
Nuclear High Mobility Group A2 (HMGA2) Interactome Revealed by Biotin Proximity Labeling

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
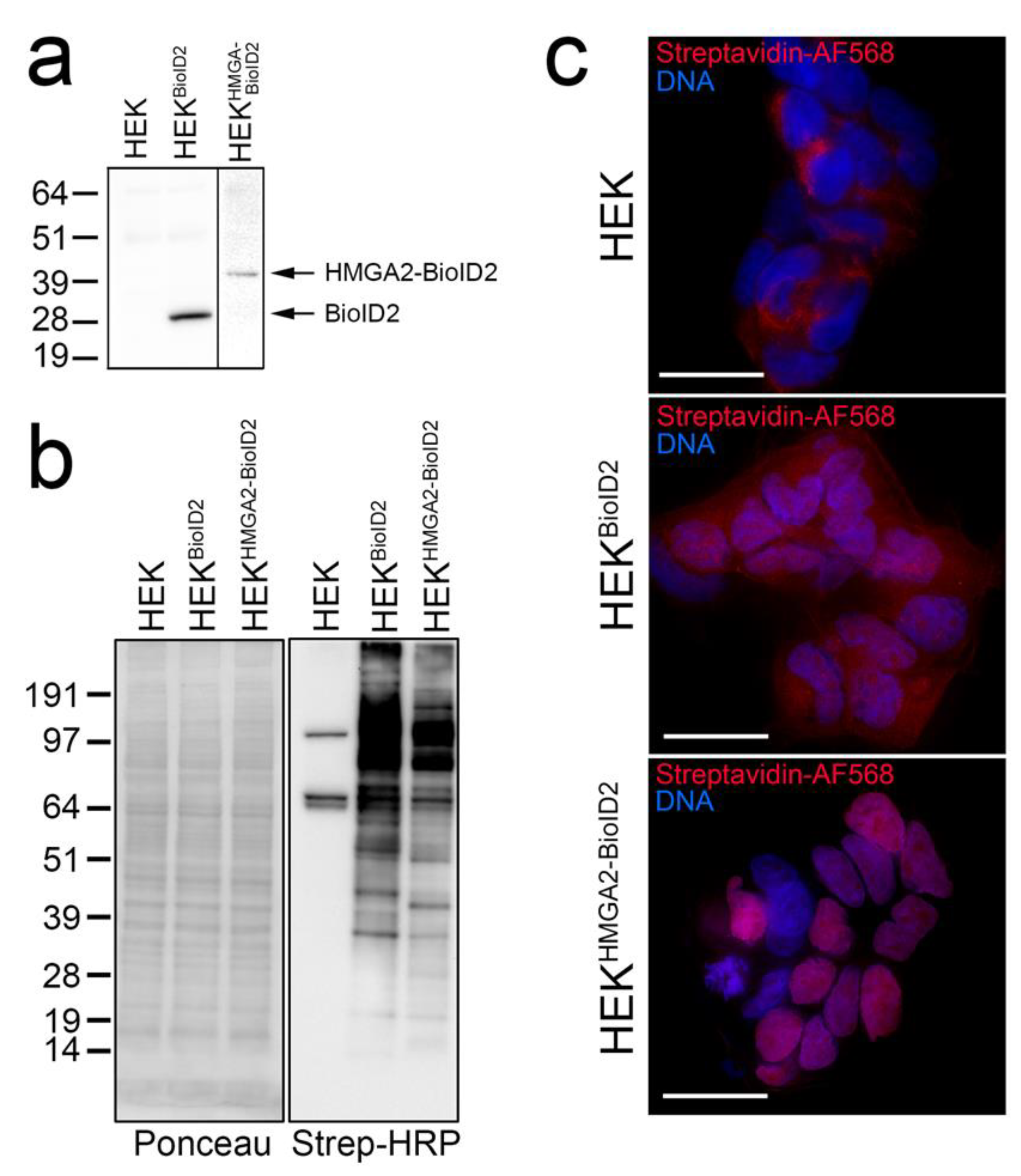
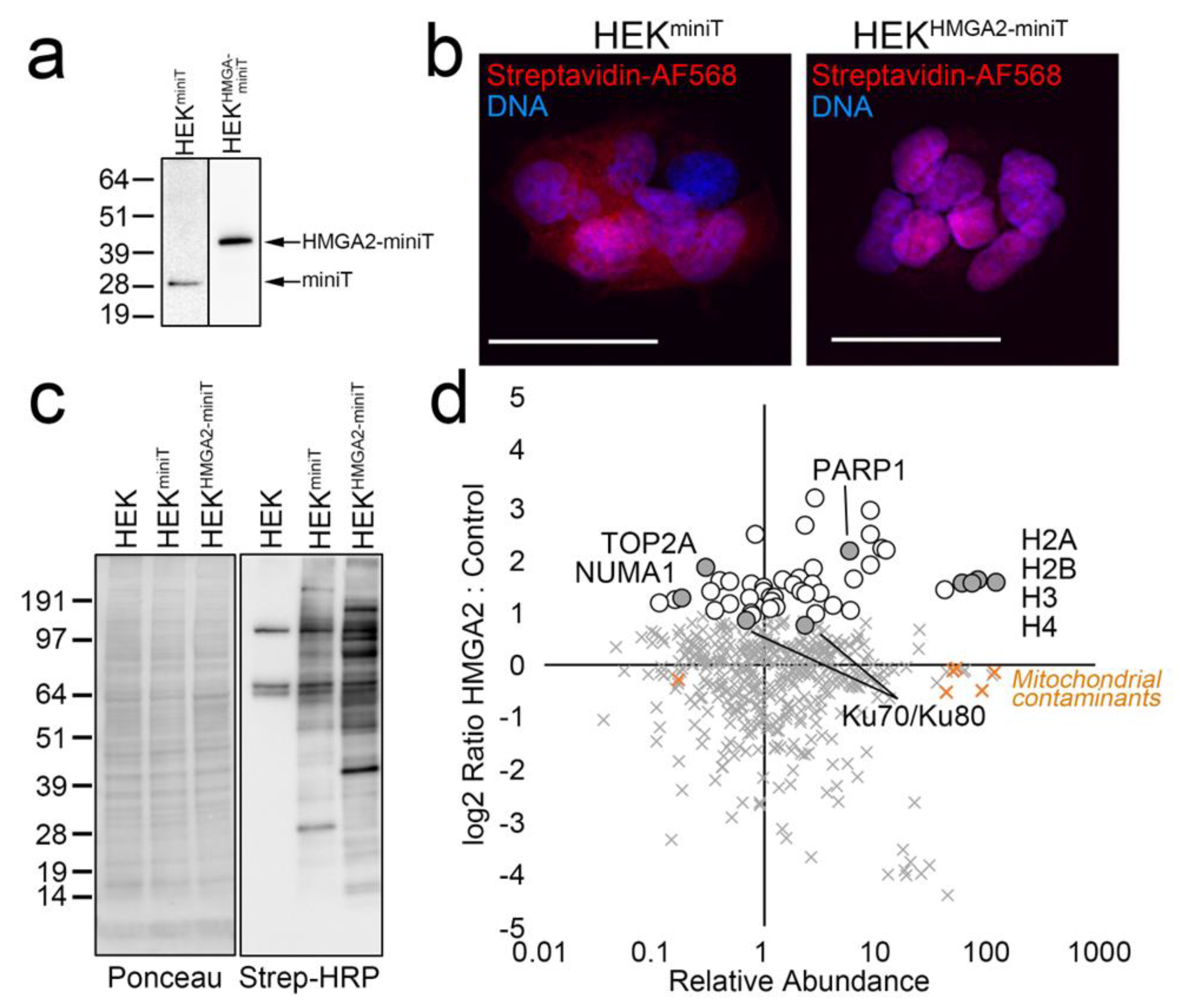
2.1. HMGA2-BioID2 Fusion Protein Biotinylates Nuclear Proteins
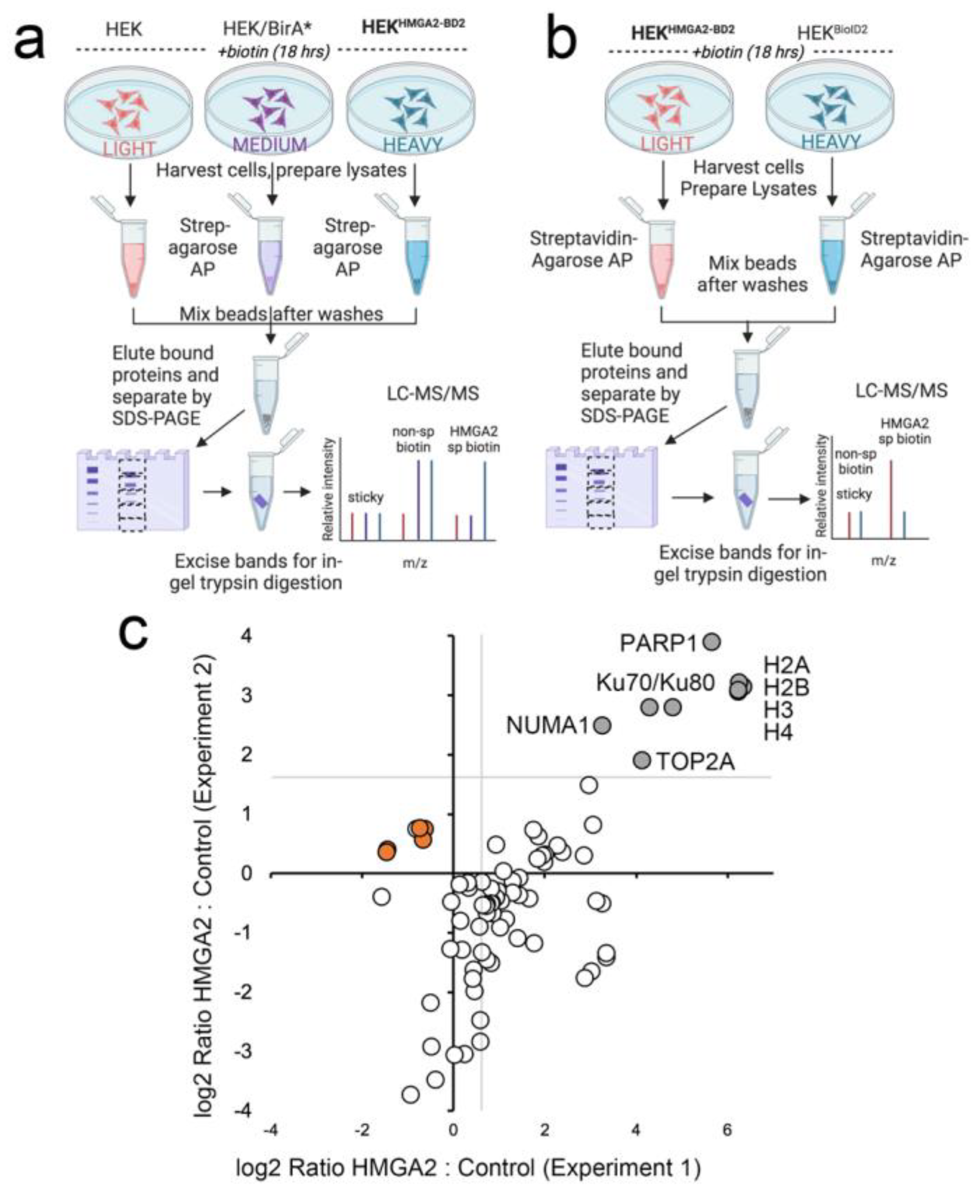
2.2. HMGA2 Proximity Labeling Identifies A Unique Nuclear HMGA2 Interactome
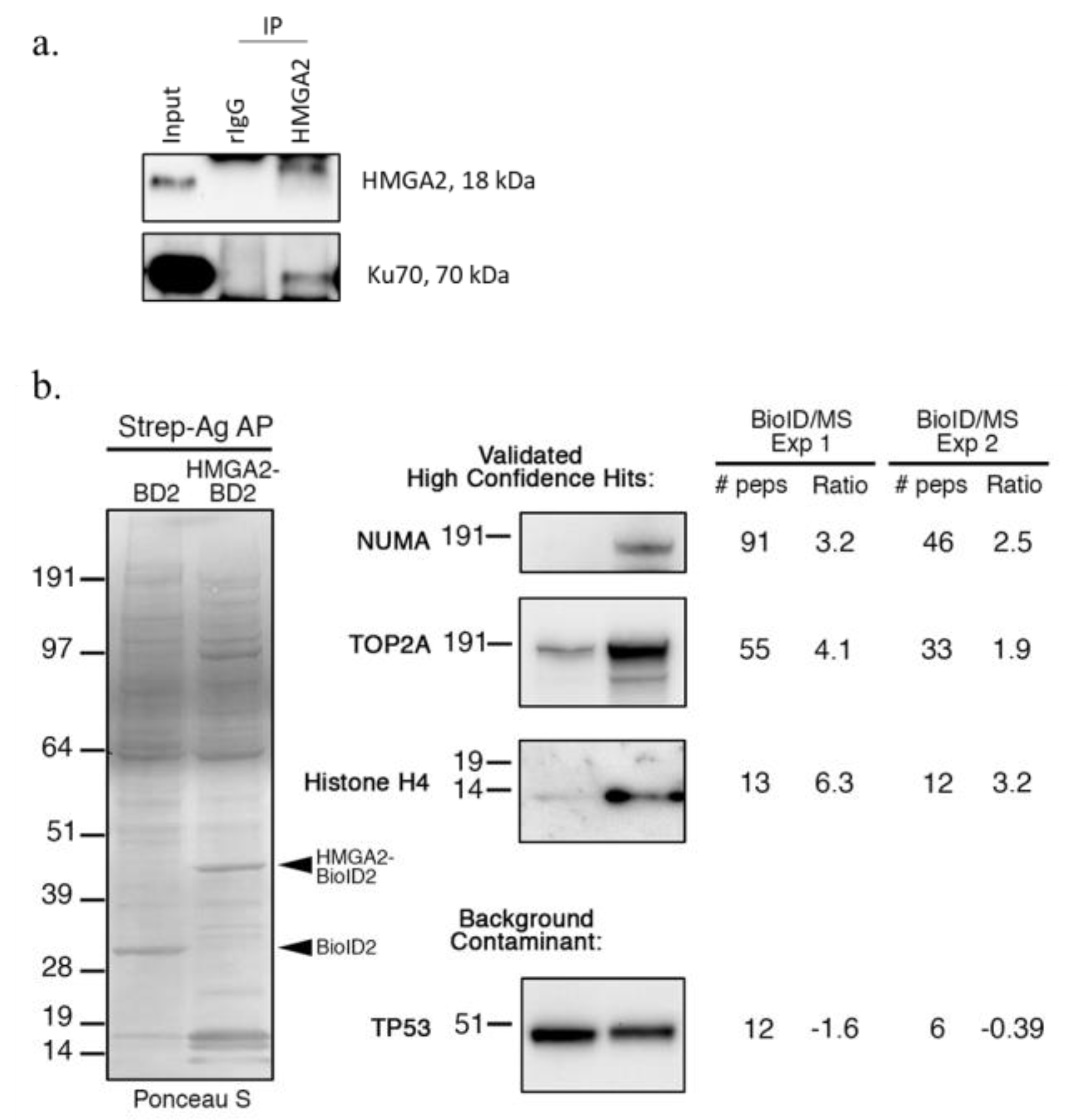
2.3. Nuclear HMGA2-BioID2 Interactome Confirms Known and Discovers New Interaction Partners
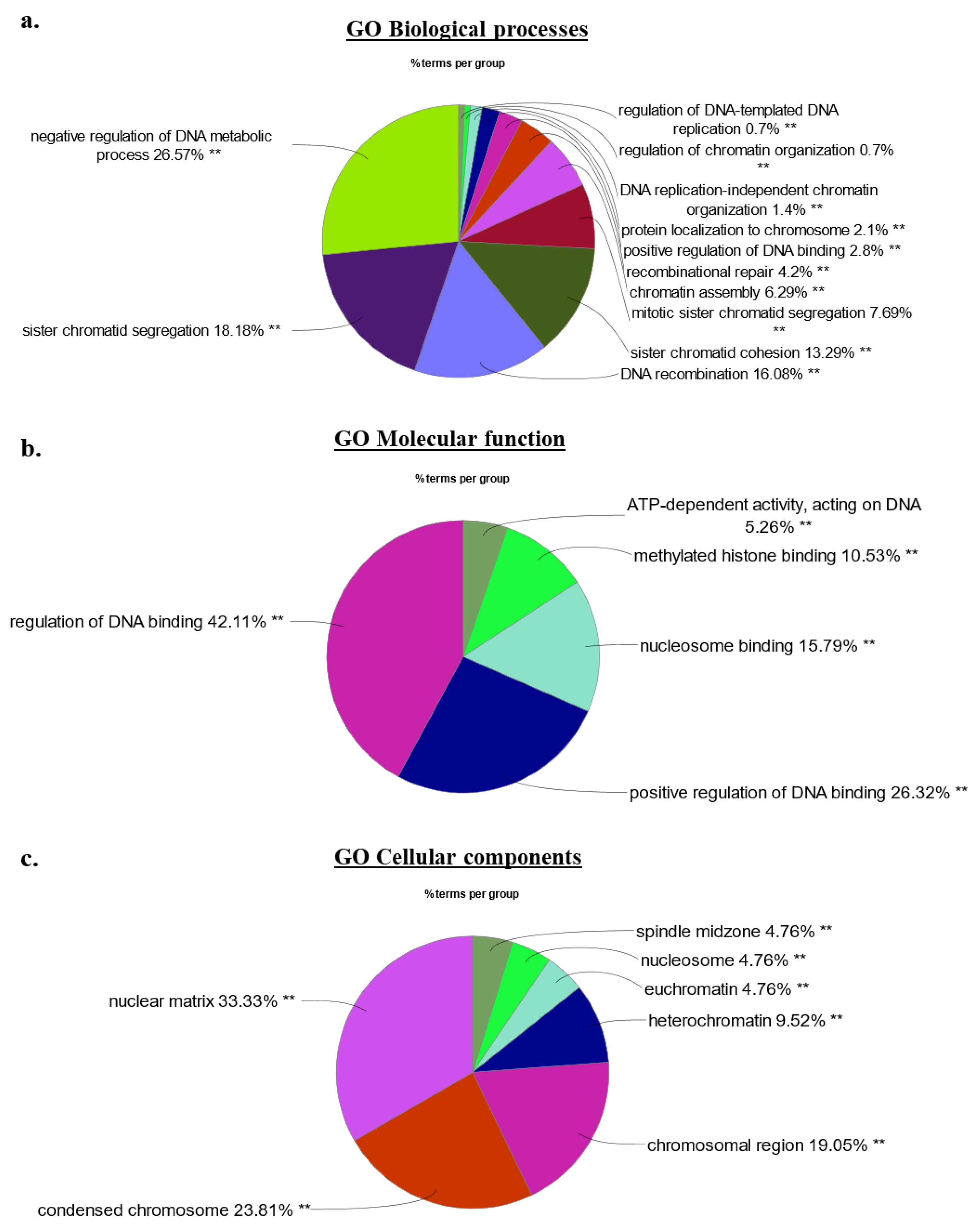
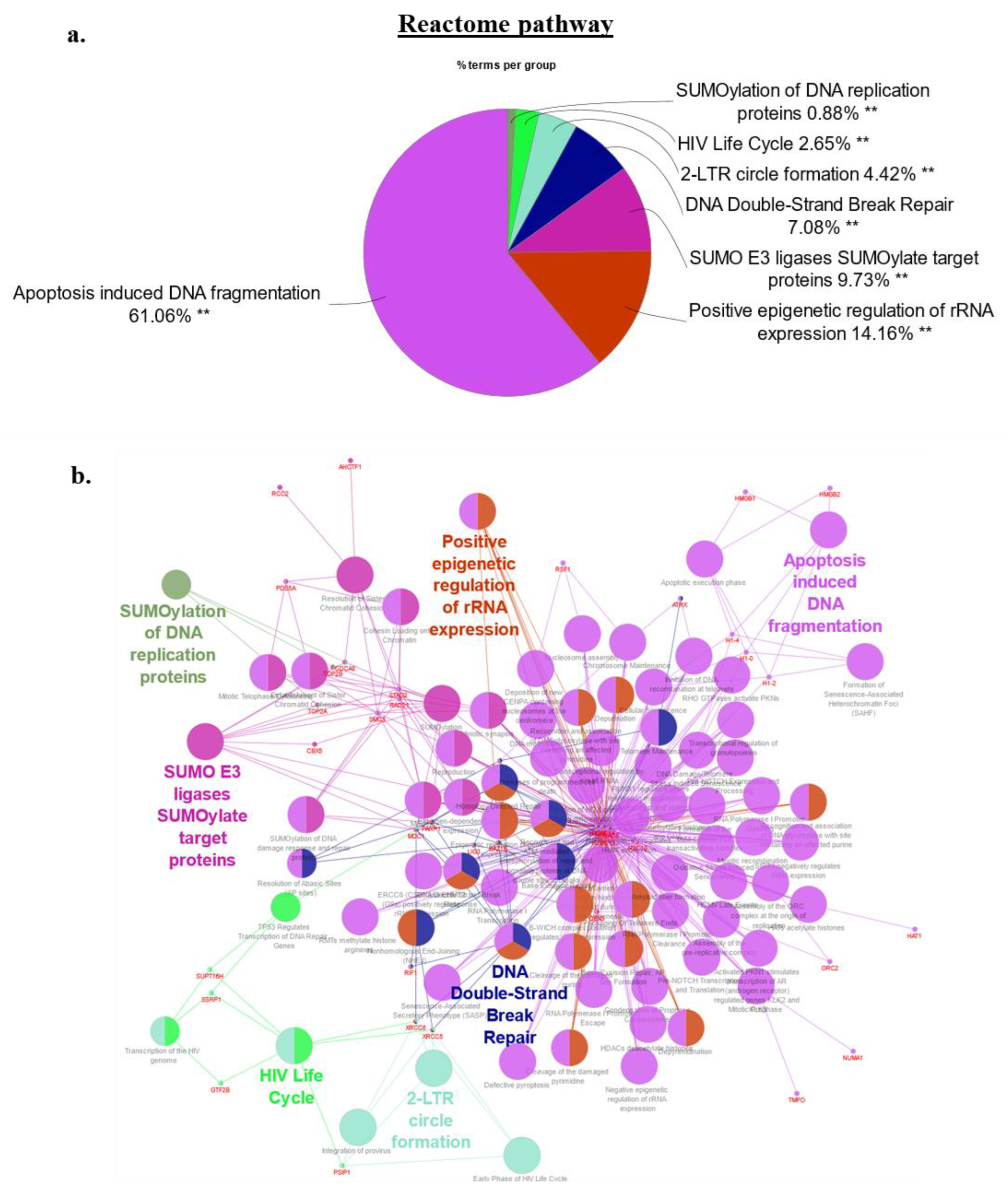
2.4. Bioinformatics Signature of the HMGA2 Proximity Interactome
3. Discussion
4. Materials and Methods
4.1. Creation of Stable HEK293 Cell Lines Expressing BioID2 and Miniturbo Fusion Proteins
4.2. Metabolic Labeling and Liquid Chromatography Mass Spectrometry (LC-MS/MS) Analysis
4.3. Western Blot Validation by Co-Immunoprecipitation (IP) or BioID
4.4. Bioinformatics Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gattas, G.J.; Quade, B.J.; Nowak, R.A.; Morton, C.C. HMGIC expression in human adult and fetal tissues and in uterine leiomyomata. Genes Chromosomes Cancer 1999, 25, 316–322. [Google Scholar] [CrossRef]
- Droge, P.; Davey, C.A. Do cells let-7 determine stemness? Cell Stem Cell 2008, 2, 8–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cello, F.; Hillion, J.; Hristov, A.; Wood, L.; Mukherjee, M.; Schuldenfrei, A.; Kowalski, J.; Bhattacharya, R.; Ashfaq, R.; Resar, L. HMGA2 participates in transformation in human lung cancer. Mol. Cancer Res. 2008, 6, 743–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedele, M.; Fusco, A. HMGA and cancer. Biochim. Biophys. Acta 2010, 1799, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Fusco, A.; Fedele, M. Roles of HMGA proteins in cancer. Nat. Rev. Cancer 2007, 7, 899–910. [Google Scholar] [CrossRef]
- Wood, L.; Maher, J.; Bunton, T.; Resar, L. The oncogenic properties of the HMG-I gene family. Cancer Res. 2000, 60, 4256–4261. [Google Scholar]
- Hristov, A.; Cope, L.; Reyes, M.; Singh, M.; Iacobuzio-Donahue, C.; Maitra, A.; Resar, L. HMGA2 protein expression correlates with lymph node metastasis and increased tumor grade in pancreatic ductal adenocarcinoma. Mod. Pathol. 2009, 22, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Morishita, A.; Zaidi, M.; Mitoro, A.; Sankarasharma, D.; Szabolcs, M.; Okada, Y.; D’Armiento, J.; Chada, K. HMGA2 is a driver of tumor metastasis. Cancer Res. 2013, 73, 4289–4299. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, X.; Li, A.; Chen, L.; Lai, L.; Lin, H.; Hu, S.; Yao, L.; Peng, J.; Loera, S.; et al. Overexpression of HMGA2 promotes metastasis and impacts survival of colorectal cancers. Clin. Cancer Res. 2011, 17, 2570–2580. [Google Scholar] [CrossRef] [Green Version]
- Bansod, S.; Kageyama, R.; Ohtsuka, T. Hes5 regulates the transition timing of neurogenesis and gliogenesis in mammalian neocortical development. Development 2017, 144, 3156–3167. [Google Scholar] [CrossRef] [Green Version]
- Parisi, S.; Piscitelli, S.; Passaro, F.; Russo, T. HMGA Proteins in Stemness and Differentiation of Embryonic and Adult Stem Cells. Int. J. Mol. Sci. 2020, 21, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.; Mou, Y.; Shi, X.; Liu, T.; Chen, Z.; Zuo, X. Circular RNA 100146 Promotes Colorectal Cancer Progression by the MicroRNA 149/HMGA2 Axis. Mol. Cell Biol. 2021, 41, e00445-20. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Lv, J.; Jiang, T.; Li, B.; Li, Y.; He, Z.; Xuan, Z.; Sun, G.; Wang, S.; Li, Z.; et al. CircFAM73A promotes the cancer stem cell-like properties of gastric cancer through the miR-490-3p/HMGA2 positive feedback loop and HNRNPK-mediated beta-catenin stabilization. J. Exp. Clin. Cancer Res. 2021, 40, 103. [Google Scholar] [CrossRef]
