

Universal Stress Proteins: From Gene to Function

Abstract
:1. Introduction
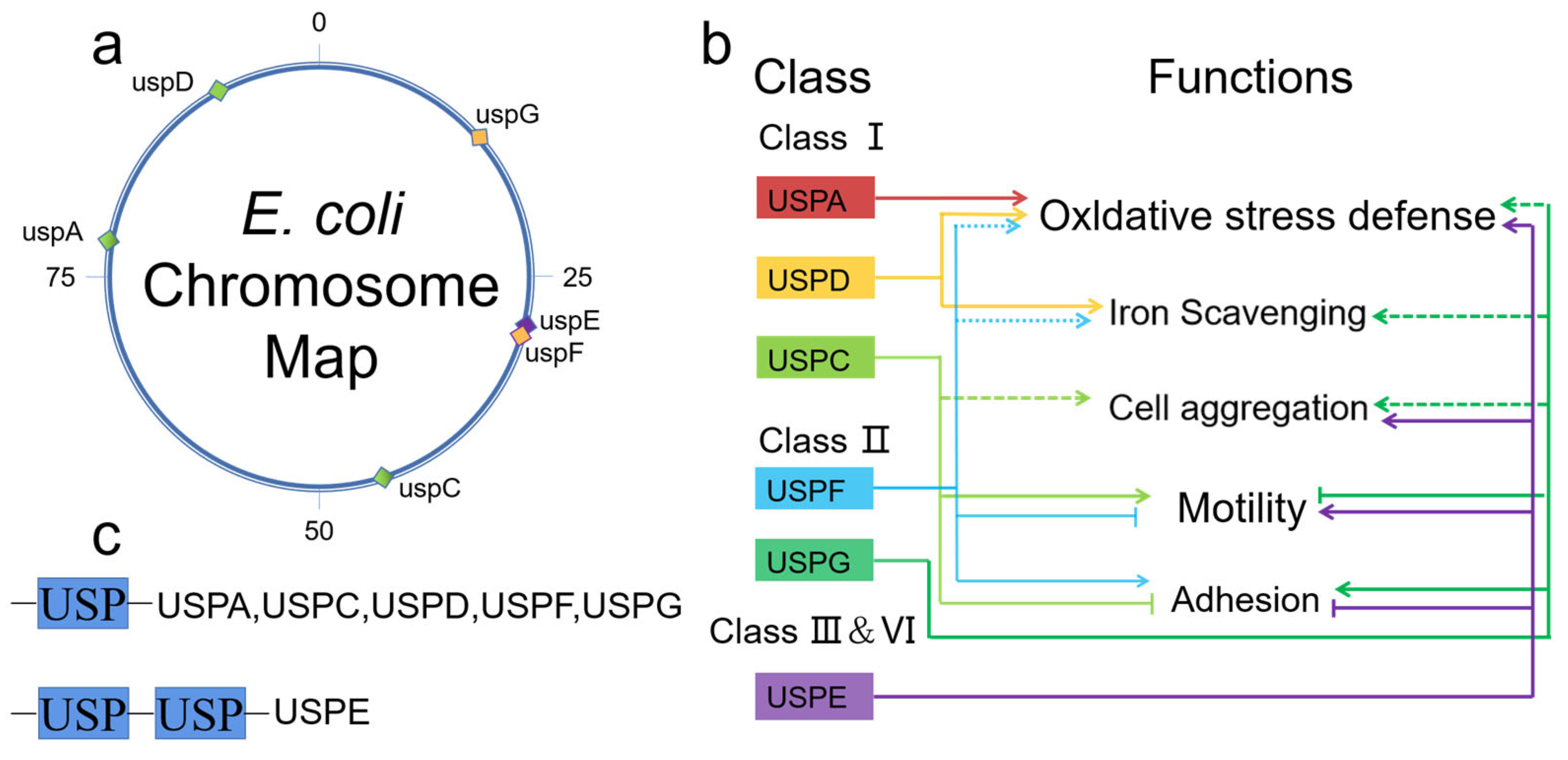
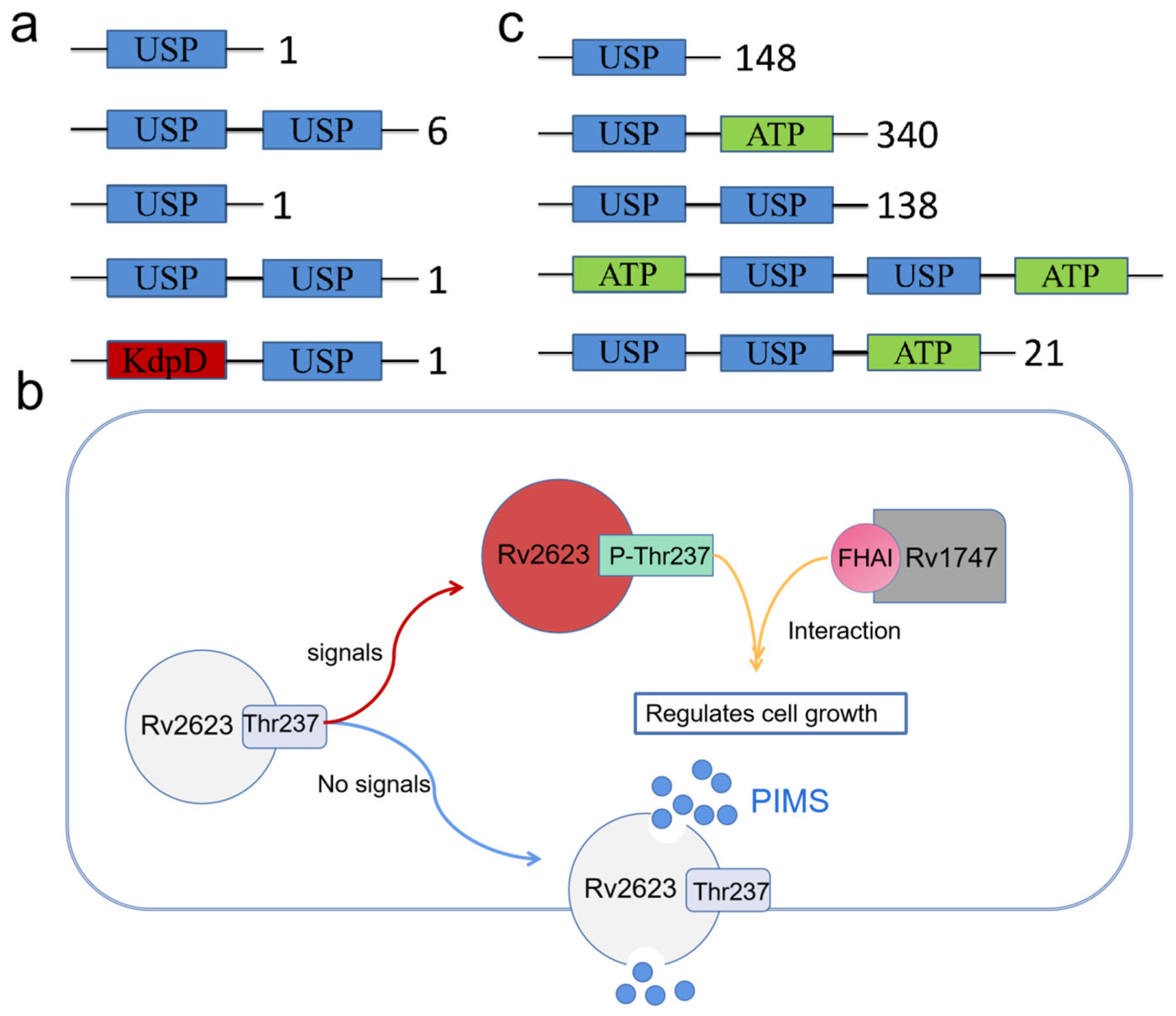
2. Diversity of USP Genes
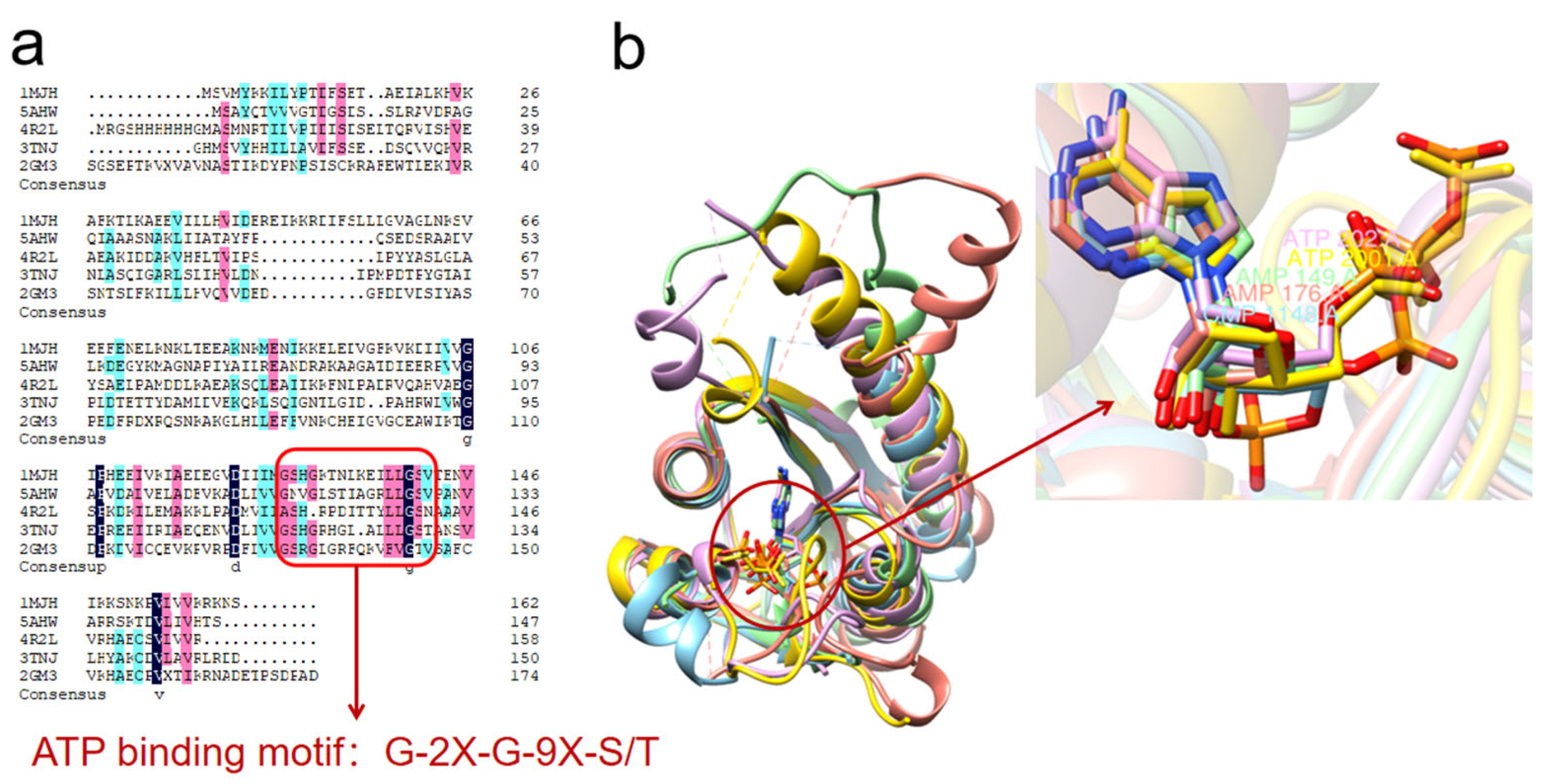
3. USP Crystal Structure
4. Functional Diversity of USPs
4.1. Functional Diversity of Prokaryotic USPs
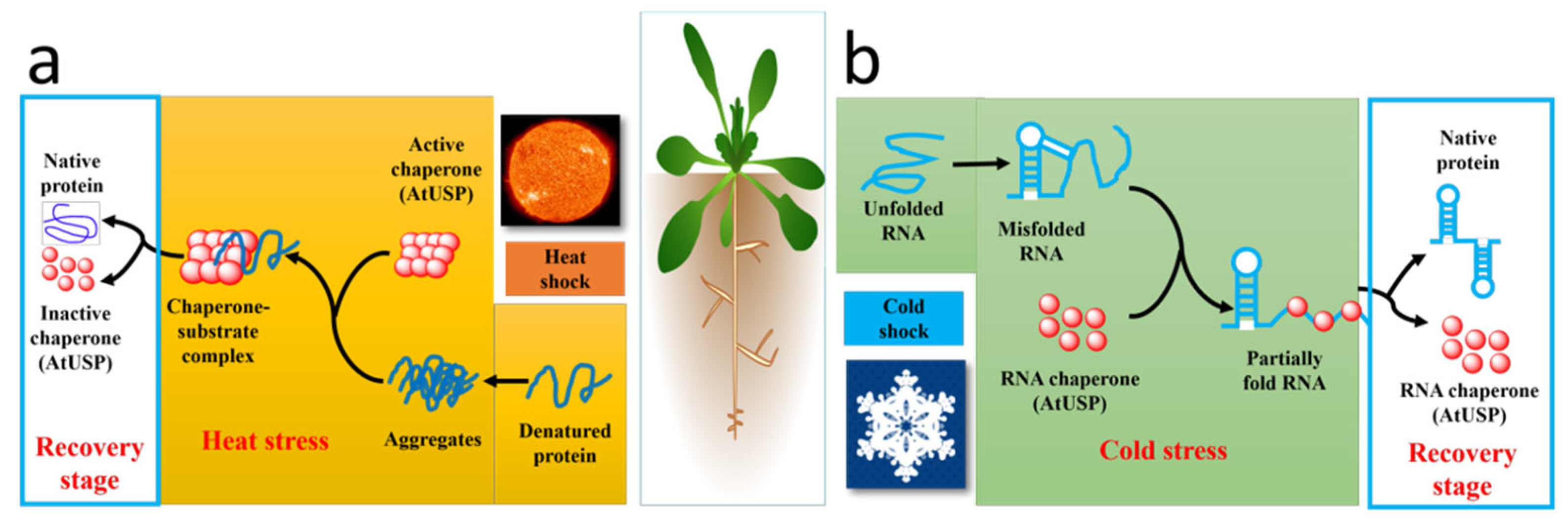
4.2. Functional Diversity of Eukaryotic USPs
4.2.1. Versatility of A. thaliana USPs
4.2.2. Functional Diversity of USPs in Other Plants
5. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nyström, T.; Neidhardt, F.C. Cloning, mapping and nucleotide sequencing of a gene encoding a universal stress protein in Eschericha coli. Mol. Microbiol. 1992, 6, 3187–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyström, T.; Neidhardt, F.C. Isolation and properties of a mutant of Escherichia coli with an insertional inactivation of the uspA gene, which encodes a universal stress protein. J. Bacteriol. 1993, 175, 3949–3956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyström, T.; Neidhardt, F.C. Expression and role of the universal stress protein, UspA, of Escherichia coli during growth arrest. Mol. Microbiol. 1994, 11, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freestone, P.; Nystrom, T.; Trinei, M.; Norris, V. The universal stress protein, UspA, of Escherichia coli is phosphorylated in response to stasis. J. Mol. Biol. 1997, 274, 318–324. [Google Scholar] [CrossRef]
