7697851. [Google Scholar] [CrossRef]
- Danielewski, M.; Matuszewska, A.; Nowak, B.; Kucharska, A.Z.; Sozański, T. The Effects of Natural Iridoids and Anthocyanins on Selected Parameters of Liver and Cardiovascular System Functions. Oxid. Med. Cell. Longev. 2020, 2020, 2735790. [Google Scholar] [CrossRef]
- Park, E.; Sozański, T.; Lee, C.G.; Kucharska, A.Z.; Przybylska, D.; Piórecki, N.; Jeong, S.Y. A Comparison of the Antiosteoporotic Effects of Cornelian Cherry (Cornus mas L.) Extracts from Red and Yellow Fruits Containing Different Constituents of Polyphenols and Iridoids in Osteoblasts and Osteoclasts. Oxid. Med. Cell. Longev. 2022, 2022, 4122253. [Google Scholar] [CrossRef]
- Przybylska, D.; Kucharska, A.Z.; Sozański, T. A Review on Bioactive Iridoids in Edible Fruits—From Garden to Food and Pharmaceutical Products. Food Rev. Int. 2022. [Google Scholar] [CrossRef]
- Danielewski, M.; Kucharska, A.Z.; Matuszewska, A.; Rapak, A.; Gomułkiewicz, A.; Dzimira, S.; Dzięgiel, P.; Nowak, B.; Trocha, M.; Magdalan, J.; et al. Cornelian Cherry (Cornus mas L.) Iridoid and Anthocyanin Extract Enhances PPAR-α, PPAR-γ Expression and Reduces I/M Ratio in Aorta, Increases LXR-α Expression and Alters Adipokines and Triglycerides Levels in Cholesterol-Rich Diet Rabbit Model. Nutrients 2021, 13, 3621. [Google Scholar] [CrossRef]
- Rao, V.H.; Kansal, V.; Stoupa, S.; Agrawal, D.K. MMP-1 and MMP-9 regulate epidermal growth factor-dependent collagen loss in human carotid plaque smooth muscle cells. Physiol. Rep. 2014, 2, e00224. [Google Scholar] [CrossRef]
- Butoi, E.; Gan, A.M.; Tucureanu, M.M.; Stan, D.; Macarie, R.D.; Constantinescu, C.; Calin, M.; Simionescu, M.; Manduteanu, I. Cross-talk between macrophages and smooth muscle cells impairs collagen and metalloprotease synthesis and promotes angiogenesis. Biochim. Biophys. Acta 2016, 1863, 1568–1578. [Google Scholar] [CrossRef]
- Wells, J.M.; Gaggar, A.; Blalock, J.E. MMP generated matrikines. Matrix Biol. 2015, 44–46, 122–129. [Google Scholar] [CrossRef]
- Nackman, G.B.; Karkowski, F.J.; Halpern, V.J.; Gaetz, H.P.; Tilson, M.D. Elastin degradation products induce adventitial angiogenesis in the Anidjar/Dobrin rat aneurysm model. Surgery 1997, 122, 39–44. [Google Scholar] [CrossRef]
- Hinek, A.; Boyle, J.; Rabinovitch, M. Vascular smooth muscle cell detachment from elastin and migration through elastic laminae is promoted by chondroitin sulfate-induced “shedding” of the 67-kDa cell surface elastin binding protein. Exp. Cell. Res. 1992, 203, 344–353. [Google Scholar] [CrossRef]
- Hu, W.; Wei, R.; Wang, L.; Lu, J.; Liu, H.; Zhang, W. Correlations of MMP-1, MMP-3, and MMP-12 with the degree of atherosclerosis, plaque stability and cardiovascular and cerebrovascular events. Exp. Ther. Med. 2018, 15, 1994–1998. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Moon, B.R.; Kim, J.E.; Kim, H.J.; Zhang, T. Aqueous Extracts of Morus alba Root Bark and Cornus officinalis Fruit Protect against Osteoarthritis Symptoms in Testosterone-Deficient and Osteoarthritis-Induced Rats. Pharmaceutics 2020, 12, 1245. [Google Scholar] [CrossRef]