- Ma, W.; Xue, N.; Zhang, J.; Wang, D.; Yao, X.; Lin, L.; Xu, Q. circUBAP2 regulates osteosarcoma progression via the miR-204-3p/HMGA2 axis. Int. J. Oncol. 2021, 58, 298–311. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Mateos, C.; Sanchez-Castillo, A.; Soler, M.; Obiols-Guardia, A.; Pineyro, D.; Boque-Sastre, R.; Calleja-Cervantes, M.; de Moura, M.C.; Martinez-Cardus, A.; Rubio, T.; et al. The transcribed pseudogene RPSAP52 enhances the oncofetal HMGA2-IGF2BP2-RAS axis through LIN28B-dependent and independent let-7 inhibition. Nat. Commun. 2019, 10, 3979. [Google Scholar] [CrossRef] [Green Version]
- Young, A.; Narita, M. Oncogenic HMGA2: Short or small? Genes. Dev. 2007, 21, 1005–1009. [Google Scholar] [CrossRef] [Green Version]
- Copley, M.; Babovic, S.; Benz, C.; Knapp, D.; Beer, P.; Kent, D.; Wohrer, S.; Treloar, D.; Day, C.; Rowe, K.; et al. The Lin28b-let-7-Hmga2 axis determines the higher self-renewal potential of fetal haematopoietic stem cells. Nat. Cell Biol. 2013, 15, 916–925. [Google Scholar] [CrossRef]
- Navarra, A.; Musto, A.; Gargiulo, A.; Petrosino, G.; Pierantoni, G.; Fusco, A.; Russo, T.; Parisi, S. Hmga2 is necessary for Otx2-dependent exit of embryonic stem cells from the pluripotent ground state. BMC Biol. 2016, 14, 24. [Google Scholar] [CrossRef] [Green Version]
- Acampora, D.; Omodei, D.; Petrosino, G.; Garofalo, A.; Savarese, M.; Nigro, V.; Di Giovannantonio, L.; Mercadante, V.; Simeone, A. Loss of the Otx2-Binding Site in the Nanog Promoter Affects the Integrity of Embryonic Stem Cell Subtypes and Specification of Inner Cell Mass-Derived Epiblast. Cell Rep. 2016, 15, 2651–2664. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, T.; Saito, K.; Higurashi, M.; Ishikawa, F.; Kohno, Y.; Mori, K.; Shibanuma, M. HMGA2 drives the IGFBP1/AKT pathway to counteract the increase in P27KIP1 protein levels in mtDNA/RNA-less cancer cells. Cancer Sci. 2022, 114, 152–163. [Google Scholar] [CrossRef]
- Busch, B.; Bley, N.; Muller, S.; Glass, M.; Misiak, D.; Lederer, M.; Vetter, M.; Strauss, H.; Thomssen, C.; Huttelmaier, S. The oncogenic triangle of HMGA2, LIN28B and IGF2BP1 antagonizes tumor-suppressive actions of the let-7 family. Nucleic Acids Res. 2016, 44, 3845–3864. [Google Scholar] [CrossRef] [PubMed]
- Wend, P.; Runke, S.; Wend, K.; Anchondo, B.; Yesayan, M.; Jardon, M.; Hardie, N.; Loddenkemper, C.; Ulasov, I.; Lesniak, M.; et al. WNT10B/beta-catenin signalling induces HMGA2 and proliferation in metastatic triple-negative breast cancer. EMBO Mol. Med. 2013, 5, 264–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, L.; Zhang, J.; Tang, W.; Zhang, N.; He, M.; Guo, Y.; Wang, Z. HMGA2 elicits EMT by activating the Wnt/beta-catenin pathway in gastric cancer. Dig. Dis. Sci. 2013, 58, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Hawsawi, O.; Henderson, V.; Burton, L.; Dougan, J.; Nagappan, P.; Odero-Marah, V. High mobility group A2 (HMGA2) promotes EMT via MAPK pathway in prostate cancer. Biochem. Biophys. Res. Commun. 2018, 504, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Cattaruzzi, G.; Altamura, S.; Tessari, M.; Rustighi, A.; Giancotti, V.; Pucillo, C.; Manfioletti, G. The second AT-hook of the architectural transcription factor HMGA2 is determinant for nuclear localization and function. Nucleic Acids Res. 2007, 35, 1751–1760. [Google Scholar] [CrossRef] [Green Version]
