
Polarized Anti-Inflammatory Mesenchymal Stem Cells Increase Hippocampal Neurogenesis and Improve Cognitive Function in Aged Mice

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Culturing MSC in the Presence of PACAP Polarized MSC towards MSC2 with an Anti-Inflammatory Phenotype
2.2. Systemic Administration of Polarized MSC Reduced the Levels of Pro-Aging Chemokines in the Serum of Aged Mice
2.3. Systemic Administration of Polarized MSC Increased Hippocampal Neurogenesis in Aged Mice
2.4. Systemic Administration of Polarized MSC Improves Cognitive Function in Aged Mice
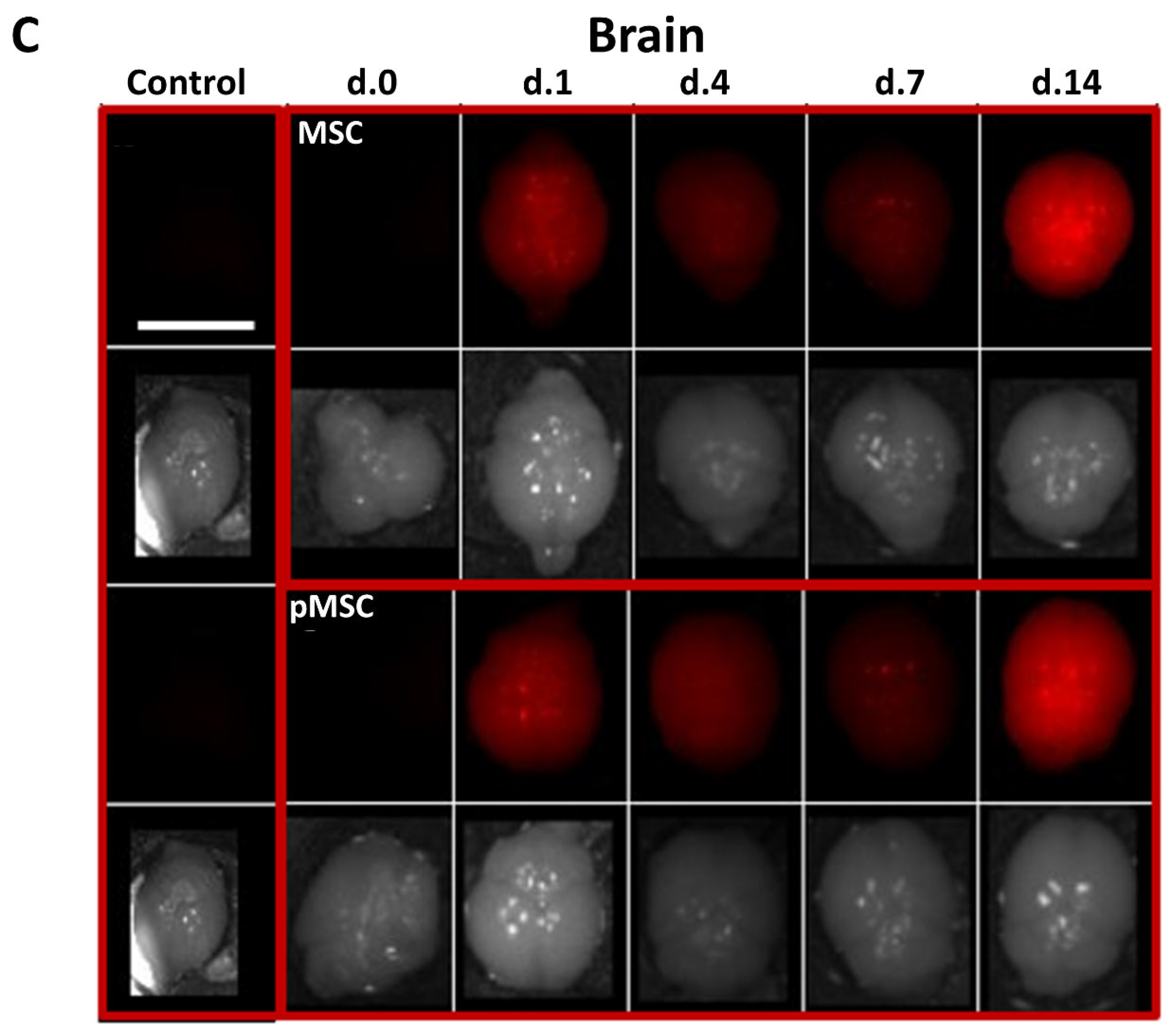
2.5. Naïve and Polarized MSC Engraft to the Brain, Lungs and Liver following Intravenous Injection to Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mesenchymal Stem Cells Isolation and Culture
4.3. Flow Cytometry Immunophenotyping
4.4. Cytokine Array
4.5. Behavioral Assays
4.5.1. Open Field Test
4.5.2. Morris Water Maze test
4.5.3. Y-Maze
4.6. Immunohistochemistry
4.7. Real-Time PCR
4.8. Cell Engraftment Analysis
4.9. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Denoth-Lippuner, A.; Jessberger, S. Formation and integration of new neurons in the adult hippocampus. Nat. Rev. Neurosci. 2021, 22, 223–236. [Google Scholar] [CrossRef]
- Toda, T.; Gage, F.H. Review: Adult neurogenesis contributes to hippocampal plasticity. Cell Tissue Res. 2018, 373, 693–709. [Google Scholar] [CrossRef] [PubMed]
- Chesnokova, V.; Pechnick, R.N.; Wawrowsky, K. Chronic peripheral inflammation, hippocampal neurogenesis, and behavior. Brain Behav. Immun. 2016, 58, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.K.; White, C.W., 3rd; Villeda, S.A. The systemic environment: At the interface of aging and adult neurogenesis. Cell Tissue Res. 2018, 371, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Ge, J.; Huang, C.; Liu, H.; Jiang, H. Application of mesenchymal stem cell therapy for aging frailty: From mechanisms to therapeutics. Theranostics 2021, 11, 5675–5685. [Google Scholar] [CrossRef] [PubMed]
- Satija, N.K.; Gurudutta, G.U.; Sharma, S.; Afrin, F.; Gupta, P.; Verma, Y.K.; Singh, V.K.; Tripathi, R.P. Mesenchymal stem cells: Molecular targets for tissue engineering. Stem Cells Dev. 2007, 16, 7–23. [Google Scholar] [CrossRef]
- Prockop, D.J. Marrow Stromal Cells as Stem Cells for Nonhematopoietic Tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Merimi, M.; El-Majzoub, R.; Lagneaux, L.; Moussa Agha, D.; Bouhtit, F.; Meuleman, N.; Fahmi, H.; Lewalle, P.; Fayyad-Kazan, M.; Najar, M. The Therapeutic Potential of Mesenchymal Stromal Cells for Regenerative Medicine: Current Knowledge and Future Understandings. Front. Cell Dev. Biol. 2021, 9, 661532. [Google Scholar] [CrossRef]