- Park, E.; Lee, C.G.; Yun, S.H.; Hwang, S.; Jeon, H.; Kim, J.; Yeo, S.; Jeong, H.; Yun, S.H.; Jeong, S.Y. Ameliorative Effects of Loganin on Arthritis in Chondrocytes and Destabilization of the Medial Meniscus-Induced Animal Model. Pharmaceuticals 2021, 14, 135. [Google Scholar] [CrossRef]
- Park, E.; Lee, C.G.; Han, S.J.; Yun, S.H.; Hwang, S.; Jeon, H.; Kim, J.; Choi, C.W.; Yang, S.; Jeong, S.Y. Antiosteoarthritic Effect of Morroniside in Chondrocyte Inflammation and Destabilization of Medial Meniscus-Induced Mouse Model. Int. J. Mol. Sci. 2021, 22, 2987. [Google Scholar] [CrossRef]
- Yang, Y.; Gu, Y.; Zhao, H.; Zhang, S. Loganin Attenuates Osteoarthritis in Rats by Inhibiting IL-1β-Induced Catabolism and Apoptosis in Chondrocytes Via Regulation of Phosphatidylinositol 3-Kinases (PI3K)/Akt. Med. Sci. Monit. 2019, 25, 4159–4168. [Google Scholar] [CrossRef]
- Kang, D.G.; Moon, M.K.; Lee, A.S.; Kwon, T.O.; Kim, J.S.; Lee, H.S. Cornuside suppresses cytokine-induced proinflammatory and adhesion molecules in the human umbilical vein endothelial cells. Biol. Pharm. Bull. 2007, 30, 1796–1799. [Google Scholar] [CrossRef] [Green Version]
- Fallon, M.E.; Hinds, M.T. Single cell morphological metrics and cytoskeletal alignment regulate VCAM-1 protein expression. Biochem. Biophys. Res. Commun. 2021, 555, 160–167. [Google Scholar] [CrossRef]
- Lawson, C.; Wolf, S. ICAM-1 signaling in endothelial cells. Pharmacol. Rep. 2009, 61, 22–32. [Google Scholar] [CrossRef]
- Sumagin, R.; Lomakina, E.; Sarelius, I.H. Leukocyte-endothelial cell interactions are linked to vascular permeability via ICAM-1-mediated signaling. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H969–H977. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M. Anticytokine Agents: Targeting Interleukin Signaling Pathways for the Treatment of Atherothrombosis. Circ. Res. 2019, 124, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.; Rosas, S.E. IL-6 Is Associated with Progression of Coronary Artery Calcification and Mortality in Incident Dialysis Patients. Am. J. Nephrol. 2021, 52, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Libby, P. Targeting Inflammatory Pathways in Cardiovascular Disease: The Inflammasome, Interleukin-1, Interleukin-6 and Beyond. Cells 2021, 10, 951. [Google Scholar] [CrossRef]
- Moriya, J. Critical roles of inflammation in atherosclerosis. J. Cardiol. 2019, 73, 22–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quah, Y.; Lee, S.J.; Lee, E.B.; Birhanu, B.T.; Ali, M.S.; Abbas, M.A.; Boby, N.; Im, Z.E.; Park, S.C. Cornus officinalis Ethanolic Extract with Potential Anti-Allergic, Anti-Inflammatory, and Antioxidant Activities. Nutrients 2020, 12, 3317. [Google Scholar] [CrossRef]