- Freestone, P.; Grant, S.; Trinei, M.; Onoda, T.; Norris, V. Protein phosphorylation in Escherichia cob L. form NC=7. Microbiology 1998, 144, 3289–3295. [Google Scholar] [CrossRef] [Green Version]
- Phadtare, S.; Inouye, M. Role of CspC and CspE in regulation of expression of RpoS and UspA, the stress response proteins in Escherichia coli. J. Bacteriol. 2001, 183, 1205–1214. [Google Scholar] [CrossRef] [Green Version]
- Tao, H.; Bausch, C.; Richmond, C.; Blattner, F.R.; Conway, T. Functional genomics: Expression analysis of Escherichia coli growing on minimal and rich media. J. Bacteriol. 1999, 181, 6425–6440. [Google Scholar] [CrossRef] [Green Version]
- Forêt, S.; Seneca, F.; De Jong, D.; Bieller, A.; Hemmrich, G.; Augustin, R.; Hayward, D.C.; Ball, E.E.; Bosch, T.C.; Agata, K. Phylogenomics reveals an anomalous distribution of USP genes in metazoans. Mol. Biol. Evol. 2011, 28, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Kvint, K.; Nachin, L.; Diez, A.; Nyström, T. The bacterial universal stress protein: Function and regulation. Curr. Opin. Microbiol. 2003, 6, 140–145. [Google Scholar] [CrossRef]
- Vollmer, A.C.; Bark, S.J. Twenty five years of investigating the universal stress protein: Function, structure, and applications. Adv. Appl. Microbiol. 2018, 102, 1–36. [Google Scholar]
- Gustavsson, N.; Diez, A.; Nyström, T. The universal stress protein paralogues of Escherichia coli are coordinately regulated and cooperate in the defence against DNA damage. Mol. Microbiol. 