- Luz-Crawford, P.; Kurte, M.; Bravo-Alegria, J.; Contreras, R.; Nova-Lamperti, E.; Tejedor, G.; Noel, D.; Jorgensen, C.; Figueroa, F.; Djouad, F.; et al. Mesenchymal stem cells generate a CD4+CD25+Foxp3+ regulatory T cell population during the differentiation process of Th1 and Th17 cells. Stem Cell Res. Ther. 2013, 4, 65. [Google Scholar] [CrossRef] [Green Version]
- Müller, L.; Tunger, A.; Wobus, M.; von Bonin, M.; Towers, R.; Bornhäuser, M.; Dazzi, F.; Wehner, R.; Schmitz, M. Immunomodulatory Properties of Mesenchymal Stromal Cells: An Update. Front. Cell Dev. Biol. 2021, 9, 637725. [Google Scholar] [CrossRef]
- Benvenuto, F.; Ferrari, S.; Gerdoni, E.; Gualandi, F.; Frassoni, F.; Pistoia, V.; Mancardi, G.; Uccelli, A. Human mesenchymal stem cells promote survival of T cells in a quiescent state. Stem Cells 2007, 25, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- English, K.; Wood, K.J. Mesenchymal stromal cells in transplantation rejection and tolerance. Cold Spring Harb. Perspect. Med. 2013, 3, a015560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoshina, O.O.; Kozhin, P.M.; Shadrin, V.S.; Romashin, D.D.; Rusanov, A.L.; Luzgina, N.G. Phenotypic Features of Mesenchymal Stem Cell Subpopulations Obtained under the Influence of Various Toll-Like Receptors Ligands. Bull. Exp. Biol. Med. 2021, 170, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Waterman, R.S.; Tomchuck, S.L.; Henkle, S.L.; Betancourt, A.M. A new mesenchymal stem cell (MSC) paradigm: Polarization into a pro-inflammatory MSC1 or an Immunosuppressive MSC2 phenotype. PLoS ONE 2010, 5, e10088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobshtis, N.; Tfilin, M.; Wolfson, M.; Fraifeld, V.E.; Turgeman, G. Transplantation of mesenchymal stem cells reverses behavioural deficits and impaired neurogenesis caused by prenatal exposure to valproic acid. Oncotarget 2017, 8, 17443–17452. [Google Scholar] [CrossRef]
- Abad, C.; Tan, Y.-V. Immunomodulatory Roles of PACAP and VIP: Lessons from Knockout Mice. J. Mol. Neurosci. 2018, 66, 102–113. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Ohtaki, H.; Song, D.; Sato, A.; Watanabe, J.; Hiraizumi, Y.; Nakamachi, T.; Xu, Z.; Dohi, K.; Hashimoto, H.; et al. Human mesenchymal stem/stromal cells suppress spinal inflammation in mice with contribution of pituitary adenylate cyclase-activating polypeptide (PACAP). J. Neuroinflamm. 2015, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, J.; Pérez, G.; Semple, S.L.; Rodríguez-Ramos, T.; Díaz-Rosales, P.; del Camino Ordás, M.; Lugo, J.M.; Dixon, B.; Tafalla, C.; Estrada, M.P.; et al. First in vivo evidence of pituitary adenylate cyclase-activating polypeptide antiviral activity in teleost. Fish Shellfish Immunol. 2020, 103, 58–65. [Google Scholar] [CrossRef]
