

The Main Theories on the Pathogenesis of Endometriosis


Abstract
:1. Introduction
1.1. Epidemiology
1.2. Symptomatology
1.3. Subtypes of Endometriosis
2. Theories on the Pathogenesis of Endometriosis
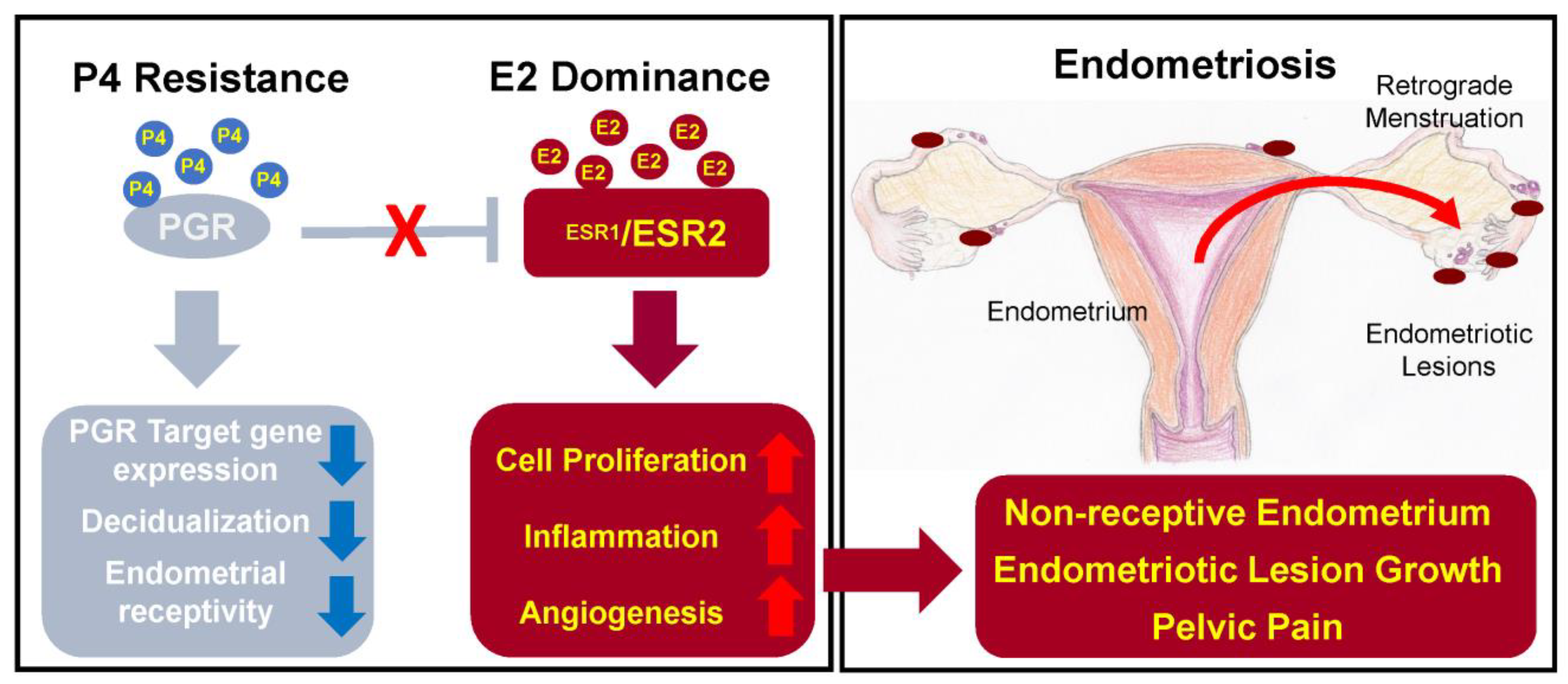
2.1. Retrograde Menstruation
2.2. Benign Metastasis
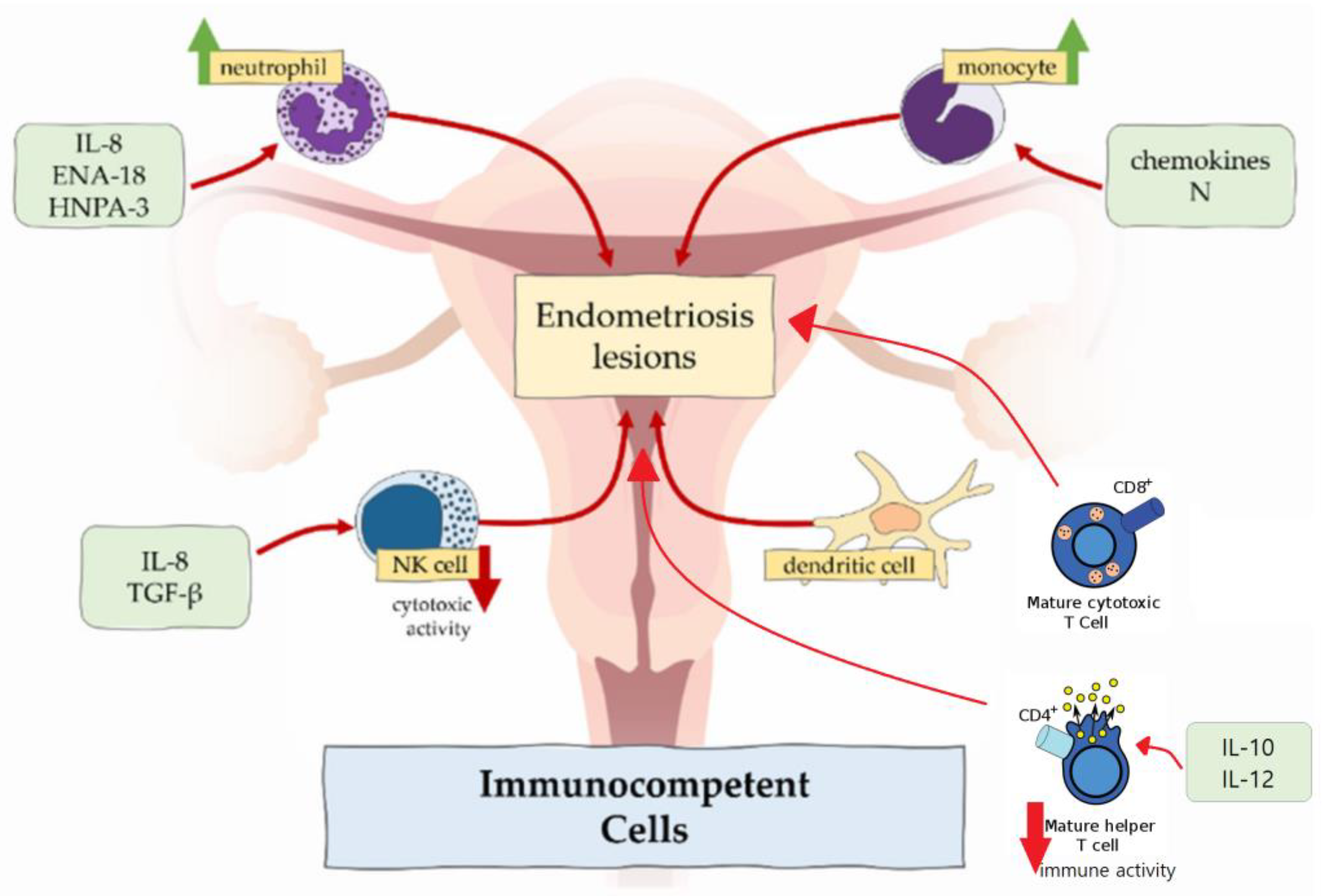
2.3. Immune Dysregulation
2.3.1. Macrophages
2.3.2. Neutrophils
2.3.3. NK Cells
2.3.4. Dendritic Cells
2.3.5. T Cells
2.4. Coelomic Metaplasia
2.5. Embryonic Rest Theory
2.6. Endometrial Stem Cell Recruitment Theory
2.7. Bone Marrow-Derived Stem Cell Theory
2.8. Hormonal Imbalance
2.8.1. Oestrogen
2.8.2. Progesterone
2.9. Alterations in Epigenetic Regulation
2.9.1. DNA Methyltransferases
2.9.2. Methylation of the HOXA10
2.9.3. Histone Acetylation
2.10. Micro-RNAs
2.11. Carcinogenetic Pathways in Endometriosis
2.12. External Environmental Factors
2.12.1. Lifestyle
2.12.2. Dioxins and Polychlorinated Biphenyls (PCBs)
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CXCL10 | C-X-C chemokine motif ligand 10 |
| ENA-78 | Epithelial neutrophil-activating peptide 8 |
| ER | Estrogen receptor |
| FGF | Fibroblast growth factor |
| hMLH1 | Human mutL homolog 1 |
| HNP1-3 | Human neutrophil peptides 1-3 |
| HOXA10 | Human Homeobox A10 |
| IL | Interleukin |
| KRAS | Kirsten rat sarcoma virus |
| LIF | Leukaemia inhibitory factor |
| LINE-1 | Long interspersed element-1 |
| NGF | Nerve growth factors |
| NK cells | Natural killer cells |
| NOD-SCID | Nonobese diabetic/severe combined immunodeficiency |
| PCBs | Polychlorinated biphenyls |
| PGR | Progesterone receptor |
| PTEN | Phosphatase and tensin homolog |
| RASSF2 | Ras-association domain family member 2 |
| RUNX3 | Runt-related transcription factor 3 |
| TCDD | 2,3,7,8-tetrachlorodibenzo-p-dioxin |
| TGF-β | Transforming growth factor β |
| Th1 and Th2 | Type 1 T lymphocytes and type 2 T lymphocytes |
| TNFα | Tumour necrosis factor α |
| Treg | Regulatory T cells |
| VEGF | Vascular endothelial growth factor |
References
- World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/endometriosis (accessed on 27 September 2022).
- Saunders, P.T.K.; Horne, A.W. Endometriosis: Etiology, pathobiology, and therapeutic prospects. Cell 2021, 184, 2807–2824. [Google Scholar] [CrossRef]