- Xu, R.; Zeng, M.; Wu, Y.; Wang, S.; Zhang, B.; Zhang, J.; Kan, Y.; Li, B.; Cao, B.; Zheng, X.; et al. Acetone Extract of Cornus officinalis Leaves Exerts Anti-Melanoma Effects via Inhibiting STAT3 Signaling. Onco Targets Ther. 2021, 14, 3487–3501. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Xing, L.; Sun, K.; Xiao, C.; Meng, X.; Yang, J. Loganin attenuates intestinal injury in severely burned rats by regulating the toll-like receptor 4/NF-κB signaling pathway. Exp. Ther. Med. 2020, 20, 591–598. [Google Scholar] [CrossRef]
- Liu, S.; Shen, H.; Li, J.; Gong, Y.; Bao, H.; Zhang, J.; Hu, L.; Wang, Z.; Gong, J. Loganin inhibits macrophage M1 polarization and modulates sirt1/NF-κB signaling pathway to attenuate ulcerative colitis. Bioengineered 2020, 11, 628–639. [Google Scholar] [CrossRef]
- Shi, J.Z.; Zheng, X.M.; Zhou, Y.F.; Yun, L.Y.; Luo, D.M.; Hao, J.J.; Liu, P.F.; Zhang, W.K.; Xu, J.K.; Yan, Y.; et al. Cornuside Is a Potential Agent against Alzheimer’s Disease via Orchestration of Reactive Astrocytes. Nutrients 2022, 14, 3179. [Google Scholar] [CrossRef]
- Tyrrell, D.J.; Blin, M.G.; Song, J.; Wood, S.C.; Zhang, M.; Beard, D.A.; Goldstein, D.R. Age-Associated Mitochondrial Dysfunction Accelerates Atherogenesis. Circ. Res. 2020, 126, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, D.J.; Goldstein, D.R. Ageing and atherosclerosis: Vascular intrinsic and extrinsic factors and potential role of IL-6. Nat. Rev. Cardiol. 2021, 18, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Burtenshaw, D.; Hakimjavadi, R.; Redmond, E.M.; Cahill, P.A. Nox, Reactive Oxygen Species and Regulation of Vascular Cell Fate. Antioxidants 2017, 6, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manea, A.; Simionescu, M. Nox enzymes and oxidative stress in atherosclerosis. Front. Biosci. Sch. 2012, 4, 651–670. [Google Scholar]
- Zhang, Y.; Murugesan, P.; Huang, K.; Cai, H. NADPH oxidases and oxidase crosstalk in cardiovascular diseases: Novel therapeutic targets. Nat. Rev. Cardiol. 2020, 17, 170–194. [Google Scholar] [CrossRef]
- Park, C.H.; Noh, J.S.; Tanaka, T.; Yokozawa, T. 7-O-galloyl-D-sedoheptulose ameliorates renal damage triggered by reactive oxygen species-sensitive pathway of inflammation and apoptosis. J. Pharm. Pharmacol. 2012, 64, 1730–1740. [Google Scholar] [CrossRef]
- Park, C.H.; Tanaka, T.; Kim, J.H.; Cho, E.J.; Park, J.C.; Shibahara, N.; Yokozawa, T. Hepato-protective effects of loganin, iridoid glycoside from Corni Fructus, against hyperglycemia-activated signaling pathway in liver of type 2 diabetic db/db mice. Toxicology 2011, 290, 14–21. [Google Scholar] [CrossRef]
- Lee, J.A.; Shin, M.R.; Roh, S.S. Corni Fructus Alleviates UUO-Induced Renal Fibrosis via TGF-β/Smad Signaling. BioMed Res. Int. 2022, 2022, 5780964. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, J.; Jiang, M.; Fu, Y.; Zhu, Y.; Jiao, N.; Liu, L.; Du, Q.; Wu, H.; Xu, H.; et al. Loganin and catalpol exert cooperative ameliorating effects on podocyte apoptosis upon diabetic nephropathy by targeting AGEs-RAGE signaling. Life Sci. 2020, 252, 117653. [Google Scholar] [CrossRef]