2002, 43, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Anantharaman, V.; Koonin, E.V. Monophyly of class I aminoacyl tRNA synthetase, USPA, ETFP, photolyase, and PP-ATPase nucleotide binding domains: Implications for protein evolution in the RNA world. Proteins Struct. Funct. Bioinform. 2002, 48, 1–14. [Google Scholar] [CrossRef]
- Nachin, L.; Nannmark, U.; Nystrom, T. Differential roles of the universal stress proteins of Escherichia coli in oxidative stress resistance, adhesion, and motility. J. Bacteriol. 2005, 187, 6265–6272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegele, D.A. Universal stress proteins in Escherichia coli. J. Bacteriol. 2005, 187, 6253–6254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boes, N.; Schreiber, K.; Härtig, E.; Jaensch, L.; Schobert, M. The Pseudomonas aeruginosa universal stress protein PA4352 is essential for surviving anaerobic energy stress. J. Bacteriol. 2006, 188, 6529–6538. [Google Scholar] [CrossRef] [Green Version]
- Chou, M.X.; Wei, X.Y.; Chen, D.S.; Zhou, J.C. A novel nodule-enhanced gene encoding a putative universal stress protein from Astragalus sinicus. J. Plant Physiol. 2007, 164, 764–772. [Google Scholar] [CrossRef]
- Galindo Blaha, C.A.; Schrank, I.S. An Azospirillum brasilense Tn5 mutant with modified stress response and impaired in flocculation. Antonie Van Leeuwenhoek 2003, 83, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Fink, R.C.; Evans, M.R.; Porwollik, S.; Vazquez-Torres, A.; Jones-Carson, J.; Troxell, B.; Libby, S.J.; McClelland, M.; Hassan, H.M. FNR is a global regulator of virulence and anaerobic metabolism in Salmonella enterica serovar Typhimurium (ATCC 14028s). J. Bacteriol. 2007, 189, 2262–2273. [Google Scholar] [CrossRef] [Green Version]