- Smolarz, B.; Szyłło, K.; Romanowicz, H. Endometriosis: Epidemiology, classification, pathogenesis, treatment and genetics (Review of literature). Int. J. Mol. Sci. 2021, 22, 10554. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Singh, V.; Tayade, C. Biomarkers in endometriosis: Challenges and opportunities. Fertil. Steril. 2017, 107, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horne, A.W.; Saunders, P.T.K. SnapShot: Endometriosis. Cell 2019, 179, 1677. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nicholes, K.; Shih, I.M. The origin and pathogenesis of endometriosis. Annu. Rev. Pathol. 2020, 15, 71–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yovich, J.L.; Rowlands, P.K.; Lingham, S.; Sillender, M.; Srinivasan, S. Pathogenesis of endometriosis: Look no further than John Sampson. Reprod. Biomed. Online 2020, 40, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampson, J.A. The development of the implantation theory for the origin of peritoneal endometriosis. Am. J. Obstet. Gynecol. 1940, 40, 549–557. [Google Scholar] [CrossRef]
- Nisolle, M.; Donnez, J. Peritoneal endometriosis, ovarian endometriosis, and adenomyotic nodules of the rectovaginal septum are three different entities. Fertil. Steril. 1997, 68, 585–596. [Google Scholar] [CrossRef]
- Izumi, G.; Koga, K.; Takamura, M.; Makabe, T.; Satake, E.; Takeuchi, A.; Taguchi, A.; Urata, Y.; Fujii, T.; Osuga, Y. Involvement of immune cells in the pathogenesis of endometriosis. J. Obstet. Gynaecol. Res. 2018, 44, 191–198. [Google Scholar] [CrossRef] [Green Version]
- D’Hooghe, T.M.; Debrock, S. Endometriosis, retrograde menstruation and peritoneal inflammation in women and in baboons. Hum. Reprod. Update 2002, 8, 84–88. [Google Scholar] [CrossRef]
- Signorile, P.G.; Viceconte, R.; Baldi, A. New insights in pathogenesis of endometriosis. Front. Med. 2022, 9, 879015. [Google Scholar] [CrossRef] [PubMed]
- Sampson, J.A. Metastatic or embolic endometriosis, due to the menstrual dissemination of endometrial tissue into the venous circulation. Am. J. Pathol. 1927, 3, 93–110. [Google Scholar] [PubMed]
- Jerman, L.F.; Hey-Cunningham, A.J. The role of the lymphatic system in endometriosis: A comprehensive review of the literature. Biol. Reprod. 2015, 64, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Du, K.; Xiao, L.; He, X.; Dai, F.; Tang, J. Endometriosis in para-aortic lymph node resembling a malignancy: A case report and literature review. BMC Womens Health 2022, 22, 101. [Google Scholar] [CrossRef]
- Parag, D.J.; Vijayanand, K.M.; Lakshmi, K. Gastrointestinal deep infiltrative endometriosis with lymph node involvement. Indian J. Pathol. Microbiol. 2021, 64, 213–215. [Google Scholar]