- Fang, W.J.; Feng, J.F.; Lu, X.M.; Lv, M.P.; Cao, S.S.; Li, R.F.; Li, Y.; Li, X.M. Effect of Cornus officinalis fruit core extract on the cardiac hypertrophy induced by two kidney two clip. Zhong Yao Cai Zhongyaocai J. Chin. Med. Mater. 2012, 35, 1985–1989. [Google Scholar]
- Libby, P. Inflammation in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2045–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asgary, S.; Kelishadi, R.; Rafieian-Kopaei, M.; Najafi, S.; Najafi, M.; Sahebkar, A. Investigation of the lipid-modifying and antiinflammatory effects of Cornus mas L. supplementation on dyslipidemic children and adolescents. Pediatr. Cardiol. 2013, 34, 1729–1735. [Google Scholar] [CrossRef] [PubMed]
- Shunmoogam, N.; Naidoo, P.; Chilton, R. Paraoxonase (PON)-1: A brief overview on genetics, structure, polymorphisms and clinical relevance. Vasc. Health Risk Manag. 2018, 14, 137–143. [Google Scholar] [CrossRef]
- Kunutsor, S.K.; Bakker, S.J.; James, R.W.; Dullaart, R.P. Serum paraoxonase-1 activity and risk of incident cardiovascular disease: The PREVEND study and meta-analysis of prospective population studies. Atherosclerosis 2016, 245, 143–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, T.; Deckert, V.; Aires, V.; Le Guern, N.; Proukhnitzky, L.; Patoli, D.; Lemaire, S.; Maquart, G.; Bataille, A.; Xolin, M.; et al. Human apolipoprotein C1 transgenesis reduces atherogenesis in hypercholesterolemic rabbits. Atherosclerosis 2021, 320, 10–18. [Google Scholar] [CrossRef]
- Francik, R.; Kryczyk, J.; Krośniak, M.; Berköz, M.; Sanocka, I.; Francik, S. The neuroprotective effect of cornus MAS on brain tissue of Wistar rats. Sci. World J. 2014, 2014, 847368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Kakkar, V.; Lu, X. Impact of MCP-1 in atherosclerosis. Curr. Pharm. Des. 2014, 20, 4580–4588. [Google Scholar] [CrossRef]
- Bianconi, V.; Sahebkar, A.; Atkin, S.L.; Pirro, M. The regulation and importance of monocyte chemoattractant protein-1. Curr. Opin. Hematol. 2018, 25, 44–51. [Google Scholar] [CrossRef]
- Bekkering, S.; van den Munckhof, I.; Nielen, T.; Rutten, J.; Graaf, J.; Joosten, L.; Netea, M.; Gomes, M.; Riksen, N. Innate immune cell activation in symptomatic and asymptomatic atherosclerosis in humans in vivo. Atherosclerosis 2016, 252, e256. [Google Scholar] [CrossRef]
- Danielewski, M.; Matuszewska, A.; Szeląg, A.; Sozański, T. The Impact of Anthocyanins and Iridoids on Transcription Factors Crucial for Lipid and Cholesterol Homeostasis. Int. J. Mol. Sci. 2021, 22, 6074. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, Y.; Zhao, D.; Feng, X.; Zhang, L.; Liu, C. Loganin prevents BV-2 microglia cells from Aβ1-42-induced inflammation via regulating TLR4/TRAF6/NF-κB axis. Cell Biol. Int. 2018, 42, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Babu, D.; Thapa, D.; Lee, J.S.; Park, S.Y.; Kim, A.R.; Kim, Y.H.; Yang, H.C.; Kim, J.A. Inhibitory effects of an aqueous extract of Cornus kousa Burg. leaves on TNF-alpha-induced chemokine expression and monocyte adhesion to human colonic epithelial cells. Arch. Pharm. Res. 2009, 32, 91–98. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Z.; Tang, Y.; Shan, W.; Wang, L.; Shi, J.; Luo, Q.; Duan, J.; Xu, G. Association between procalcitonin levels and carotid atherosclerosis in acute ischemic stroke patients. Int. J. Neurosci. 2018, 128, 237–242. [Google Scholar] [CrossRef]
- Ilhan, F.; Akbulut, H.; Karaca, I.; Godekmerdan, A.; Ilkay, E.; Bulut, V. Procalcitonin, c-reactive protein and neopterin levels in patients with coronary atherosclerosis. Acta Cardiol. 2005, 60, 361–365. [Google Scholar] [CrossRef]
- Schiopu, A.; Hedblad, B.; Engström, G.; Struck, J.; Morgenthaler, N.G.; Melander, O. Plasma procalcitonin and the risk of cardiovascular events and death: A prospective population-based study. J. Intern. Med. 2012, 272, 484–491. [Google Scholar] [CrossRef]
- Ertem, A.G.; Efe, T.H.; Yayla, Ç.; Akboğa, M.K.; Açar, B.; Ünal, S.; Kirbaş, Ö.; Sezer Tekce, Y.; Maden, O.; Selcuk, H.; et al. The Association between Serum Procalcitonin Levels and Severity of Coronary Artery Disease Assessed by SYNTAX Score in Patients with Acute Coronary Syndrome. Angiology 2017, 68, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Kraśniak, A.; Drozdz, M.; Pasowicz, M.; Chmiel, G.; Kowalczyk-Michałek, M.; Szumilak, D.; Podolec, P.; Klimeczek, P.; Konieczyńska, M.; Wicher-Muniak, E.; et al. Influence of microinflammation and oxidative stress on atherosclerosis progression and calcifications in cardiovascular system of hemodialyzed patients during two years follow-up. Przegl Lek. 2007, 64, 140–147. [Google Scholar]
- Ozkan, H.; Celik, O.; Hatipoglu, E.; Kantarci, F.; Kadioglu, P. Procalcitonin can be used as a marker of premature atherosclerosis in acromegaly. Pituitary 2012, 15, 358–364. [Google Scholar] [CrossRef]
- Kucharska, A.Z.; Szumny, A.; Sokół-Łętowska, A.; Piórecki, N.; Klymenko, S.V. Iridoids and anthocyanins in cornelian cherry (Cornus mas L.) cultivars. J. Food Compos. Anal. 2015, 40, 95–102. [Google Scholar] [CrossRef]
- Kucharska, A.Z.; Sokół-Łętowska, A.; Oszmiański, J.; Piórecki, N.; Fecka, I. Iridoids, phenolic compounds and antioxidant activity of edible honeysuckle berries (Lonicera caerulea var. kamtschatica Sevast.). Molecules 2017, 22, 405. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | MMP-1 | MMP-9 | IL-6 | NOX | VCAM-1 |
|---|---|---|---|---|---|
| P | 1.365 ± 0.918 | 1.082 ± 0.430 | 1.099 ± 0.400 | 1.153 ± 0.543 | 1.076 ± 0.429 |
| CHOL | 3.333 ± 2.709 | 3.556 ± 1.688 | 1.623 ± 0.921 | 1.971 ± 1.140 | 2.663 ± 0.787 |
| EXT 10 | 3.017 ± 1.613 | 3.969 ± 1.287 | 1.125 ± 0.363 | 1.580 ± 0.995 | 2.807 ± 1.148 |
| EXT 50 | 1.337 ± 0.885 | 2.470 ± 2.316 | 1.021 ± 0.606 | 0.757 ± 0.528 | 2.122 ± 1.872 |
