

Exogenous Ethylene Alleviates the Inhibition of Sorbus pohuashanensis Embryo Germination in a Saline-Alkali Environment (NaHCO3)


Abstract
:1. Introduction
2. Results
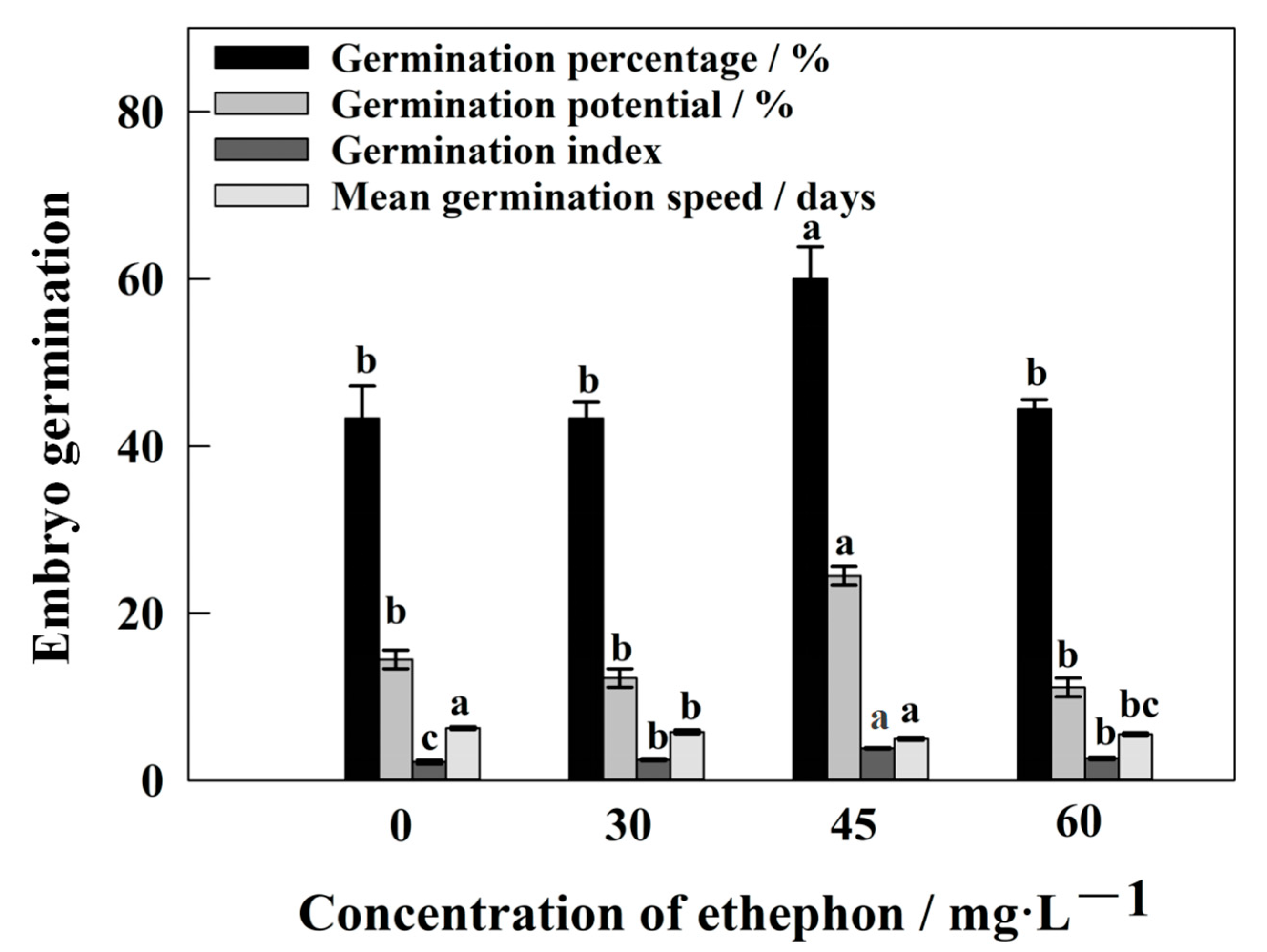
2.1. Effects of ETH and NaHCO3 on Germination of S. pohuashanensis Embryos
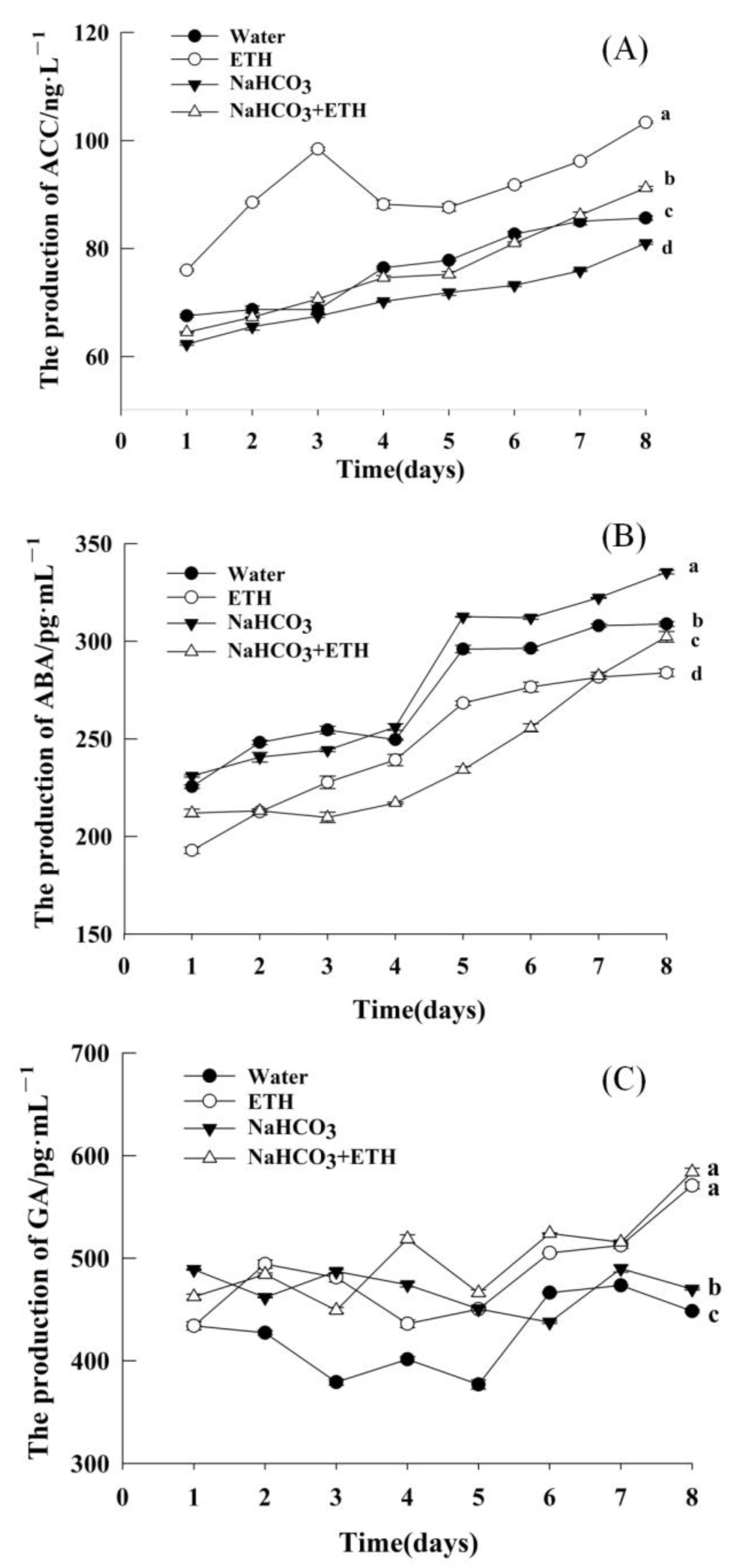
2.2. Changes in Hormone Content of Embryos during Germination in S. pohuashanensis
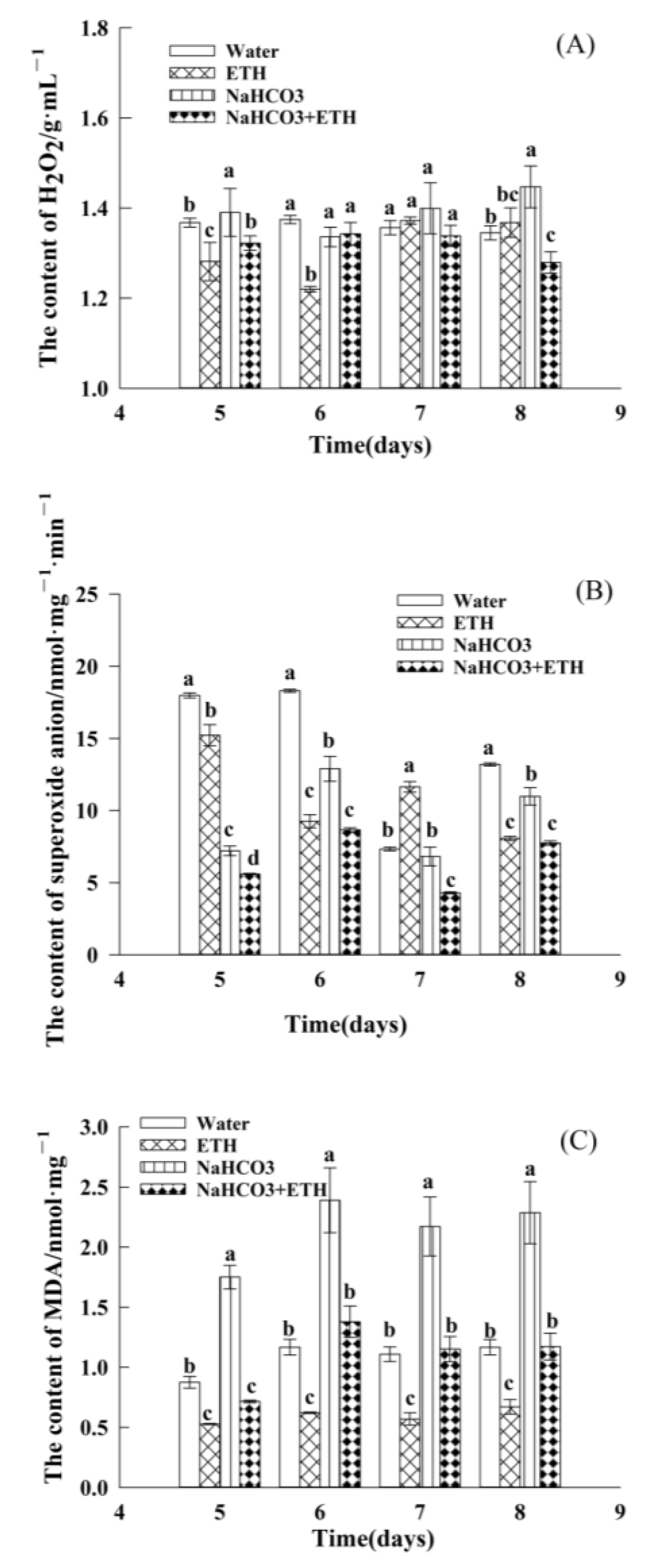
2.3. Changes in ROS during Germination of S. pohuashanensis Embryos
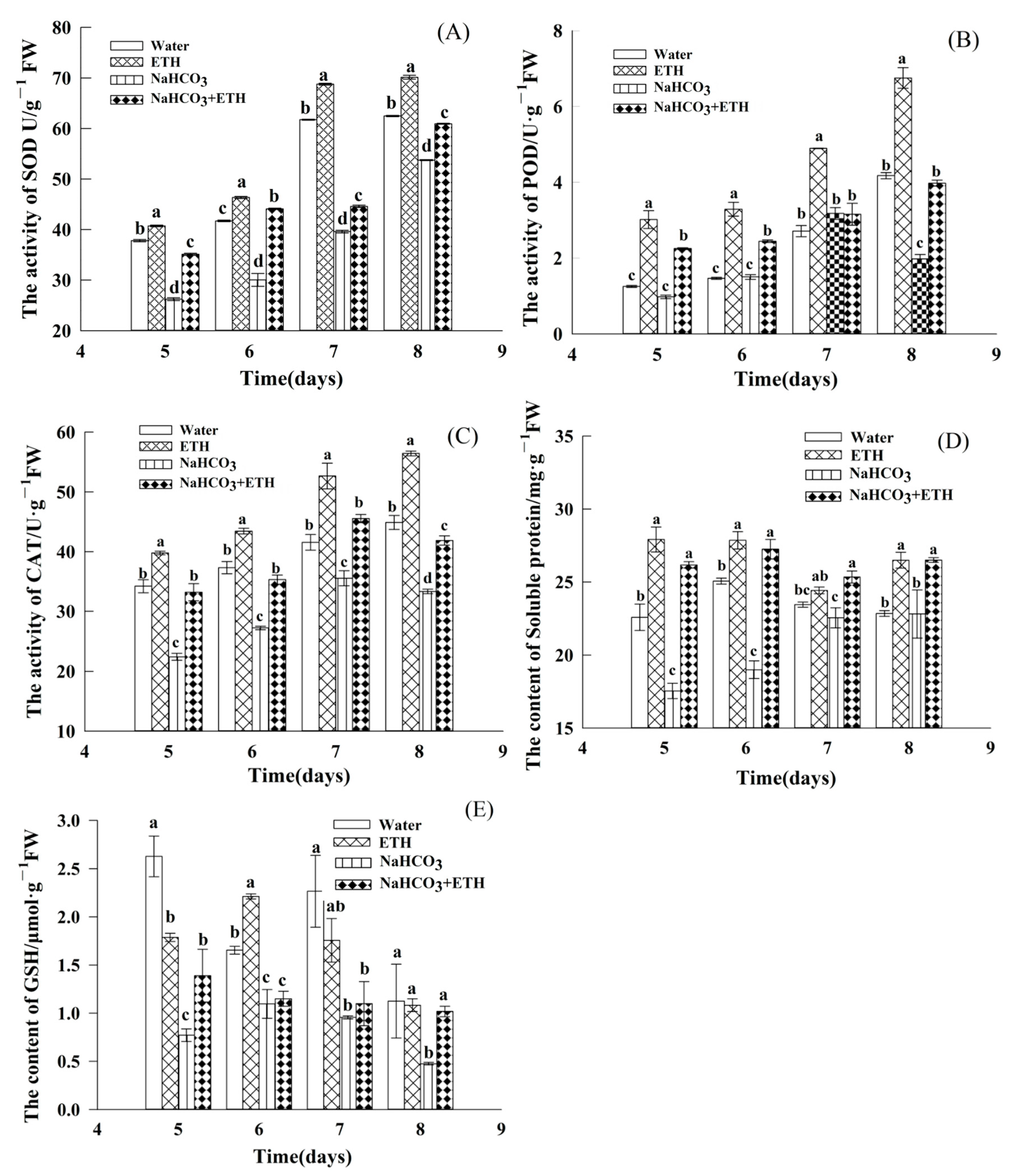
2.4. Changes in Antioxidant Enzyme Activities during Germination of S. pohuashanensis Embryos
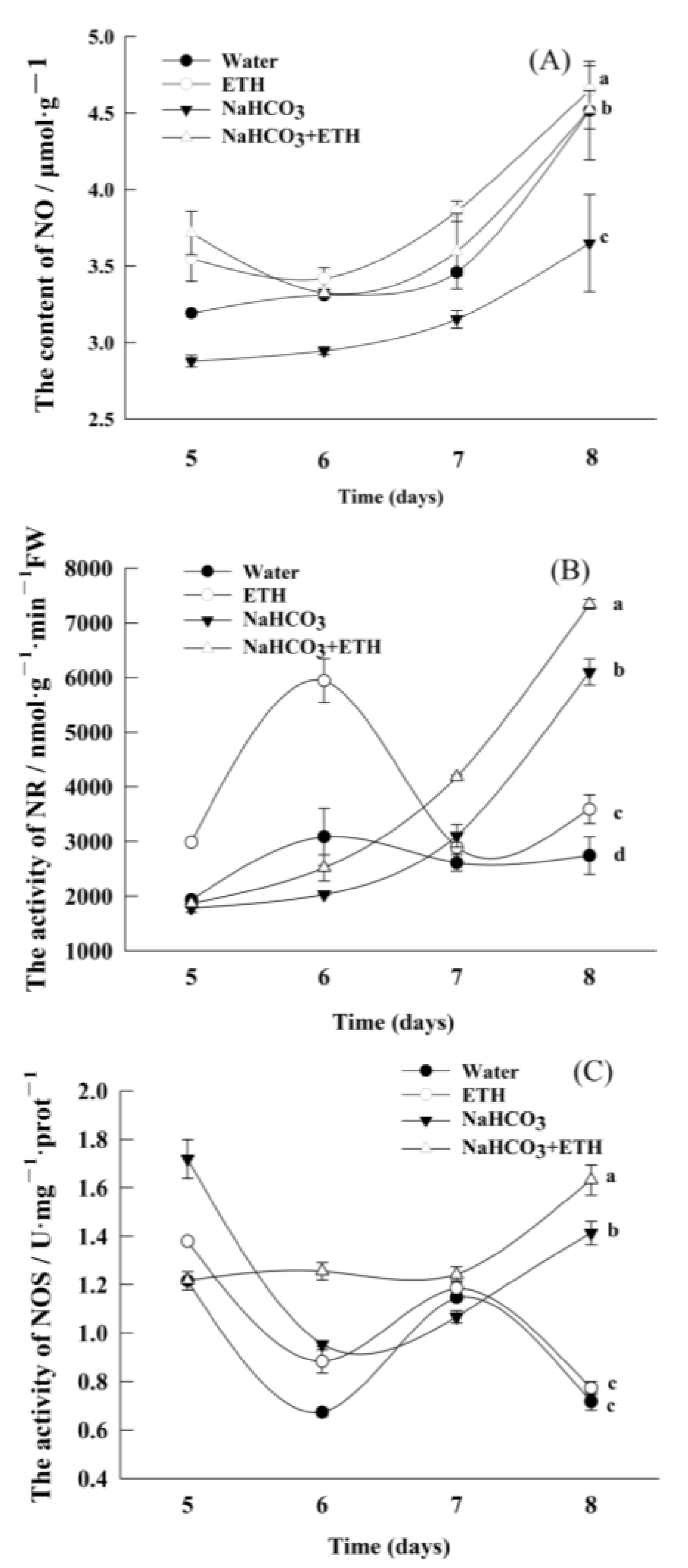
2.5. Changes in Active Nitrogen Content during Germination of S. pohuashanensis Embryos
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. Experimental Methods
4.2.1. Seed Pre-Treatment
4.2.2. Ethephon Treatment
4.2.3. Seed Germination under Salt Stress
4.2.4. Germination Experiments
4.2.5. Endogenous Hormone Assay
4.2.6. ROS Accumulation Assay
4.2.7. Antioxidant Enzyme Activity Assay
4.2.8. Reactive Nitrogen Species Accumulation Assay
4.2.9. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, P.G.; Pan, H.Q.; Boak, E.N.; Pierson, L.S.; Pierson, E.A. Phenazine-producing rhizobacteria promote plant growth and reduce redox and osmotic stress in wheat seedlings under saline conditions. Front. Plant Sci. 2020, 11, 575314. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Song, F.B. Physiological responses and adaptive capacity of oats to saline-alkali stress. Ecol. Environ. 2006, 15, 625–629. [Google Scholar]