- Law, Y.Y.; Patel, R.; Yorke, R.; Bailey, H.R.; Van Eps, J.L. A case of infiltrative cecal endometriosis with appendiceal obliteration and lymph node involvement. J. Surg. Case. Rep. 2020, 2020, rjaa396. [Google Scholar] [CrossRef]
- Takehara, M.; Ueda, M.; Yamashita, Y.; Terai, Y.; Hung, Y.C.; Ueki, M. Vascular endothelial growth factor A and C gene expression in endometriosis. Hum. Pathol. 2004, 35, 1369–1375. [Google Scholar] [CrossRef]
- Keichel, S.; Barcena de Arellano, M.L.; Reichelt, U.; Riedlinger, W.F.; Schneider, A.; Köhler, C.; Mechsner, S. Lymphangiogenesis in deep infiltrating endometriosis. Hum. Reprod. 2011, 26, 2713–2720. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, R.; Stratopoulou, C.A.; Dolmans, M.M. Pathogenesis of endometriosis: New insights into prospective therapies. Int. J. Mol. Sci. 2021, 22, 11700. [Google Scholar] [CrossRef]
- Abramiuk, M.; Grywalska, E.; Małkowska, P.; Sierawska, O.; Hrynkiewicz, R.; Niedźwiedzka-Rystwej, P. The role of the immune system in the development of endometriosis. Cells 2022, 11, 2028. [Google Scholar] [CrossRef]
- Vallvé-Juanico, J.; Houshdaran, S.; Giudice, L.C. The endometrial immune environment of women with endometriosis. Hum. Reprod. Update 2019, 25, 565–592. [Google Scholar] [CrossRef]
- Berbic, M.; Schulke, L.; Markham, R.; Tokushige, N.; Russell, P.; Fraser, I.S. Macrophage expression in endometrium of women with and without endometriosis. Hum. Reprod. 2009, 24, 325–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.H.; Chuang, P.C.; Lin, Y.J.; Tsai, S.J. Suppression of annexin A2 by prostaglandin E₂ impairs phagocytic ability of peritoneal macrophages in women with endometriosis. Hum. Reprod. 2013, 28, 1045–1053. [Google Scholar] [CrossRef]
- Bacci, M.; Capobianco, A.; Monno, A.; Cottone, L.; Di Puppo, F.; Camisa, B.; Mariani, M.; Brignole, C.; Ponzoni, M.; Ferrari, S.; et al. Macrophages are alternatively activated in patients with endometriosis and required for growth and vascularization of lesions in a mouse model of disease. Am. J. Pathol. 2009, 175, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takamura, M.; Koga, K.; Izumi, G.; Urata, Y.; Nagai, M.; Hasegawa, A.; Harada, M.; Hirata, T.; Hirota, Y.; Wada-Hiraike, O.; et al. Neutrophil depletion reduces endometriotic lesion formation in mice. Am. J. Reprod. Immunol. 2016, 76, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Jiang, I.; Yong, P.J.; Allaire, C.; Bedaiwy, M.A. Intricate connections between the microbiota and endometriosis. Int. J. Mol. Sci. 2021, 22, 5644. [Google Scholar] [CrossRef]