- Voice, J.K.; Dorsam, G.; Lee, H.; Kong, Y.; Goetzl, E.J. Allergic diathesis in transgenic mice with constitutive T cell expression of inducible vasoactive intestinal peptide receptor. FASEB J. 2001, 15, 2489–2496. [Google Scholar] [CrossRef] [Green Version]
- Villeda, S.A.; Luo, J.; Mosher, K.I.; Zou, B.; Britschgi, M.; Bieri, G.; Stan, T.M.; Fainberg, N.; Ding, Z.; Eggel, A.; et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature 2011, 477, 90–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Hur, T.; Ben-Menachem, O.; Furer, V.; Einstein, O.; Mizrachi-Kol, R.; Grigoriadis, N. Effects of proinflammatory cytokines on the growth, fate, and motility of multipotential neural precursor cells. Mol. Cell. Neurosci. 2003, 24, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Monje, M.L.; Palmer, T. Radiation injury and neurogenesis. Curr. Opin. Neurol. 2003, 16, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Boehme, M.; Guenther, M.; Stahr, A.; Liebmann, M.; Jaenisch, N.; Witte, O.W.; Frahm, C. Impact of indomethacin on neuroinflammation and hippocampal neurogenesis in aged mice. Neurosci. Lett. 2014, 572, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Gobshtis, N.; Tfilin, M.; Fraifeld, V.E.; Turgeman, G. Transplantation of mesenchymal stem cells causes long-term alleviation of schizophrenia-like behaviour coupled with increased neurogenesis. Mol. Psychiatry 2021, 26, 4448–4463. [Google Scholar] [CrossRef]
- Zappa Villar, M.F.; Lehmann, M.; García, M.G.; Mazzolini, G.; Morel, G.R.; Cónsole, G.M.; Podhajcer, O.; Reggiani, P.C.; Goya, R.G. Mesenchymal stem cell therapy improves spatial memory and hippocampal structure in aging rats. Behav. Brain Res. 2019, 374, 111887. [Google Scholar] [CrossRef]
- Park, D.; Yang, G.; Bae, D.K.; Lee, S.H.; Yang, Y.-H.; Kyung, J.; Kim, D.; Choi, E.-K.; Choi, K.-C.; Kim, S.U.; et al. Human adipose tissue-derived mesenchymal stem cells improve cognitive function and physical activity in ageing mice. J. Neurosci. Res. 2013, 91, 660–670. [Google Scholar] [CrossRef]
- Sandu, R.E.; Balseanu, A.T.; Bogdan, C.; Slevin, M.; Petcu, E.; Popa-Wagner, A. Stem cell therapies in preclinical models of stroke. Is the aged brain microenvironment refractory to cell therapy? Exp. Gerontol. 2017, 94, 73–77. [Google Scholar] [CrossRef]
- Cao, N.; Liao, T.; Liu, J.; Fan, Z.; Zeng, Q.; Zhou, J.; Pei, H.; Xi, J.; He, L.; Chen, L.; et al. Clinical-grade human umbilical cord-derived mesenchymal stem cells reverse cognitive aging via improving synaptic plasticity and endogenous neurogenesis. Cell Death Dis. 2017, 8, e2996. [Google Scholar] [CrossRef] [Green Version]