- Zhao, Y.Y.; Lu, Z.H.; He, L. Effects of Saline-Alkaline Stress on Seed Germination and Seedling Growth of Sorghum bicolor (L.) Moench. Appl. Biochem. Biotechnol. 2014, 173, 1680–1691. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Yu, J.H.; Liao, W.B.; Xie, J.M.; Niu, L.J.; Zhang, G.B.; Lv, J.; Xiao, X.M.; Wu, Y. Ethylene was Involved in Ca2+-Regulated Na+ Homeostasis.; Na+ Transport and Cell Ultrastructure During Adventitious Rooting in Cucumber Explants Under Salt Stress. J. Plant Biol. 2020, 63, 311–320. [Google Scholar] [CrossRef]
- Zhao, C.Q.; Li, J.W.; Fan, X.F.; Hou, X.C.; Wu, J.Y.; Hu, Y.G.; Liu, J.L. Effects of salt stress on biomass.; quality.; and photosynthetic physiology in switchgrass. Acta Ecol. Sinica. 2015, 35, 6489–6495. [Google Scholar]
- Han, L.; Xiao, C.X.; Xiao, B.B.; Wang, M.; Liu, J.T.; Bhanbhro, N.; Khan, A.; Wang, H.; Wang, H.; Yang, C.W. Proteomic profiling sheds light on alkali tolerance of common wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2019, 138, 58–64. [Google Scholar] [CrossRef]
- Fan, Y.P.; Lu, X.K.; Chen, X.G.; Wang, J.J.; Wang, D.L.; Wang, S.; Guo, L.X.; Rui, C.; Zhang, Y.X.; Cui, R.F. Cotton transcriptome analysis reveals novel biological pathways that eliminate reactive oxygen species (ROS) under sodium bicarbonate (NaHCO3) alkaline stress. Genomics 2021, 113, 1157–1169. [Google Scholar] [CrossRef]
