
NUF2 Promotes Breast Cancer Development as a New Tumor Stem Cell Indicator

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
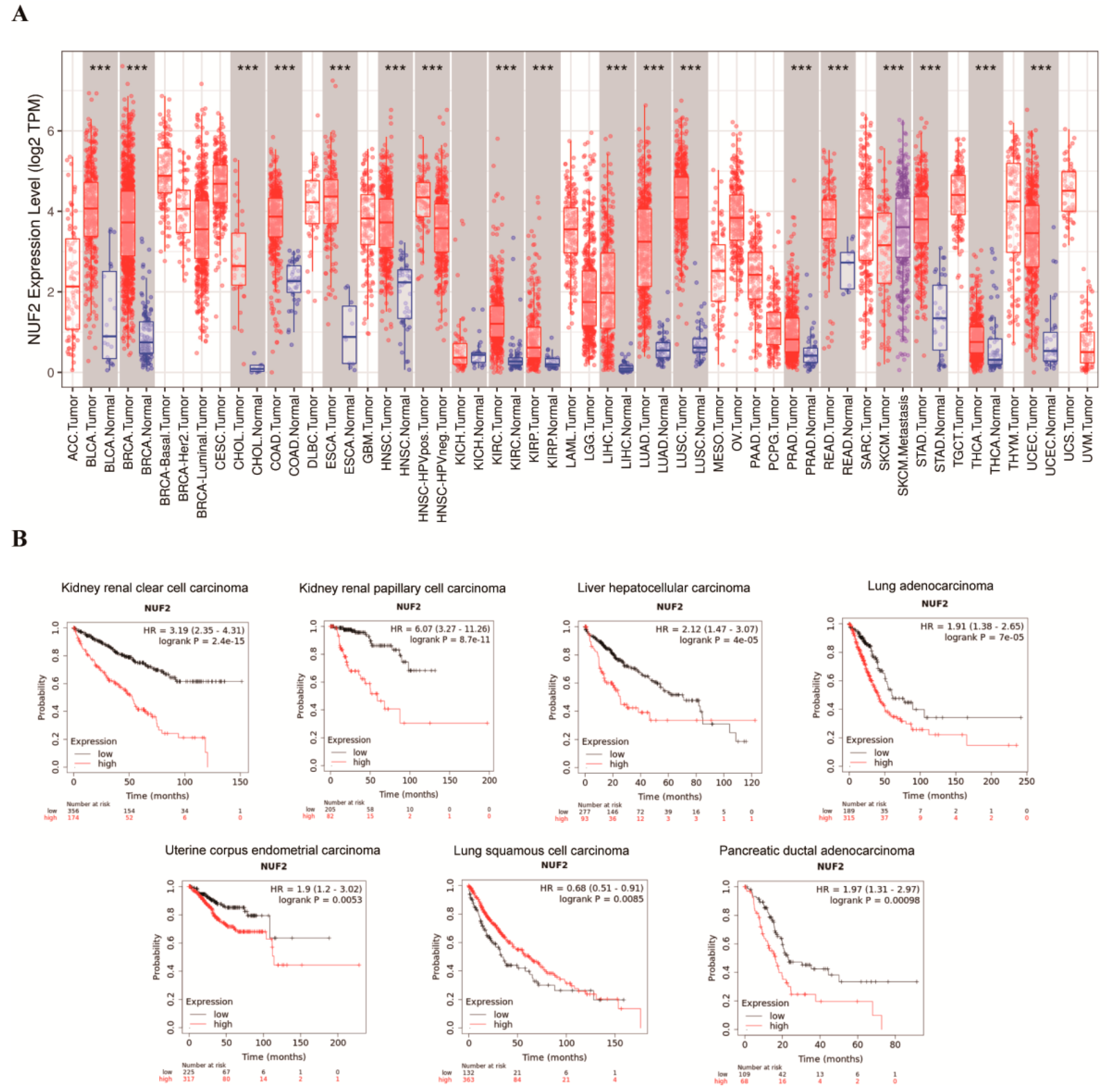
2.1. Expression Pattern of NUF2 from a Pan-Cancer Perspective
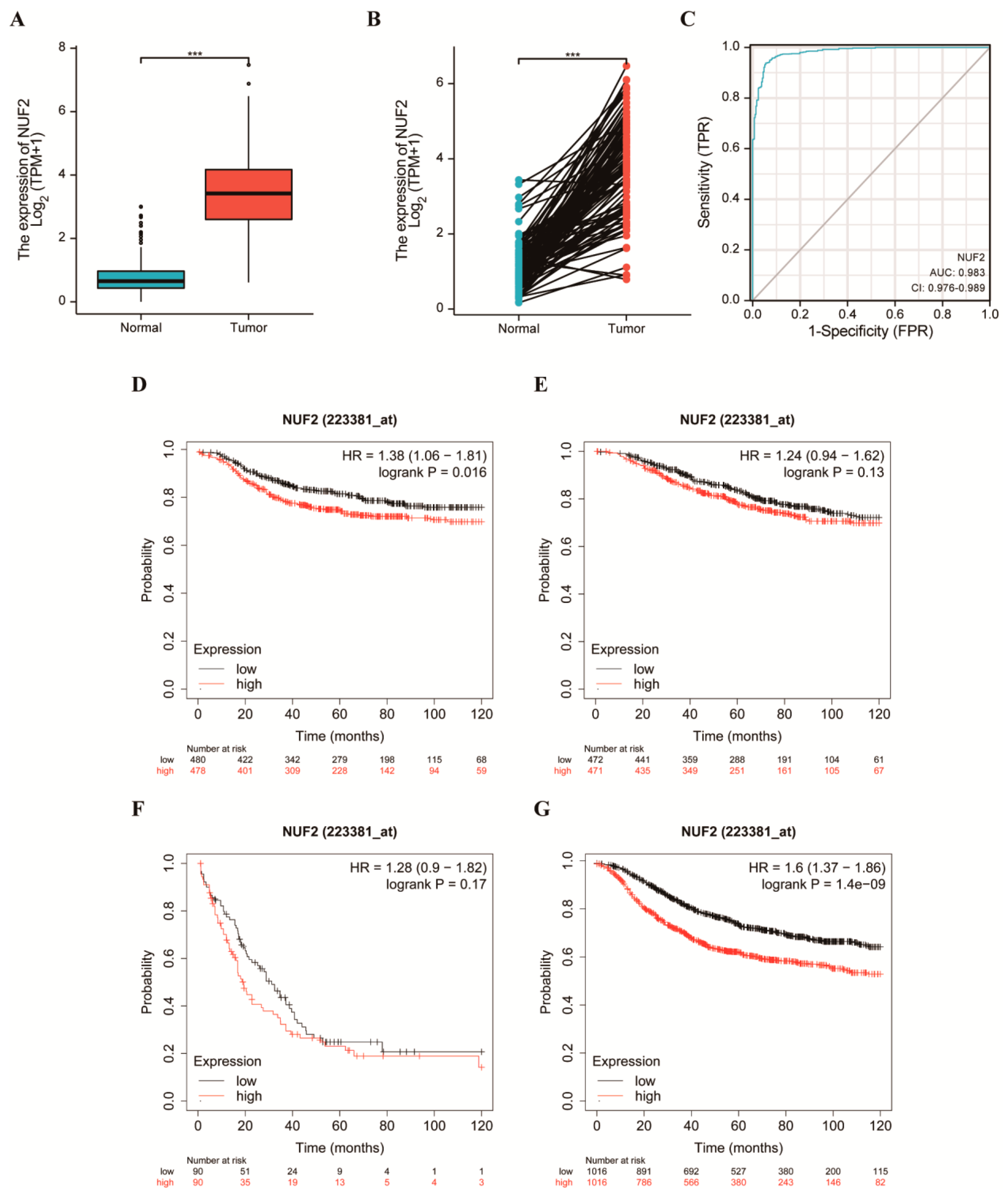
2.2. Elevated NUF2 Expression and Worse Prognosis in Patients with Breast Cancer
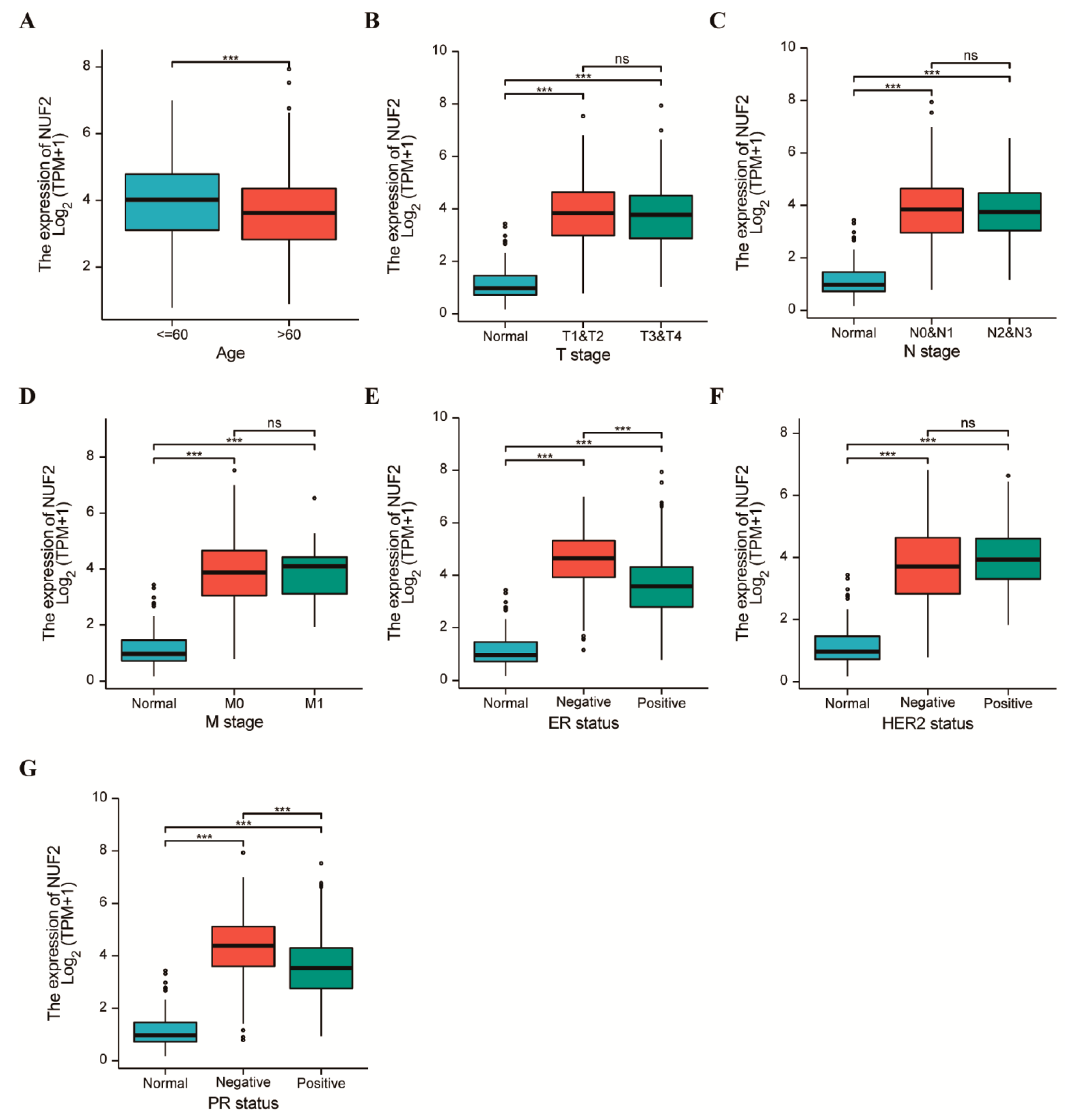
2.3. Clinicopathological Features of Breast Cancer with Respect to the NUF2 Expression Level
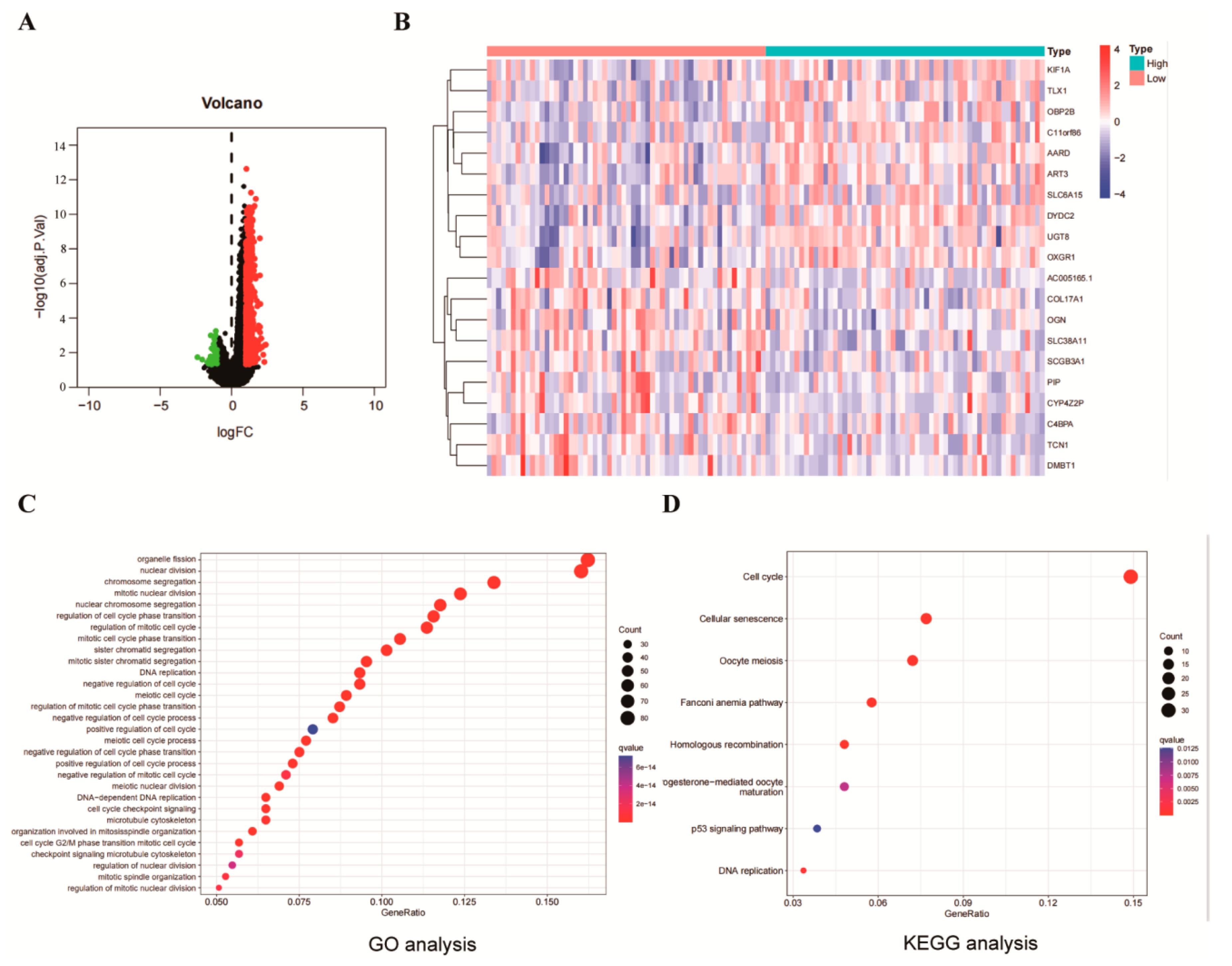
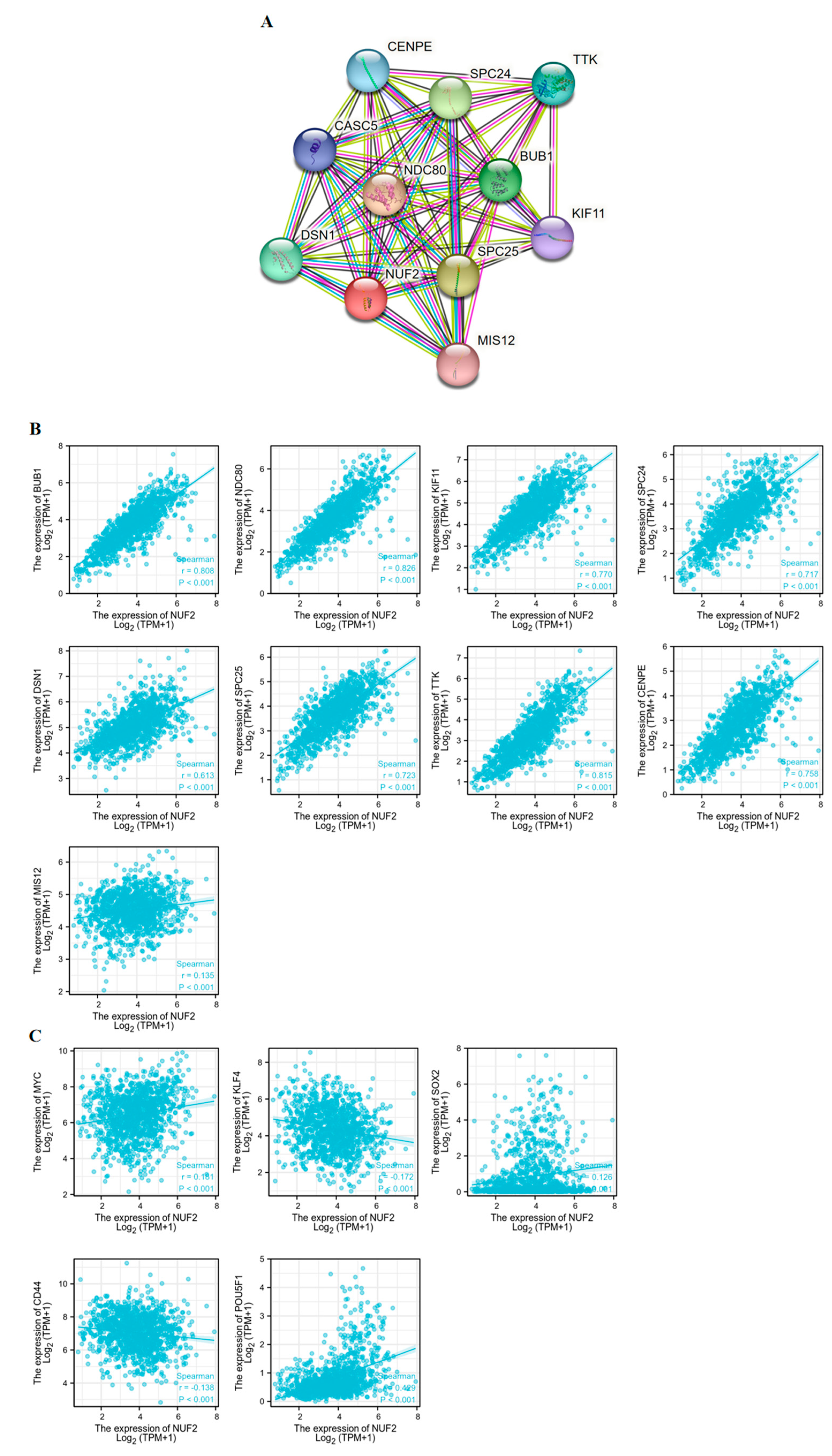
2.4. Identification and Functional Enrichment of NUF2 Co-Expressed Genes
2.5. Correlation between NUF2 and Tumor Stemness Markers
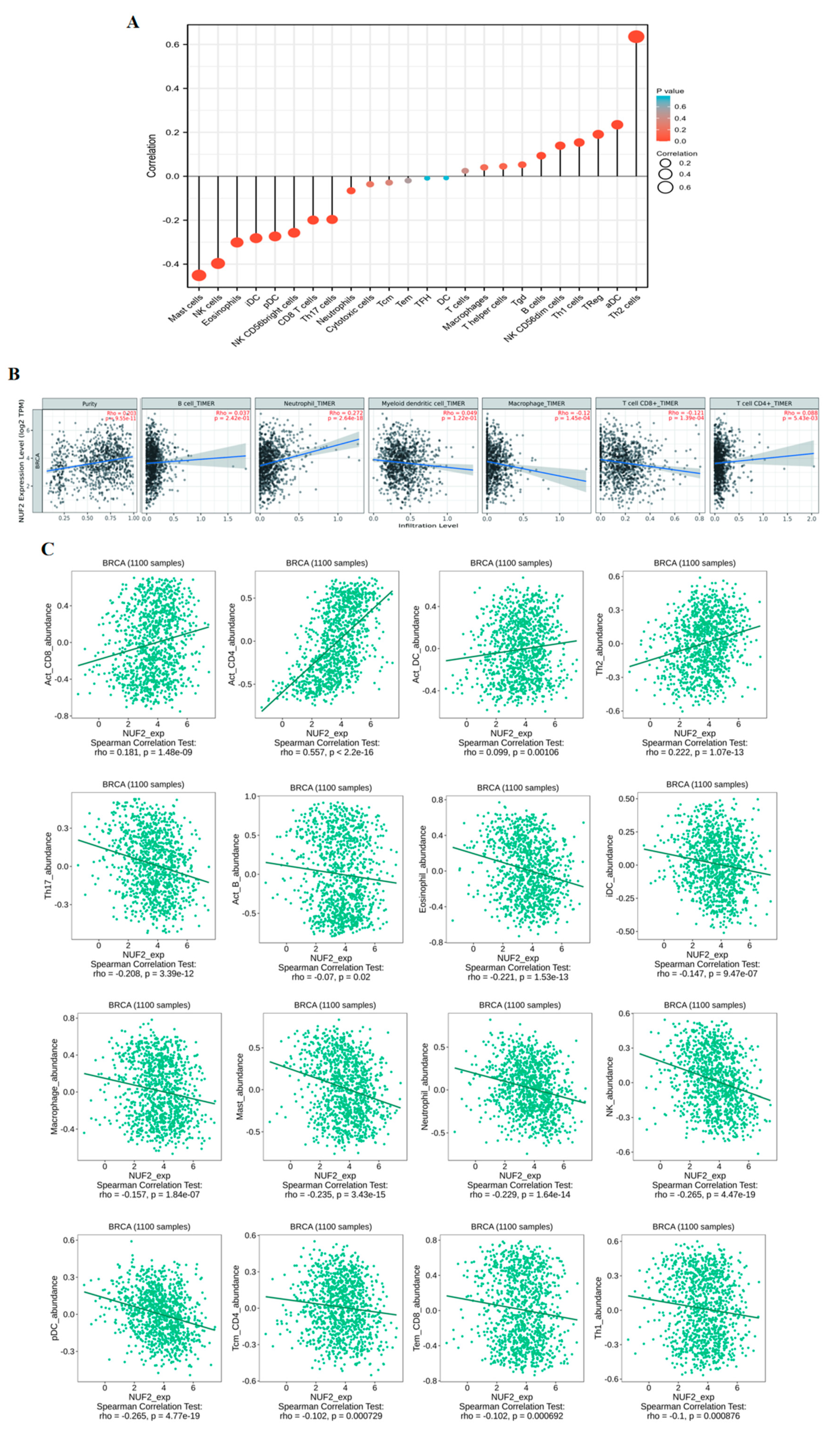
2.6. The Effect of NUF2 on Immune Cell Infiltration in Breast Cancer
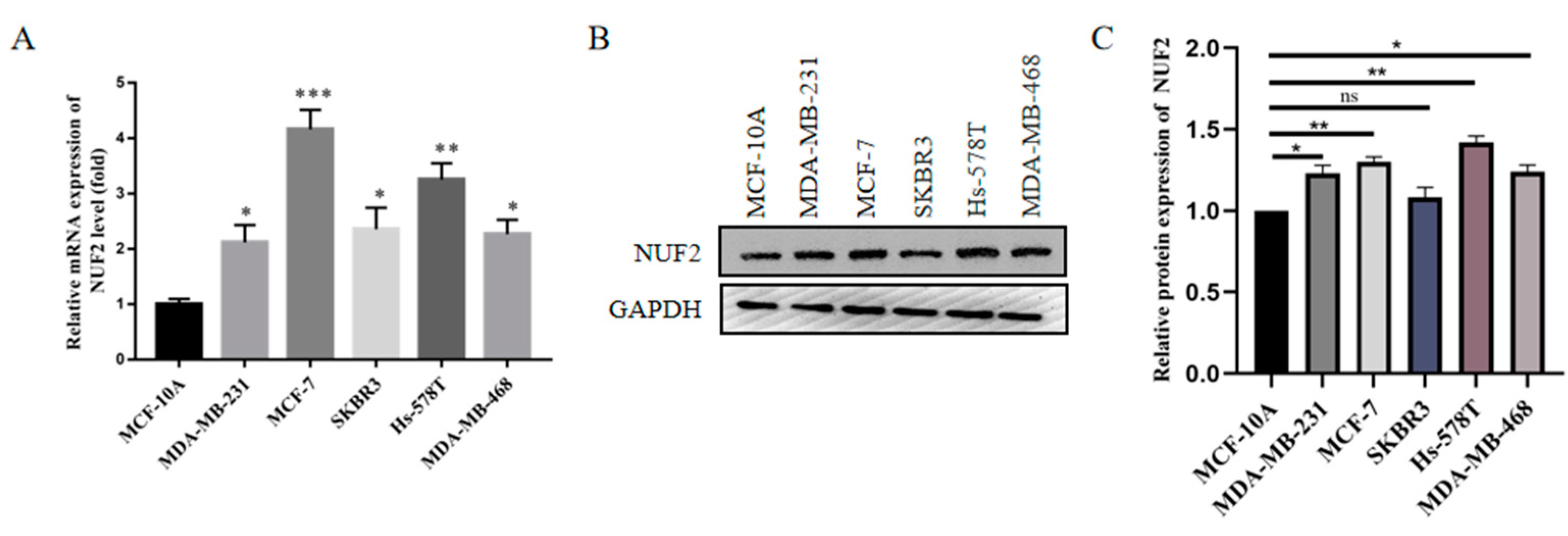
2.7. NUF2 Expression in Breast Cancer Cell Lines
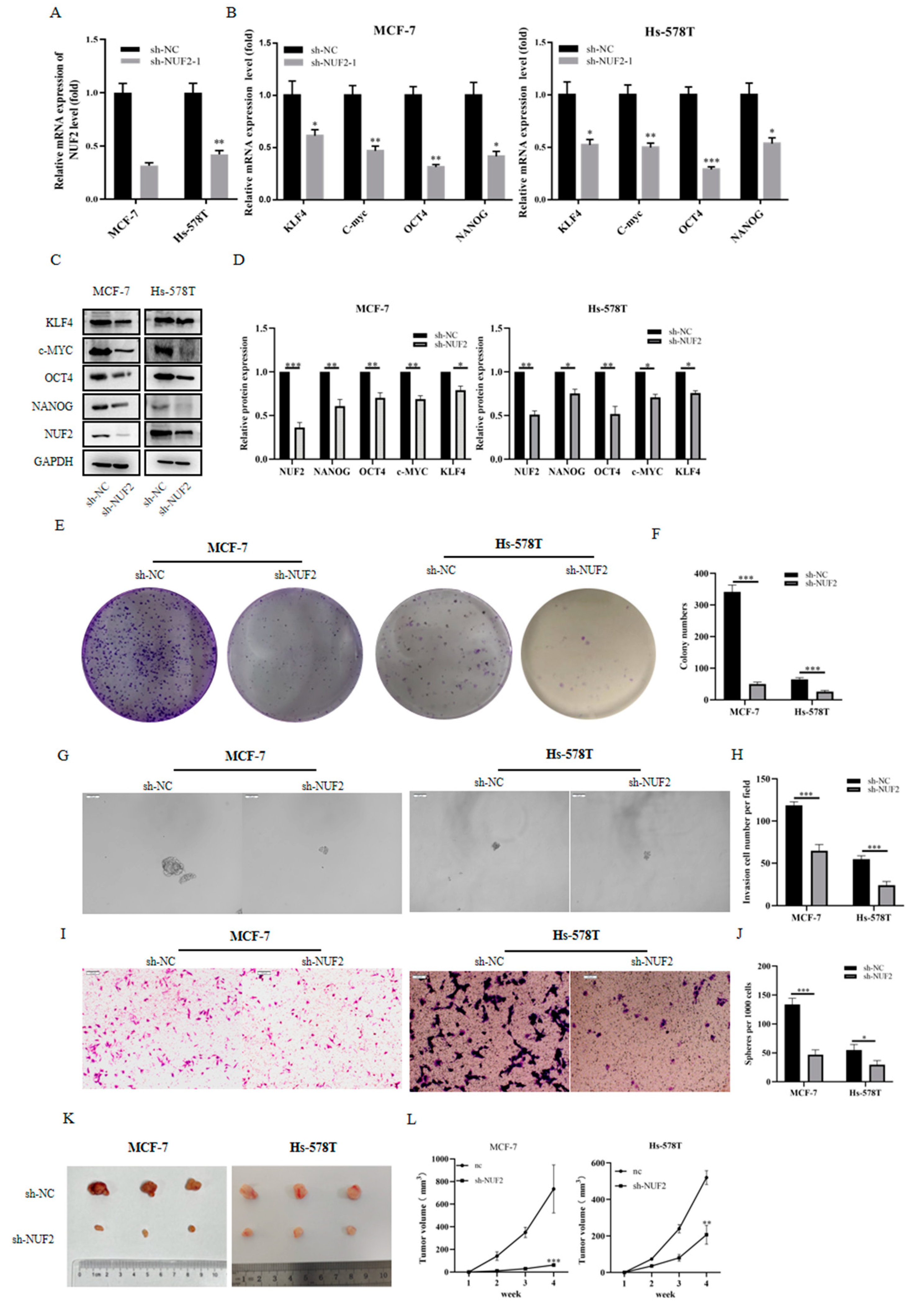
2.8. The Efficiency of NUF2 Knockdown in MCF-7 Cells