- Sauter, M.; Rzewuski, G.; Marwedel, T.; Lorbiecke, R. The novel ethylene regulated gene OsUsp1 from rice encodes a member of a plant protein family related to prokaryotic universal stress proteins. J. Exp. Bot. 2002, 53, 2325–2331. [Google Scholar] [CrossRef] [Green Version]
- Coetzer, N.; Gazendam, I.; Oelofse, D.; Berger, D.K. SSHscreen and SSHdb, generic software for microarray based gene discovery: Application to the stress response in cowpea. Plant Methods 2010, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Kerk, D.; Bulgrien, J.; Smith, D.W.; Gribskov, M. Arabidopsis proteins containing similarity to the universal stress protein domain of bacteria. Plant Physiol. 2003, 131, 1209–1219. [Google Scholar] [CrossRef] [Green Version]
- Merkouropoulos, G.; Andreasson, E.; Hess, D.; Boller, T.; Peck, S.C. An Arabidopsis protein phosphorylated in response to microbial elicitation, AtPHOS32, is a substrate of MAP kinases 3 and 6. J. Biol. Chem. 2008, 283, 10493–10499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maqbool, A.; Zahur, M.; Irfan, M.; Younas, M.; Barozai, K.; Rashid, B.; Husnain, T.; Riazuddin, S. Identification and expression of six drought-responsive transcripts through differential display in desi cotton (Gossypium arboreum). Mol. Biol. 2008, 42, 492–498. [Google Scholar] [CrossRef]