- Wang, B.L.; Zhang, J.L.; Xia, X.Z.; Zhang, W.H. Ameliorative effect of brassinosteroid and ethylene on germination of cucumber seeds in the presence of sodium chloride. Plant Growth Regulation 2011, 65, 407–413. [Google Scholar] [CrossRef]
- Yang, L.; Shen, H.L. Effect of electrostatic field on seed germination and seedling growth of Sorbus pohuashanesis. J. For. Res. 2011, 22, 27–34. [Google Scholar] [CrossRef]
- Yang, L.; Wang, J.A.; Bian, L.; Li, Y.H.; Shen, H.L. Cyclic secondary somatic embryogenesis and efficient plant regeneration in mountain ash (Sorbus pohuashanensis). Plant Cell Tissue Organ Culture 2012, 111, 173–182. [Google Scholar] [CrossRef]
- Yang, L.; Li, Y.H.; Shen, H.L. Somatic embryogenesis and plant regeneration from immature zygotic embryo cultures of mountain ash (Sorbus pohuashanensis). Plant Cell Tissue and Organ Culture 2012, 109, 547–556. [Google Scholar] [CrossRef]
- Xu, J.W.; Shen, H.L.; Zhang, X.L.; Zhang, P.; Huang, J. Sorbus pohuashanensis seed dispersal and germination and their relationships with population natural regeneration. J. Appl. Ecol. 2010, 21, 2536–2544. [Google Scholar]
- Zhang, D.Y.; Wei, C.; Liu, H.N.; Yang, L. Effects of Ethrel and SNP on the release of embryo dormancy in Sorbus pohuashanensis. Seed 2018, 37, 14–17. [Google Scholar]
- Wang, H.; Tang, S.R.; Wang, J.N.; Sheng, H.L.; Yang, L. Interaction between reactive oxygen species and hormones during the breaking of embryo dormancy in Sorbus pohuashanensis by exogenous nitric oxide. J. For. Res. 2022, 33, 435–444. [Google Scholar] [CrossRef]