- Maridas, D.E.; Hey-Cunningham, A.J.; Ng, C.H.M.; Markham, R.; Fraser, I.S.; Berbic, M. Peripheral and endometrial dendritic cell populations during the normal cycle and in the presence of endometriosis. J. Endometr. Pelvic Pain Disord. 2014, 6, 67–119. [Google Scholar] [CrossRef] [Green Version]
- Rizner, T.L. Estrogen metabolism and action in endometriosis. Mol. Cell. Endocrinol. 2009, 307, 8–18. [Google Scholar] [CrossRef]
- Slabe, N.; Meden-Vrtovec, H.; Verdenik, I.; Kosir-Pogacnik, R.; Ihan, A. Cytotoxic T-cells in peripheral blood in women with endometriosis. Geburtshilfe. Frauenheilkd. 2013, 73, 1042–1048. [Google Scholar] [CrossRef] [Green Version]
- Konrad, L.; Dietze, R.; Kudipudi, P.K.; Horné, F.; Meinhold-Heerlein, I. Endometriosis in MRKH cases as a proof for the coelomic metaplasia hypothesis? Reproduction 2019, 158, R41–R47. [Google Scholar] [CrossRef]
- Cho, M.K.; Kim, C.H.; Oh, S.T. Endometriosis in a patient with Rokitansky-Kuster-Hauser syndrome. J. Obstet. Gynaecol. Res. 2009, 35, 994–996. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Yoshimura, Y. Stem cell theory for the pathogenesis of endometriosis. Front. Biosci. 2012, 4, 2754–2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordts, S.; Koninckx, P.; Brosens, I. Pathogenesis of deep endometriosis. Fertil. Steril. 2017, 108, 872–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueira, P.G.; Abrão, M.S.; Krikun, G.; Taylor, H.S. Stem cells in endometrium and their role in the pathogenesis of endometriosis. Ann. N. Y. Acad Sci. 2011, 1221, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Djokovic, D.; Calhaz-Jorge, C. Somatic stem cells and their dysfunction in endometriosis. Front. Surg. 2014, 1, 51. [Google Scholar] [CrossRef] [PubMed]
- Cervelló, I.; Mas, A.; Gil-Sanchis, C.; Peris, L.; Faus, A.; Saunders, P.T.; Critchley, H.O.; Simón, C. Reconstruction of endometrium from human endometrial side population cell lines. PLoS ONE 2011, 6, e21221. [Google Scholar] [CrossRef]
- Marquardt, R.M.; Kim, T.H.; Shin, J.H.; Jeong, J.W. Progesterone and estrogen signaling in the endometrium: What goes wrong in endometriosis? Int. J. Mol. Sci. 2019, 20, 3822. [Google Scholar] [CrossRef] [Green Version]
- Koukoura, O.; Sifakis, S.; Spandidos, D.A. DNA methylation in endometriosis (Review). Mol. Med. Rep. 2016, 13, 2939–2948. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Yan, Y.; Liu, Z.; Wang, Y. Inflammation and endometriosis. Front. Biosci. 2016, 21, 941–948. [Google Scholar]