2.9. Downregulated/Inhibited Tumor Stemness as a Result of NUF2 Knockdown in Breast Cancer Cell Lines
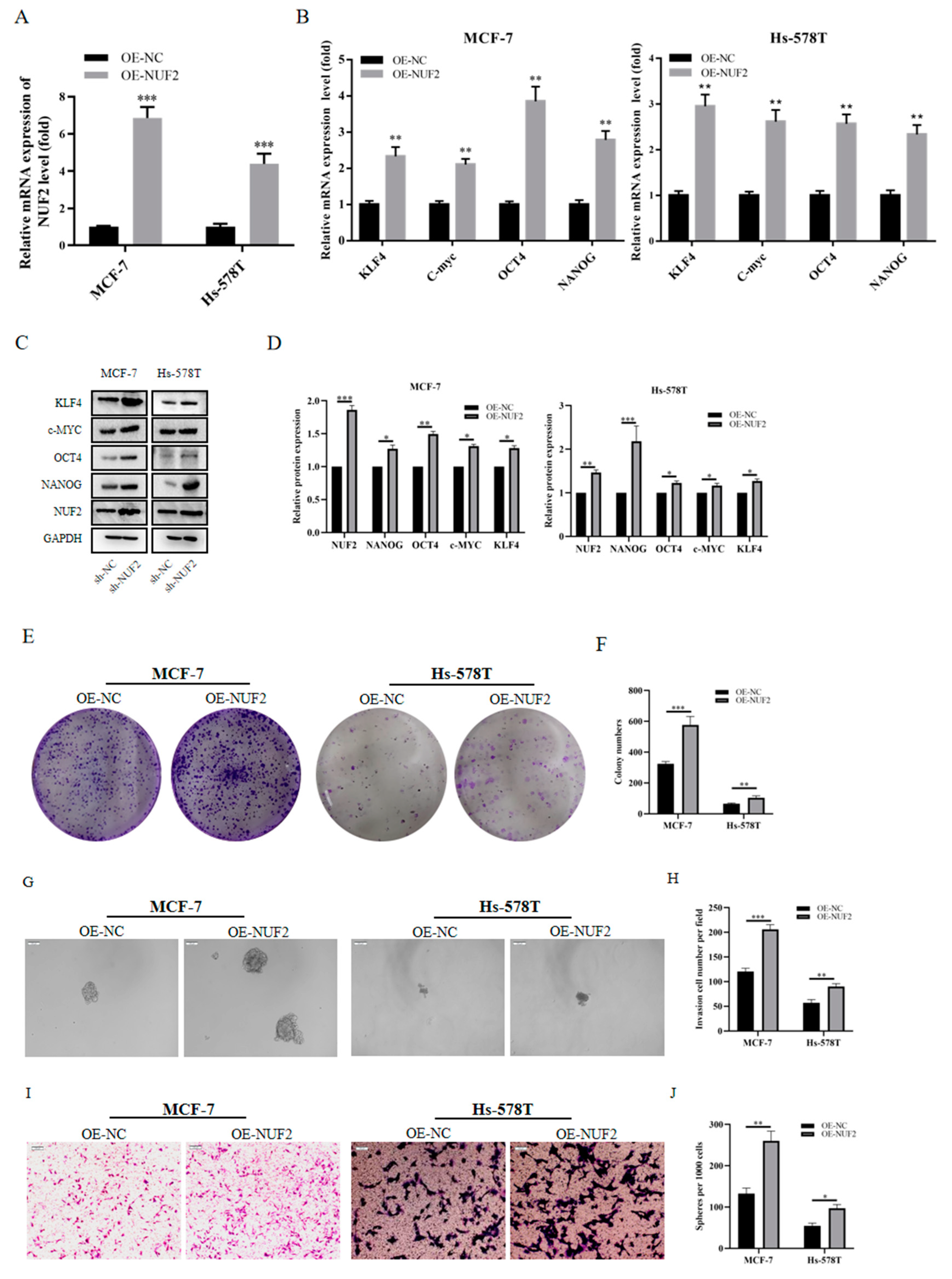
2.10. Tumor Stemness induced by NUF2 Overexpression in Breast Cancer Cell Lines
3. Discussion
4. Materials and Methods
4.1. Research Workflow
4.2. TCGA Data Acquisition
4.3. NUF2 mRNA Expression Data and Prognosis
4.4. Pathway Analysis
4.5. Protein–Protein Interaction (PPI) Network Analysis
4.6. Tumor Immune Assessment
4.7. Cell Lines
4.8. qRT-PCR Analysis
4.9. Cell Line Construction
4.10. Western Blot
4.11. Clone Formation Assay
4.12. Migration Assay
4.13. Spheroid Experiment
4.14. Nude Mouse Xenograft Model
4.15. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Loibl, S.; Poortmans, P. Breast cancer. Lancet 2021, 397, 1750–1769. [Google Scholar] [CrossRef] [PubMed]
- Brenton, J.D.; Carey, L.A.; Ahmed, A.A.; Caldas, C. Molecular classification and molecular forecasting of breast cancer: Ready for clinical application? J. Clin. Oncol. 