- Wang, Y.; Diao, P.F.; Kong, L.Q.; Yu, R.N.; Zhang, M.; Zuo, T.T.; Fan, Y.Y.; Niu, Y.D.; Yan, F.; Wuriyanghan, H. Ethylene Enhances Seed Germination and Seedling Growth Under Salinity by Reducing Oxidative Stress and Promoting Chlorophyll ContentviaETR2 Pathway. Front. Plant Sci. 2020, 11, 1066. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Krasuska, U.; Bogatek, R. Dormancy removal in apple embryos by nitric oxide or cyanide involves modifications in ethylene biosynthetic pathway. Planta 2010, 232, 1397–1407. [Google Scholar] [CrossRef]
- Ye, N.H.; Zhu, G.H.; Liu, Y.G.; Zhang, A.Y.; Li, Y.X.; Liu, R.; Shi, L.; Jia, L.G.; Zhang, J.H. Ascorbic acid and reactive oxygen species are involved in the inhibition of seed germination by abscisic acid in rice seeds. J. Exp. Bot. 2012, 63, 1809–1822. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, S.G.; Park, C.M. Salicylic acid promotes seed germination under high salinity by modulating antioxidant activity in Arabidopsis. New Phytologist 2010, 189, 644. [Google Scholar] [CrossRef]
- Lin, Y.C.; Wang, J.J.; Zu, Y.G.; Tang, Z.H. Ethylene antagonizes the inhibition of germination in Arabidopsis induced by salinity by modulating the concentration of hydrogen peroxide. Acta Physiol. Plantarum 2012, 34, 1895–1904. [Google Scholar] [CrossRef]
- James, R.A.; Blake, C.; Byrt, C.S.; Munns, R. Major genes for Na+ exclusion; Nax1 and Nax2 (wheat HKT1;4 and HKT1;5); decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions. J. Exp. Botany 2011, 62, 2939–2947. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Liu, Y.; Zhang, Y.; Zhang, H.; Mu, P.; Lin, Q. Responses of antioxidant properties and root activity of different wheat varieties to salt stress at seedling stage. J. Triticeae Crops 2015, 35, 1168–1175. [Google Scholar]