- Wang, H. Construction and Analysis of subtractive cDNA library of phellodendron amurense under drought stress. Chin. J. Biotechnol. 2008, 24, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Sapitnitskaya, M.; Maul, P.; McCollum, G.T.; Guy, C.L.; Weiss, B.; Samach, A.; Porat, R. Postharvest heat and conditioning treatments activate different molecular responses and reduce chilling injuries in grapefruit. J. Exp. Bot. 2006, 57, 2943–2953. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.A.; Raymond, M.J.; Smirnoff, N. Analysis of the root-hair morphogenesis transcriptome reveals the molecular identity of six genes with roles in root-hair development in Arabidopsis. Plant J. 2006, 45, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Drumm, J.E.; Mi, K.; Bilder, P.; Sun, M.; Lim, J.; Bielefeldt-Ohmann, H.; Basaraba, R.; So, M.; Zhu, G.; Tufariello, J.M. Mycobacterium tuberculosis universal stress protein Rv2623 regulates bacillary growth by ATP-Binding: Requirement for establishing chronic persistent infection. PLoS Pathog. 2009, 5, e1000460. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, M.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, S.; Rai, R.; Chatterjee, A.; Rai, S.; Yadav, S.; Agrawal, C.; Rai, L. Molecular characterization of two novel proteins All1122 and Alr0750 of Anabaena PCC 7120 conferring tolerance to multiple abiotic stresses in Escherichia coli. Gene 2019, 685, 230–241. [Google Scholar] [CrossRef]