2005, 23, 7350–7360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørlie, T.; Perou, C.M. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Minckwitz, G.; Procter, M. Adjuvant Pertuzumab and Trastuzumab in Early HER2-Positive Breast Cancer. N. Engl. J. Med. 2017, 377, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Nabetani, A.; Koujin, T. A conserved protein, NUF2, is implicated in connecting the centromere to the spindle during chromosome segregation: A link between the kinetochore function and the spindle checkpoint. Chromosoma 2001, 110, 322–334. [Google Scholar] [CrossRef] [PubMed]
- Riggio, A.I.; Varley, K.E.; Welm, A.L. The lingering mysteries of metastatic recurrence in breast cancer. Br. J. Cancer 2021, 124, 13–26. [Google Scholar] [CrossRef]
- Wei, R.R.; Al-Bassam, J.; Harrison, S.C. The Ndc80/HEC1 complex is a contact point for kinetochore-microtubule attachment. Nat. Struct. Mol. Biol. 2007, 14, 54–59. [Google Scholar] [CrossRef]
- Sun, J.; Chen, J. Expression of NUF2 in breast cancer and its clinical significance. Nan Fang Yi Ke Da Xue Xue Bao 2019, 39, 591–597. [Google Scholar]
- Ciferri, C.; Pasqualato, S. Implications for kinetochore-microtubule attachment from the structure of an engineered Ndc80 complex. Cell 2008, 133, 427–439. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.; Xu, W. NUF2 as an anticancer therapeutic target and prognostic factor in breast cancer. Int. J. Oncol. 2020, 57, 1358–1367. [Google Scholar] [CrossRef]
- Chen, M.; Li, S. Integrative Multi-Omics Analysis of Identified NUF2 as a Candidate Oncogene Correlates With Poor Prognosis and Immune Infiltration in Non-Small Cell Lung Cancer. Front. Oncol. 2021, 11, 656509. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, P.Y. NUF2 is a valuable prognostic biomarker to predict early recurrence of hepatocellular carcinoma after surgical resection. Int. J. Cancer 2019, 145, 662–670. [Google Scholar] [CrossRef]