- Xu, C.; Zhang, Y.P.; Yang, H.B. Effects of ethephon on physiological characteristics and gene expression of Tartary buckwheat under salt stress. Chil. J. Agric. Res. 2022, 82, 234–243. [Google Scholar] [CrossRef]
- Yang, L.; Wang, J.N.; Bian, L.; Shen, H.L. Effects of exogenous NO on the accumulation of reactive oxygen species in embryo germination and early seedling development of Sorbus pohuashanensis. Sci. Silvae Sinice 2013, 49, 60–67. [Google Scholar]
- Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard. J. Plant Physiol. 2015, 178, 84–91. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A.; Khan, M.I.R. A new perspective of phytohormones in salinity tolerance: Regulation of proline metabolism. Environ Exp Bot. 2014, 100, 34–42. [Google Scholar] [CrossRef]
- Nazar, R.; Khan, M.I.R.; Iqbal, N.; Masood, A.; Khan, N.A. Involvement of ethylene in reversal of salt-inhibited photosynthesis by sulfur in mustard. Physiol Plant. 2014, 152, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Mou, W.S.; Kao, Y.T.; Michard; Simon, A.A.; Li, D.; Wudick, M.M.; Lizzio, A.; Feijó, J.A.; Chang, C.R. Ethylene-independent signaling by the ethylene precursor ACC in Arabidopsis ovular pollen tube attraction. Nat. Commun. 2020, 11, 4082. [Google Scholar] [CrossRef]