- Pfannkuche, K.; Summer, H.; Li, O.; Hescheler, J.; Droge, P. The high mobility group protein HMGA2: A co-regulator of chromatin structure and pluripotency in stem cells? Stem. Cell Rev. 2009, 5, 224–230. [Google Scholar] [CrossRef]
- Summer, H.; Li, O.; Bao, Q.; Zhan, L.; Peter, S.; Sathiyanathan, P.; Henderson, D.; Klonisch, T.; Goodman, S.; Droge, P. HMGA2 exhibits dRP/AP site cleavage activity and protects cancer cells from DNA-damage-induced cytotoxicity during chemotherapy. Nucleic Acids Res. 2009, 37, 4371–4384. [Google Scholar] [CrossRef]
- Reeves, R.; Nissen, M. The A.T-DNA-binding domain of mammalian high mobility group I chromosomal proteins. A novel peptide motif for recognizing DNA structure. J. Biol. Chem. 1990, 265, 8573–8582. [Google Scholar] [CrossRef]
- Sgarra, R.; Maurizio, E.; Zammitti, S.; Sardo, A.L.; Giancotti, V.; Manfioletti, G. Macroscopic differences in HMGA oncoproteins post-translational modifications: C-terminal phosphorylation of HMGA2 affects its DNA binding properties. J. Proteome. Res. 2009, 8, 2978–2989. [Google Scholar] [CrossRef]
- Noro, B.; Licheri, B.; Sgarra, R.; Rustighi, A.; Tessari, M.; Chau, K.; Ono, S.; Giancotti, V.; Manfioletti, G. Molecular dissection of the architectural transcription factor HMGA2. Biochemistry 2003, 42, 4569–4577. [Google Scholar] [CrossRef]
- Xu, J.; Fang, X.; Long, L.; Wang, S.; Qian, S.; Lyu, J. HMGA2 promotes breast cancer metastasis by modulating Hippo-YAP signaling pathway. Cancer Biol. Ther. 2021, 22, 5–11. [Google Scholar] [CrossRef]
- Fujikane, R.; Komori, K.; Sekiguchi, M.; Hidaka, M. Function of high-mobility group A proteins in the DNA damage signaling for the induction of apoptosis. Sci. Rep. 2016, 6, 31714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobersch, S.; Rubio, K.; Singh, I.; Gunther, S.; Graumann, J.; Cordero, J.; Castillo-Negrete, R.; Huynh, M.; Mehta, A.; Braubach, P.; et al. Positioning of nucleosomes containing gamma-H2AX precedes active DNA demethylation and transcription initiation. Nat. Commun. 2021, 12, 1072. [Google Scholar] [CrossRef] [PubMed]
- Fedele, M.; Visone, R.; De Martino, I.; Troncone, G.; Palmieri, D.; Battista, S.; Ciarmiello, A.; Pallante, P.; Arra, C.; Melillo, R.; et al. HMGA2 induces pituitary tumorigenesis by enhancing E2F1 activity. Cancer Cell 2006, 9, 459–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedele, M.; Pierantoni, G.; Visone, R.; Fusco, A. E2F1 activation is responsible for pituitary adenomas induced by HMGA2 gene overexpression. Cell Div. 2006, 1, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedele, M.; Pierantoni, G.; Visone, R.; Fusco, A. Critical role of the HMGA2 gene in pituitary adenomas. Cell Cycle 2006, 5, 2045–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, D.; Valentino, T.; D’Angelo, D.; De Martino, I.; Postiglione, I.; Pacelli, R.; Croce, C.; Fedele, M.; Fusco, A. HMGA proteins promote ATM expression and enhance cancer cell resistance to genotoxic agents. Oncogene 2011, 30, 3024–3035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SNatarajan; Hombach-Klonisch, S.; Droge, P.; Klonisch, T. HMGA2 inhibits apoptosis through interaction with ATR-CHK1 signaling complex in human cancer cells. Neoplasia 2013, 15, 263–280. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, S.; Begum, F.; Gim, J.; Wark, L.; Henderson, D.; Davie, J.; Hombach-Klonisch, S.; Klonisch, T. High Mobility Group A2 protects cancer cells against telomere dysfunction. Oncotarget 2016, 7, 12761–12782. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Lim, H.; Tjokro, N.; Sathiyanathan, P.; Natarajan, S.; Chew, T.; Klonisch, T.; Goodman, S.; Surana, U.; Droge, P. Chaperoning HMGA2 protein protects stalled replication forks in stem and cancer cells. Cell Rep. 2014, 6, 684–697. [Google Scholar] [CrossRef] [Green Version]
- Hombach-Klonisch, S.; Kalantari, F.; Medapati, M.; Natarajan, S.; Krishnan, S.; Kumar-Kanojia, A.; Thanasupawat, T.; Begum, F.; Xu, F.; Hatch, G.; et al. HMGA2 as a functional antagonist of PARP1 inhibitors in tumor cells. Mol. Oncol. 2019, 13, 153–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinkle-Mulcahy, L. Recent advances in proximity-based labeling methods for interactome mapping. F1000Research 2019, 8, F1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, K.; Kim, D.; Raida, M.; Burke, B. A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J. Cell Biol. 2012, 196, 801–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branon, T.; Bosch, J.; Sanchez, A.; Udeshi, N.; Svinkina, T.; Carr, S.; Feldman, J.; Perrimon, N.; Ting, A. Efficient proximity labeling in living cells and organisms with TurboID. Nat. Biotechnol. 2018, 36, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Kubitz, L.; Bitsch, S.; Zhao, X.; Schmitt, K.; Deweid, L.; Roehrig, A.; Barazzone, E.; Valerius, O.; Kolmar, H.; Bethune, J. Engineering of ultraID, a compact and hyperactive enzyme for proximity-dependent biotinylation in living cells. Commun. Biol. 2022, 5, 657. [Google Scholar] [CrossRef]
- Kim, D.; Jensen, S.; Noble, K.; Kc, B.; Roux, K.; Motamedchaboki, K.; Roux, K. An improved smaller biotin ligase for BioID proximity labeling. Mol. Biol. Cell 2016, 27, 1188–1196. [Google Scholar] [CrossRef]
- Lambert, J.; Tucholska, M.; Go, C.; Knight, J.; Gingras, A. Proximity biotinylation and affinity purification are complementary approaches for the interactome mapping of chromatin-associated protein complexes. J. Proteom. 2015, 118, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Sgarra, R.; Rustighi, A.; Tessari, M.; Di Bernardo, J.; Altamura, S.; Fusco, A.; Manfioletti, G.; Giancotti, V. Nuclear phosphoproteins HMGA and their relationship with chromatin structure and cancer. FEBS Lett. 2004, 574, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.; Wang, J.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome. Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Otasek, D.; Morris, J.; Boucas, J.; Pico, A.; Demchak, B. Cytoscape Automation: Empowering workflow-based network analysis. Genome. Biol. 2019, 20, 185. [Google Scholar] [CrossRef] [Green Version]
- Stabell, M.; Saether, T.; Rohr, A.; Gabrielsen, O.; Myklebost, O. Methylation-dependent SUMOylation of the architectural transcription factor HMGA2. Biochem. Biophys. Res. Commun. 2021, 552, 91–97. [Google Scholar] [CrossRef]
- Cao, X.; Clavijo, C.; Li, X.; Lin, H.; Chen, Y.; Shih, H.; Ann, D. SUMOylation of HMGA2: Selective destabilization of promyelocytic leukemia protein via proteasome. Mol. Cancer Ther. 2008, 7, 923–934. [Google Scholar] [CrossRef] [Green Version]
- Sears, R.; May, D.; Roux, K. BioID as a Tool for Protein-Proximity Labeling in Living Cells. Methods Mol. Biol. 2019, 2012, 299–313. [Google Scholar] [PubMed]