- Millard, S.M.; Fisk, N.M. Mesenchymal stem cells for systemic therapy: Shotgun approach or magic bullets? BioEssays 2013, 35, 173–182. [Google Scholar] [CrossRef]
- El-Akabawy, G.; Aabed, K.; Rashed, L.; Amin, S.; AlSaati, I.; Al-Fayez, M. Preventive effects of bone marrow-derived mesenchymal stem cell transplantation in a D-galactose-induced brain ageing in rats. Folia Morphol. 2021, 81, 632–649. [Google Scholar] [CrossRef] [PubMed]
- Fabian, C.; Naaldijk, Y.; Leovsky, C.; Johnson, A.A.; Rudolph, L.; Jaeger, C.; Arnold, K.; Stolzing, A. Distribution pattern following systemic mesenchymal stem cell injection depends on the age of the recipient and neuronal health. Stem Cell Res. Ther. 2017, 8, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.K.; He, Y.; Park, J.S.; Bieri, G.; Snethlage, C.E.; Lin, K.; Gontier, G.; Wabl, R.; Plambeck, K.E.; Udeochu, J.; et al. beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis. Nat. Med. 2015, 21, 932–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.W.; Haditsch, U.; Cord, B.J.; Guzman, R.; Kim, S.J.; Boettcher, C.; Priller, J.; Ormerod, B.K.; Palmer, T.D. Absence of CCL2 is sufficient to restore hippocampal neurogenesis following cranial irradiation. Brain Behav. Immun. 2013, 30, 33–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tfilin, M.; Gobshtis, N.; Fozailoff, D.; Fraifeld, V.E.; Turgeman, G. Polarized Anti-Inflammatory Mesenchymal Stem Cells Increase Hippocampal Neurogenesis and Improve Cognitive Function in Aged Mice. Int. J. Mol. Sci. 2023, 24, 4490. https://doi.org/10.3390/ijms24054490
Tfilin M, Gobshtis N, Fozailoff D, Fraifeld VE, Turgeman G. Polarized Anti-Inflammatory Mesenchymal Stem Cells Increase Hippocampal Neurogenesis and Improve Cognitive Function in Aged Mice. International Journal of Molecular Sciences. 2023; 24(5):4490. https://doi.org/10.3390/ijms24054490
Chicago/Turabian StyleTfilin, Matanel, Nikolai Gobshtis, David Fozailoff, Vadim E. Fraifeld, and Gadi Turgeman. 2023. "Polarized Anti-Inflammatory Mesenchymal Stem Cells Increase Hippocampal Neurogenesis and Improve Cognitive Function in Aged Mice" International Journal of Molecular Sciences 24, no. 5: 4490. https://doi.org/10.3390/ijms24054490
APA StyleTfilin, M., Gobshtis, N., Fozailoff, D., Fraifeld, V. E., & Turgeman, G. (2023). Polarized Anti-Inflammatory Mesenchymal Stem Cells Increase Hippocampal Neurogenesis and Improve Cognitive Function in Aged Mice. International Journal of Molecular Sciences, 24(5), 4490. https://doi.org/10.3390/ijms24054490