- Ahmad, I.; Zhu, G.L.; Zhou, G.S.; Song, X.D.; Ibrahim, M.E.H.; Salih, E.G.I.; Hussain, S.; Younas, M.U. Pivotal Role of Phytohormones and Their Responsive Genes in Plant Growth and Their Signaling and Transduction Pathway under Salt Stress in Cotton. Int. J. Mol. Sci. 2022, 23, 7339. [Google Scholar] [CrossRef]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Fornt. Plant Sci. 2013, 4, 63. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.C.; Wang, Q.Y.; Gao, Z.W.; Wang, Y.; Liu, Y.J.; Ma, Z.P.; Chen, Y.W.; Zhang, Y.C.; Yan, F. Analysis of Phytohormone Signal Transduction in Sophora alopecuroides under Salt Stress. Int. J. Mol. Sci. 2021, 22, 7313. [Google Scholar] [CrossRef]
- Yin, D.C.; Halifu, S.; Song, R.Q.; Qi, J.Y.; Deng, X.; Deng, J.F. Effects of an ectomycorrhizal fungus on the growth and physiology of Pinus sylvestris var. mongolica seedlings subjected to saline–alkali stress. J. For. Res. 2020, 31, 781–788. [Google Scholar] [CrossRef]
- Katsuhara, M.; Kawasaki, T. Salt stress induced nuclear and DNA degradation in meristematic cells of barley roots. Plant Cell Physiol. 1996, 37, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Teh, C.Y.; Mahmood, M.; Shaharuddin, N.A.; Ho, C.L. In vitro rice shoot apices as simple model to study the effect of NaCl and the potential of exogenous proline and glutathione in mitigating salinity stress. Plant Growth Regul. 2014, 75, 1–11. [Google Scholar] [CrossRef]
- Ozt€urk, L.; Demir, Y. Effects of putrescine and ethephon on some oxidative stress enzyme activities and proline content in salt stressed spinach leaves. Plant Growth Regul. 2003, 40, 89–95. [Google Scholar] [CrossRef]