- Krahn, N.; Meier, M.; To, V.; Booy, E.; McEleney, K.; O’Neil, J.D.; McKenna, S.; Patel, T.; Stetefeld, J. Nanoscale Assembly of High-Mobility Group AT-Hook 2 Protein with DNA Replication Fork. Biophys. J. 2017, 113, 2609–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, L.; Baez, M.; Harrilal, C.; Garabedian, A.; Fernandez-Lima, F.; Leng, F. The Dimerization State of the Mammalian High Mobility Group Protein AT-Hook 2 (HMGA2). PLoS ONE 2015, 10, e0130478. [Google Scholar] [CrossRef]
- Malini, E.; Maurizio, E.; Bembich, S.; Sgarra, R.; Edomi, P.; Manfioletti, G. HMGA Interactome: New insights from phage display technology. Biochemistry 2011, 50, 3462–3468. [Google Scholar] [CrossRef] [PubMed]
- Fell, V.; Schild-Poulter, C. The Ku heterodimer: Function in DNA repair and beyond. Mutat. Res. Rev. Mutat. Res. 2015, 763, 15–29. [Google Scholar] [CrossRef]
- Roberts, S.; Strande, N.; Burkhalter, M.; Strom, C.; Havener, J.; Hasty, P.; Ramsden, D. Ku is a 5′-dRP/AP lyase that excises nucleotide damage near broken ends. Nature 2010, 464, 1214–1217. [Google Scholar] [CrossRef] [Green Version]
- Tuteja, N.; Tuteja, R.; Ochem, A.; Taneja, P.; Huang, N.; Simoncsits, A.; Susic, S.; Rahman, K.; Marusic, L.; Chen, J.; et al. Human DNA helicase II: A novel DNA unwinding enzyme identified as the Ku autoantigen. EMBO J. 1994, 13, 4991–5001. [Google Scholar] [CrossRef]
- West, R.; Yaneva, M.; Lieber, M. Productive and nonproductive complexes of Ku and DNA-dependent protein kinase at DNA termini. Mol. Cell Biol. 1998, 18, 5908–5920. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.; Corpina, R.; Goldberg, J. Structure of the Ku heterodimer bound to DNA and its implications for double-strand break repair. Nature 2001, 412, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, S.; Schild-Poulter, C. Mapping the Ku Interactome Using Proximity-Dependent Biotin Identification in Human Cells. J. Proteome Res. 2019, 18, 1064–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couto, C.; Wang, H.; Green, J.; Kiely, R.; Siddaway, R.; Borer, C.; Pears, C.; Lakin, N. PARP regulates nonhomologous end joining through retention of Ku at double-strand breaks. J. Cell Biol. 2011, 194, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Galande, S.; Kohwi-Shigematsu, T. Poly(ADP-ribose) polymerase and Ku autoantigen form a complex and synergistically bind to matrix attachment sequences. J. Biol. Chem. 1999, 274, 20521–20528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariumi, Y.; Masutani, M.; Copeland, T.; Mimori, T.; Sugimura, T.; Shimotohno, K.; Ueda, K.; Hatanaka, M.; Noda, M. Suppression of the poly(ADP-ribose) polymerase activity by DNA-dependent protein kinase in vitro. Oncogene 1999, 18, 4616–4625. [Google Scholar] [CrossRef] [Green Version]
- BLi; Navarro, S.; Kasahara, N.; Comai, L. Identification and biochemical characterization of a Werner’s syndrome protein complex with Ku70/80 and poly(ADP-ribose) polymerase-1. J. Biol. Chem. 2004, 279, 13659–13667. [Google Scholar] [CrossRef] [Green Version]
- Tramontano, F.; Di Meglio, S.; Quesada, P. Co-localization of poly(ADPR)polymerase 1 (PARP-1) poly(ADPR)polymerase 2 (PARP-2) and related proteins in rat testis nuclear matrix defined by chemical cross-linking. J. Cell Biochem. 2005, 94, 58–66. [Google Scholar] [CrossRef]
- Pellarin, I.; Arnoldo, L.; Costantini, S.; Pegoraro, S.; Ros, G.; Penzo, C.; Triolo, G.; Demarchi, F.; Sgarra, R.; Vindigni, A.; et al. The Architectural Chromatin Factor High Mobility Group A1 Enhances DNA Ligase IV Activity Influencing DNA Repair. PLoS ONE 2016, 11, e0164258. [Google Scholar]