- Guo, L.; Wang, Z. Random-forest algorithm based biomarkers in predicting prognosis in the patients with hepatocellular carcinoma. Cancer Cell Int. 2020, 20, 251. [Google Scholar] [CrossRef]
- Shi, L.; Shang, X. Identification of potential crucial genes associated with the pathogenesis and prognosis of liver hepatocellular carcinoma. J. Clin. Pathol. 2021, 74, 504–512. [Google Scholar] [CrossRef]
- Zheng, L.; Li, L. Six Novel Biomarkers for Diagnosis and Prognosis of Esophageal squamous cell carcinoma: Validated by scRNA-seq and qPCR. J. Cancer 2021, 12, 899–911. [Google Scholar] [CrossRef]
- Shan, L.; Zhu, X.L. Expression and clinical significance of NUF2 in kidney renal clear cell carcinoma. Transl. Urol. 2021, 10, 3628–3637. [Google Scholar] [CrossRef]
- Tomczak, K.; Czerwińska, P.; Wiznerowicz, M. The Cancer Genome Atlas (TCGA): An immeasurable source of knowledge. Contemp. Oncol. (Pozn) 2015, 19, A68–A77. [Google Scholar] [CrossRef]
- Liu, D.; Xu, X. Integrated Genome-Wide Analysis of Gene Expression and DNA Copy Number Variations Highlights Stem Cell-Related Pathways in Small Cell Esophageal Carcinoma. Stem Cells Int. 2018, 2018, 3481783. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Huang, X. NUF2 Expression Promotes Lung Adenocarcinoma Progression and Is Associated With Poor Prognosis. Front. Oncol. 2022, 12, 795971. [Google Scholar] [CrossRef]
- Li, X.; Zhang, L. NUF2 Is a Potential Immunological and Prognostic Marker for Non-Small-Cell Lung Cancer. J. Immunol. Res. 2022, 2022, 1161931. [Google Scholar] [CrossRef]
- Xie, X.; Jiang, S.; Li, X. NUF2 Is a Prognostic-Related Biomarker and Correlated With Immune Infiltrates in Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 621373. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, D561–D568. [Google Scholar] [CrossRef] [Green Version]
- Houghton, S.C.; Hankinson, S.E. Cancer Progress and Priorities: Breast Cancer. Cancer Epidemiol. Biomark. Prev. 2021, 30, 822–844. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Zhou, H. Role, molecular mechanism and the potential target of breast cancer stem cells in breast cancer development. Biomed. Pharm. 2022, 147, 112616. [Google Scholar] [CrossRef]
- Xu, W.; Wang, Y. Screening of differentially expressed genes and identification of NUF2 as a prognostic marker in breast cancer. Int. J. Mol. Med. 2019, 44, 390–404. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, T.; Terada, N. Cancer/Testis Antigens as potential predictors of biochemical recurrence of prostate cancer following radical prostatectomy. J. Transl. Med. 2011, 9, 153. [Google Scholar] [CrossRef] [Green Version]
- Bashiri, H.; Rahmani, H. EMDIP: An Entropy Measure to Discover Important Proteins in PPI networks. Comput. Biol. Med. 2020, 120, 103740. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fan, J. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Y.; Li, J.; Zhang, Y.; Hu, H.; Wan, F.; Min, H.; Zhou, H.; Gu, L.; Liao, X.; Zhou, J.; et al. NUF2 Promotes Breast Cancer Development as a New Tumor Stem Cell Indicator. Int. J. Mol. Sci. 2023, 24, 4226. https://doi.org/10.3390/ijms24044226
Deng Y, Li J, Zhang Y, Hu H, Wan F, Min H, Zhou H, Gu L, Liao X, Zhou J, et al. NUF2 Promotes Breast Cancer Development as a New Tumor Stem Cell Indicator. International Journal of Molecular Sciences. 2023; 24(4):4226. https://doi.org/10.3390/ijms24044226
Chicago/Turabian StyleDeng, Yang, Jiapeng Li, Yingjie Zhang, Hao Hu, Fujian Wan, Hang Min, Hao Zhou, Lixing Gu, Xinghua Liao, Jingjiao Zhou, and et al. 2023. "NUF2 Promotes Breast Cancer Development as a New Tumor Stem Cell Indicator" International Journal of Molecular Sciences 24, no. 4: 4226. https://doi.org/10.3390/ijms24044226
APA StyleDeng, Y., Li, J., Zhang, Y., Hu, H., Wan, F., Min, H., Zhou, H., Gu, L., Liao, X., Zhou, J., & Zhou, J. (2023). NUF2 Promotes Breast Cancer Development as a New Tumor Stem Cell Indicator. International Journal of Molecular Sciences, 24(4), 4226. https://doi.org/10.3390/ijms24044226