- Sehar, Z.; Iqbal, N.; Khan, M.I.R.; Masood, A.; Rehman, M.T.; Hussain, A.; AlAjmi, M.F.; Ahmad, A.; Khan, N.A. Ethylene reduces glucose sensitivity and reverses photosynthetic repression through optimization of glutathione production in salt-stressed wheat (Triticum aestivum L.). Sci. Rep. 2021, 11, 1. [Google Scholar] [CrossRef]
- Phartyal, S.S.; Rosbakh, S.; Gruber, M.; Poschlod, P. The sweet and musky scent of home: Biogenic ethylene fine-tunes seed germination in wetlands. Plant Biol. 2022, 24, 278–285. [Google Scholar] [CrossRef]
- Liu, Y.G.; Shi, L.; Ye, N.H.; Liu, R.; Jia, W.S.; Zhang, J.H. Nitric oxide-induced rapid decrease of abscisic acid concentration is required in breaking seed dormancy in Arabidopsis. New Phytol. 2009, 183, 1030–1042. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Dobrzynska, U.; Babanczyk, T.; Bogatek, R. Breaking the apple embryo dormancy by nitric oxide involves the stimulation of ethylene production. Planta 2007, 225, 1051–1057. [Google Scholar] [CrossRef]
- García, M.J.; Suárez, V.; Romera, F.J.; Alcántara, E.; Pérez-Vicente, R. A new model involving ethylene.; nitric oxide and Fe to explain the regulation of Fe-acquisition genes in strategy I plants. Plant Physiol. Biochem. 2011, 49, 537–544. [Google Scholar] [CrossRef]
- Bian, L.; Yang, L.; Wang, J.A.; Shen, H.L. Effects of KNO3 pretreatment and temperature on seed germination of Sorbus pohuashanensis. J. For. Res. 2013, 24, 309–316. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Water | ETH | NaHCO3 | NaHCO3 + ETH |
|---|---|---|---|---|