- Parvathaneni, S.; Stortchevoi, A.; Sommers, J.; Brosh, R., Jr.; Sharma, S. Human RECQ1 interacts with Ku70/80 and modulates DNA end-joining of double-strand breaks. PLoS ONE 2013, 8, e62481. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, R.; Russo, N.; Liu, M.; Basrur, V.; Bellile, E.; Palanisamy, N.; Scanlon, C.; van Tubergen, E.; Inglehart, R.; Metwally, T.; et al. TRIP13 promotes error-prone nonhomologous end joining and induces chemoresistance in head and neck cancer. Nat. Commun. 2014, 5, 4527. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Peter, S.; Droge, P.; Yan, J. Oncofetal HMGA2 effectively curbs unconstrained (+) and (−) DNA supercoiling. Sci. Rep. 2017, 7, 8440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.; Ramani, P.; Wong, S.; Zhao, X.; Ivanyi-Nagy, R.; Leong, T.; Chua, C.; Li, Z.; Hentze, H.; Tan, I.; et al. The chromatin structuring protein HMGA2 influences human subtelomere stability and cancer chemosensitivity. PLoS ONE 2019, 14, e0215696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, S.; Yu, H.; Ivanyi-Nagy, R.; Droge, P. Cell-based high-throughput compound screening reveals functional interaction between oncofetal HMGA2 and topoisomerase I. Nucleic Acids Res. 2016, 44, e162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luger, K.; Mader, A.; Richmond, R.; Sargent, D.; Richmond, T. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef]
- Funayama, R.; Saito, M.; Tanobe, H.; Ishikawa, F. Loss of linker histone H1 in cellular senescence. J. Cell Biol. 2006, 175, 869–880. [Google Scholar] [CrossRef] [Green Version]
- Go, C.; Knight, J.; Rajasekharan, A.; Rathod, B.; Hesketh, G.; Abe, K.; Youn, J.; Samavarchi-Tehrani, P.; Zhang, H.; Zhu, L.; et al. A proximity-dependent biotinylation map of a human cell. Nature 2021, 595, 120–124. [Google Scholar] [CrossRef]
- Divisato, G.; Chiariello, A.; Esposito, A.; Zoppoli, P.; Zambelli, F.; Elia, M.; Pesole, G.; Incarnato, D.; Passaro, F.; Piscitelli, S.; et al. Hmga2 protein loss alters nuclear envelope and 3D chromatin structure. BMC Biol. 2022, 20, 171. [Google Scholar] [CrossRef]
- Dangi-Garimella, S.; Sahai, V.; Ebine, K.; Kumar, K.; Munshi, H. Three-dimensional collagen I promotes gemcitabine resistance in vitro in pancreatic cancer cells through HMGA2-dependent histone acetyltransferase expression. PLoS ONE 2013, 8, e64566. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Niu, J.; Wang, X.; Zhang, Z.; Yang, R.; Yao, X.; Liu, F.; Li, W.; Pei, S.; Sun, H.; et al. P300-dependent acetylation of histone H3 is required for epidermal growth factor receptor-mediated high-mobility group protein A2 transcription in hepatocellular carcinoma. Cancer Sci. 2021, 112, 679–690. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, H.; Yu, L. HMGA2 promotes glioma invasion and poor prognosis via a long-range chromatin interaction. Cancer Med. 2018, 7, 3226–3239. [Google Scholar] [CrossRef]
- Yoo, Y.; Park, S.; Jo, E.; Choi, M.; Lee, K.; Hong, D.; Lee, S.; Lee, C.; Lee, Y.; Um, J.; et al. Overexpression of Replication-Dependent Histone Signifies a Subset of Dedifferentiated Liposarcoma with Increased Aggressiveness. Cancers 2021, 13, 3122. [Google Scholar] [CrossRef] [PubMed]
- Disney, J.; Johnson, K.; Magnuson, N.; Sylvester, S.; Reeves, R. High-mobility group protein HMG-I localizes to G/Q- and C-bands of human and mouse chromosomes. J. Cell Biol. 1989, 109, 1975–1982. [Google Scholar] [CrossRef]