| SIMV 5 | 0.702 ± 0.335 | 0.773 ± 0.153 | 1.344 ± 0.518 | 0.553 ± 0.384 | 0.662 ± 0.266 |
| Group | VCAM-1 | ICAM-1 | CRP | PON-1 | MCP-1 | PCT |
|---|---|---|---|---|---|---|
| P | 6.490 ± 0.923 | 0.381 ± 0.123 | 6.241 ± 3.039 | 20.700 ± 1.828 | 137.949 ± 34.880 | 0.750 ± 0.331 |
| CHOL | 8.421 ± 0.644 | 0.557 ± 0.121 | 14.734 ± 5.797 | 25.135 ± 1.868 | 180.805 ± 51.942 | 1.585 ± 0.524 |
| EXT 10 | 8.532 ± 0.931 | 0.424 ± 0.064 | 11.720 ± 4.535 | 22.660 ± 1.880 | 133.751 ± 36.279 | 1.358 ± 0.649 |
| EXT 50 | 7.310 ± 0.801 | 0.458 ± 0.059 | 12.568 ± 5.080 | 21.114 ± 1.904 | 150.531 ± 49.552 | 1.068 ± 0.665 |
| SIMV 5 | 7.630 ± 0.648 | 0.405 ± 0.122 | 9.239 ± 5.052 | 19.684 ± 2.643 | 132.770 ± 41.303 | 0.850 ± 0.539 |
| Group | Chow | Dose of Tested Substance |
|---|---|---|
| P | standard chow | none (normal saline solution) |
| CHOL | standard chow +1% cholesterol | none (normal saline solution) |
| EXT 10 | standard chow +1% cholesterol | Cornelian cherry extract 10 mg/kg bw |
| EXT 50 | standard chow +1% cholesterol | Cornelian cherry extract 50 mg/kg bw |
| SIMV 5 | standard chow +1% cholesterol | simvastatin 5 mg/kg bw |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danielewski, M.; Gomułkiewicz, A.; Kucharska, A.Z.; Matuszewska, A.; Nowak, B.; Piórecki, N.; Trocha, M.; Szandruk-Bender, M.; Jawień, P.; Szeląg, A.; et al. Cornelian Cherry (Cornus mas L.) Iridoid and Anthocyanin-Rich Extract Reduces Various Oxidation, Inflammation, and Adhesion Markers in a Cholesterol-Rich Diet Rabbit Model. Int. J. Mol. Sci. 2023, 24, 3890. https://doi.org/10.3390/ijms24043890
Danielewski M, Gomułkiewicz A, Kucharska AZ, Matuszewska A, Nowak B, Piórecki N, Trocha M, Szandruk-Bender M, Jawień P, Szeląg A, et al. Cornelian Cherry (Cornus mas L.) Iridoid and Anthocyanin-Rich Extract Reduces Various Oxidation, Inflammation, and Adhesion Markers in a Cholesterol-Rich Diet Rabbit Model. International Journal of Molecular Sciences. 2023; 24(4):3890. https://doi.org/10.3390/ijms24043890
Chicago/Turabian StyleDanielewski, Maciej, Agnieszka Gomułkiewicz, Alicja Z. Kucharska, Agnieszka Matuszewska, Beata Nowak, Narcyz Piórecki, Małgorzata Trocha, Marta Szandruk-Bender, Paulina Jawień, Adam Szeląg, and et al. 2023. "Cornelian Cherry (Cornus mas L.) Iridoid and Anthocyanin-Rich Extract Reduces Various Oxidation, Inflammation, and Adhesion Markers in a Cholesterol-Rich Diet Rabbit Model" International Journal of Molecular Sciences 24, no. 4: 3890. https://doi.org/10.3390/ijms24043890
APA StyleDanielewski, M., Gomułkiewicz, A., Kucharska, A. Z., Matuszewska, A., Nowak, B., Piórecki, N., Trocha, M., Szandruk-Bender, M., Jawień, P., Szeląg, A., Dzięgiel, P., & Sozański, T. (2023). Cornelian Cherry (Cornus mas L.) Iridoid and Anthocyanin-Rich Extract Reduces Various Oxidation, Inflammation, and Adhesion Markers in a Cholesterol-Rich Diet Rabbit Model. International Journal of Molecular Sciences, 24(4), 3890. https://doi.org/10.3390/ijms24043890