- Kim, J.J.; Kurita, T.; Bulun, S.E. Progesterone action in endometrial cancer, endometriosis, uterine fibroids, and breast cancer. Endocr. Rev. 2013, 34, 130–162. [Google Scholar] [CrossRef] [Green Version]
- Asghari, S.; Valizadeh, A.; Aghebati-Maleki, L.; Nouri, M.; Yousefi, M. Endometriosis: Perspective, lights, and shadows of etiology. Biomed. Pharmacother. 2018, 106, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Laganà, A.S.; Garzon, S.; Götte, M.; Viganò, P.; Franchi, M.; Ghezzi, F.; Martin, D.C. The pathogenesis of endometriosis: Molecular and cell biology insights. Int. J. Mol. Sci. 2019, 20, 5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esfandiari, F.; Favaedi, R.; Heidari-Khoei, H.; Chitsazian, F.; Yari, S.; Piryaei, A.; Ghafari, F.; Baharvand, H.; Shahhoseini, M. Insight into epigenetics of human endometriosis organoids: DNA methylation analysis of HOX genes and their cofactors. Fertil. Steril. 2021, 115, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Teague, E.M.; Print, C.G.; Hull, M.L. The role of microRNAs in endometriosis and associated reproductive conditions. Hum Reprod. Update 2010, 16, 142–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raja, M.H.R.; Farooqui, N.; Zuberi, N.; Ashraf, M.; Azhar, A.; Baig, R.; Badar, B.; Rehman, R. Endometriosis, infertility and microRNA’s: A review. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 102157. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Lin, X.; Shi, T.; Tian, Y. MiRNA-223 expression in patient-derived eutopic and ectopic endometrial stromal cells and its effect on epithelial-to-mesenchymal transition in endometriosis. Clinics 2022, 77, 100112. [Google Scholar] [CrossRef]
- Wu, Y.; Yuan, W.; Ding, H.; Wu, X. Serum exosomal miRNA from endometriosis patients correlates with disease severity. Gynecol. Obstet. 2022, 305, 117–127. [Google Scholar] [CrossRef]
- Králíčková, M.; Losan, P.; Vetvicka, V. Endometriosis and cancer. Womens Health 2014, 10, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Anglesio, M.S.; Papadopoulos, N.; Ayhan, A.; Nazeran, T.M.; Noë, M.; Horlings, H.M.; Lum, A.; Jones, S.; Senz, J.; Seckin, T.; et al. Cancer-associated mutations in endometriosis without cancer. N. Engl. J. Med. 2017, 376, 1835–1848. [Google Scholar] [CrossRef] [Green Version]
- Coiplet, E.; Courbiere, B.; Agostini, A.; Boubli, L.; Bretelle, F.; Netter, A. Endometriosis and environmental factors: A critical review. J. Gynecol. Obstet. Hum. Reprod. 2022, 51, 102418. [Google Scholar] [CrossRef]