- De Rop, V.; Padeganeh, A.; Maddox, P. CENP-A: The key player behind centromere identity, propagation, and kinetochore assembly. Chromosoma 2012, 121, 527–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weeks, S.; Metge, B.; Samant, R. The nucleolus: A central response hub for the stressors that drive cancer progression. Cell Mol. Life Sci. 2019, 76, 4511–4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Paul, L.; Grafi, G. The maize HMGA protein is localized to the nucleolus and can be acetylated in vitro at its globular domain, and phosphorylation by CDK reduces its binding activity to AT-rich DNA. Biochim. Biophys. Acta 2009, 1789, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Barboro, P.; D’Arrigo, C.; Mormino, M.; Coradeghini, R.; Parodi, S.; Patrone, E.; Balbi, C. An intranuclear frame for chromatin compartmentalization and higher-order folding. J. Cell Biochem. 2003, 88, 113–120. [Google Scholar] [CrossRef]
- Gribbon, C.; Dahm, R.; Prescott, A.; Quinlan, R. Association of the nuclear matrix component NuMA with the Cajal body and nuclear speckle compartments during transitions in transcriptional activity in lens cell differentiation. Eur. J. Cell Biol. 2002, 81, 557–566. [Google Scholar] [CrossRef]
- Ohata, H.; Miyazaki, M.; Otomo, R.; Matsushima-Hibiya, Y.; Otsubo, C.; Nagase, T.; Arakawa, H.; Yokota, J.; Nakagama, H.; Taya, Y.; et al. NuMA is required for the selective induction of p53 target genes. Mol. Cell Biol. 2013, 33, 2447–2457. [Google Scholar] [CrossRef] [Green Version]
- Endo, A.; Moyori, A.; Kobayashi, A.; Wong, R. Nuclear mitotic apparatus protein, NuMA, modulates p53-mediated transcription in cancer cells. Cell Death Dis. 2013, 4, e713. [Google Scholar] [CrossRef] [Green Version]
- Vidi, P.; Liu, J.; Salles, D.; Jayaraman, S.; Dorfman, G.; Gray, M.; Abad, P.; Moghe, P.; Irudayaraj, J.; Wiesmuller, L.; et al. NuMA promotes homologous recombination repair by regulating the accumulation of the ISWI ATPase SNF2h at DNA breaks. Nucleic Acids Res. 2014, 42, 6365–6379. [Google Scholar] [CrossRef] [Green Version]
- Vidi, P.; Chandramouly, G.; Gray, M.; Wang, L.; Liu, E.; Kim, J.; Roukos, V.; Bissell, M.; Moghe, P.; Lelievre, S. Interconnected contribution of tissue morphogenesis and the nuclear protein NuMA to the DNA damage response. J. Cell Sci. 2012, 125, 350–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Hsu, H.; Yeh, N. Apoptotic cleavage of NuMA at the C-terminal end is related to nuclear disruption and death amplification. J. Biomed. Sci. 2007, 14, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Lelievre, S.; Weaver, V.; Nickerson, J.; Larabell, C.; Bhaumik, A.; Petersen, O.; Bissell, M. Tissue phenotype depends on reciprocal interactions between the extracellular matrix and the structural organization of the nucleus. Proc. Natl. Acad. Sci. USA 1998, 95, 14711–14716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraman, S.; Chittiboyina, S.; Bai, Y.; Abad, P.; Vidi, P.; Stauffacher, C.; Lelievre, S. The nuclear mitotic apparatus protein NuMA controls rDNA transcription and mediates the nucleolar stress response in a p53-independent manner. Nucleic Acids Res. 2017, 45, 11725–11742. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BioID2 AP/MS | miniTurbo AP/MS | |||||||
|---|---|---|---|---|---|---|---|---|
| Experiment 1 | Experiment 2 | |||||||
| UniProt | Gene | # pep | log2 Ratio | # pep | log2 Ratio | # pep | log2 Ratio | Biogrid |
| Bait | ||||||||