| Germination percentage (%) | 43.33 ± 3.95 b | 60.00 ± 3.85 a | 6.67 ± 1.92 d | 18.89 ± 4.01 c |
| Mean germination speed | 6.25 ± 0.16 b | 4.98 ± 0.18 c | 7.17 ± 0.44 a | 6.68 ± 0.09 ab |
| Germination index | 2.19 ± 0.24 b | 3.84 ± 0.10 a | 0.29 ± 0.09 d | 0.87 ± 0.18 c |
| Germination potential (%) | 14.44 ± 1.11 b | 24.44 ± 1.11 a | 0.29 ± 0.10 c | 6.67 ± 1.92 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhao, C.; Wang, X.; Shen, H.; Yang, L. Exogenous Ethylene Alleviates the Inhibition of Sorbus pohuashanensis Embryo Germination in a Saline-Alkali Environment (NaHCO3). Int. J. Mol. Sci. 2023, 24, 4244. https://doi.org/10.3390/ijms24044244
Wang Y, Zhao C, Wang X, Shen H, Yang L. Exogenous Ethylene Alleviates the Inhibition of Sorbus pohuashanensis Embryo Germination in a Saline-Alkali Environment (NaHCO3). International Journal of Molecular Sciences. 2023; 24(4):4244. https://doi.org/10.3390/ijms24044244
Chicago/Turabian StyleWang, Yutong, Caihong Zhao, Xiaodong Wang, Hailong Shen, and Ling Yang. 2023. "Exogenous Ethylene Alleviates the Inhibition of Sorbus pohuashanensis Embryo Germination in a Saline-Alkali Environment (NaHCO3)" International Journal of Molecular Sciences 24, no. 4: 4244. https://doi.org/10.3390/ijms24044244
APA StyleWang, Y., Zhao, C., Wang, X., Shen, H., & Yang, L. (2023). Exogenous Ethylene Alleviates the Inhibition of Sorbus pohuashanensis Embryo Germination in a Saline-Alkali Environment (NaHCO3). International Journal of Molecular Sciences, 24(4), 4244. https://doi.org/10.3390/ijms24044244