- Kechagias, K.S.; Katsikas Triantafyllidis, K.; Kyriakidou, M.; Giannos, P.; Kalliala, I.; Veroniki, A.A.; Paraskevaidi, M.; Kyrgiou, M. The relation between caffeine consumption and endometriosis: An updated systematic review and meta-analysis. Nutrients 2021, 13, 3457. [Google Scholar] [CrossRef] [PubMed]
- Polak, G.; Banaszewska, B.; Filip, M.; Radwan, M.; Wdowiak, A. Environmental factors and endometriosis. Int. J. Environ. Res. Public Health 2021, 18, 11025. [Google Scholar] [CrossRef] [PubMed]
- Ballester, M.; Dehan, P.; Béliard, A.; Brichant, G.; Nisolle, M. Role of genetic and environmental factors in the development of endometriosis. Rev. Med. Liege 2012, 67, 374–380. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Micro-RNAs | Changes | Effect |
|---|---|---|
| miRNA-135a/b | Upregulated | Dysregulation of HOXA10 expression, progesterone resistance |
| miR-199 | Downregulated | Synthesis of pro-inflammatory prostaglandins due to lack of COX-2 suppression |
| miRNA-96b | Downregulated | Increased proliferation of the endometrial lesions |
| miR-126 | Upregulated | Neoangiogenesis due to increased VEGF and FGF |
| miRNA-223 | Downregulated | Proliferation, invasion, migration of endometrial stromal cells, epithelial-to-mesenchymal transition |
| miRNA-21 | Upregulated | Growth, proliferation and angiogenesis of ectopic stromal cells |
| miR-26b-5p | Downregulated | Activation of MAPK and PI3K-Akt pathways: inflammation, cell growth, differentiation and proliferation, angiogenesis |
| miR-215-5p | Downregulated | |
| miR-6795-3p | Upregulated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamceva, J.; Uljanovs, R.; Strumfa, I. The Main Theories on the Pathogenesis of Endometriosis. Int. J. Mol. Sci. 2023, 24, 4254. https://doi.org/10.3390/ijms24054254
Lamceva J, Uljanovs R, Strumfa I. The Main Theories on the Pathogenesis of Endometriosis. International Journal of Molecular Sciences. 2023; 24(5):4254. https://doi.org/10.3390/ijms24054254
Chicago/Turabian StyleLamceva, Jelizaveta, Romans Uljanovs, and Ilze Strumfa. 2023. "The Main Theories on the Pathogenesis of Endometriosis" International Journal of Molecular Sciences 24, no. 5: 4254. https://doi.org/10.3390/ijms24054254
APA StyleLamceva, J., Uljanovs, R., & Strumfa, I. (2023). The Main Theories on the Pathogenesis of Endometriosis. International Journal of Molecular Sciences, 24(5), 4254. https://doi.org/10.3390/ijms24054254