| FSH2U8 | HMGA2 | 6 | Bait only | 4 | Bait only | 3 | Bait only | |
| Histones | ||||||||
| O75367 | H2AY | 13 | 8.52 | 9 | 2.95 | 4 | 1.28 | * |
| P16403 | HIST1H1C | 11 | 2.96 | 12 | 1.49 | 12 | 0.68 | |
| P10412 | HIST1H1E | 9 | Bait only | 11 | 1.46 | 11 | 0.42 | |
| Q93077 | HIST1H2AC | 6 | 5.5 | 7 | 3.16 | 5 | 1.55 | * |
| Q99878 | HIST1H2AJ | 8 | 6.22 | 8 | 3.22 | 5 | 1.57 | |
| P62805 | HIST1H4F | 13 | 6.34 | 12 | 3.15 | 10 | 1.58 | |
| Q71DI3 | HIST2H3D | 9 | 6.23 | 11 | 3.06 | 4 | 1.63 | |
| Q99880 | H2B1L/M | 11 | 6.22 | 9 | 3.1 | 4 | 1.56 | |
| Other Hits | ||||||||
| P25440 | BRD2 | 10 | Bait only | 5 | Bait only | 7 | Bait only | |
| Q13185 | CBX3 | 6 | Bait only | 6 | 1.07 | 7 | 1.38 | |
| E9PEI0 | CDCA2 | 4 | Bait only | 1 | 1.73 | 7 | 3.26 | |
| Q53HL2 | CDCA8 | 6 | Bait only | 2 | 2.53 | 10 | 5.55 | |
| Q96JM3 | CHAMP1 | 18 | Bait only | 14 | 1.34 | 13 | 1.41 | |
| Q5QJE6 | DNTTIP2 | 8 | Bait only | 3 | 2.05 | 7 | 1.41 | |
| P78347 | GTF2I | 26 | Bait only | 16 | 1.59 | 20 | 0.46 | |
| O14929 | HAT1 | 3 | Bait only | 4 | 2.48 | 1 | Bait only | |
| Q5T7C4 | HMGB1 | 6 | 3.05 | 4 | 0.82 | 5 | 1.85 | |
| D6R9A6 | HMGB2 | 5 | 4.13 | 4 | 0.82 | 4 | 1.66 | |
| Q9H0C8 | ILKAP | 11 | Bait only | 9 | 1.8 | 5 | 0.85 | |
| P42167 | LAP2B | 19 | Bait only | 22 | 1.08 | 9 | 2.19 | |
| Q14676 | MDC1 | 32 | Bait only | 39 | 2.61 | 5 | 0.62 | |
| P46013 | MKI67 | 4 | Bait only | 83 | 1.62 | 106 | 2.51 | |
| Q14980 | NUMA1 | 91 | 3.25 | 46 | 2.49 | 12 | 1.29 | |
| P09874 | PARP1 | 66 | 5.65 | 54 | 3.9 | 24 | 2.19 | * |
| Q7Z3K3 | POGZ | 15 | Bait only | 6 | 4.23 | 4 | 1.19 | |
| O75475 | PSIP1 | 16 | Bait only | 7 | 3.02 | 3 | Bait only | * |
| Q08945 | SSRP1 | 17 | Bait only | 8 | 1.48 | 2 | 1.26 | |
| P11388 | TOP2A | 55 | 4.13 | 33 | 1.9 | 15 | 1.86 | |
| Q9ULW0 | TPX2 | 20 | Bait only | 20 | 1.56 | 23 | 2.96 | |
| Q14669 | TRIPC | 25 | Bait only | 14 | 2.37 | 20 | 1.59 | |
| B7ZM82 | WIZ | 13 | Bait only | 9 | 1.95 | 11 | Bait only | |
| P13010 | XRCC5 | 32 | 4.13 | 20 | 2.8 | 10 | 0.86 | * |
| P12956 | XRCC6 | 29 | 4.29 | 20 | 2.8 | 10 | 0.75 | * |
| P17028 | ZNF24 | 6 | Bait only | 3 | Bait only | 2 | Bait only | |
| Q96KM6 | ZNF512B | 3 | Bait only | 5 | 2.57 | 2 | Bait only | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaudreau-Lapierre, A.; Klonisch, T.; Nicolas, H.; Thanasupawat, T.; Trinkle-Mulcahy, L.; Hombach-Klonisch, S. Nuclear High Mobility Group A2 (HMGA2) Interactome Revealed by Biotin Proximity Labeling. Int. J. Mol. Sci. 2023, 24, 4246. https://doi.org/10.3390/ijms24044246
Gaudreau-Lapierre A, Klonisch T, Nicolas H, Thanasupawat T, Trinkle-Mulcahy L, Hombach-Klonisch S. Nuclear High Mobility Group A2 (HMGA2) Interactome Revealed by Biotin Proximity Labeling. International Journal of Molecular Sciences. 2023; 24(4):4246. https://doi.org/10.3390/ijms24044246
Chicago/Turabian StyleGaudreau-Lapierre, Antoine, Thomas Klonisch, Hannah Nicolas, Thatchawan Thanasupawat, Laura Trinkle-Mulcahy, and Sabine Hombach-Klonisch. 2023. "Nuclear High Mobility Group A2 (HMGA2) Interactome Revealed by Biotin Proximity Labeling" International Journal of Molecular Sciences 24, no. 4: 4246. https://doi.org/10.3390/ijms24044246
APA StyleGaudreau-Lapierre, A., Klonisch, T., Nicolas, H., Thanasupawat, T., Trinkle-Mulcahy, L., & Hombach-Klonisch, S. (2023). Nuclear High Mobility Group A2 (HMGA2) Interactome Revealed by Biotin Proximity Labeling. International Journal of Molecular Sciences, 24(4), 4246. https://doi.org/10.3390/ijms24044246