- Tkaczuk, K.L.; Shumilin, I.A.; Chruszcz, M.; Evdokimova, E.; Savchenko, A.; Minor, W. Structural and functional insight into the universal stress protein family. Evol. Appl. 2013, 6, 434–449. [Google Scholar] [CrossRef]
- Elhosseiny, N.M.; Amin, M.A.; Yassin, A.S.; Attia, A.S. Acinetobacter baumannii universal stress protein A plays a pivotal role in stress response and is essential for pneumonia and sepsis pathogenesis. Int. J. Med. Microbiol. 2015, 305, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Isokpehi, R.D.; Mahmud, O.; Mbah, A.N.; Simmons, S.S.; Brown, S.D. Developmental regulation of genes encoding universal stress proteins in Schistosoma mansoni. Gene Regul. Syst. Biol. 2011, 5, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinola, S.M.; Cancela, M.P.; Corrêa, L.B.; Zaha, A. Evolutionary fates of universal stress protein paralogs in Platyhelminthes. BMC Evol. Biol. 2018, 18, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rekadwad, B.N.; Li, W.J.; Rekha, P.D. The diversity of unique 1,4,5,6-Tetrahydro-2-methyl-4-pyrimidinecarboxylic acid coding common genes and Universal stress protein in Ectoine TRAP cluster (UspA) in 32 Halomonas species. BMC Res. Notes 2021, 14, 296. [Google Scholar] [CrossRef] [PubMed]
- Bhuria, M.; Goel, P.; Kumar, S.; Singh, A.K. Genome-wide identification and expression profiling of genes encoding universal stress proteins (USP) identify multi-stress responsive USP genes in Arabidopsis thaliana. Plant Physiol. Rep. 2019, 24, 434–445. [Google Scholar] [CrossRef]
- Gorshkova, D.; Getman, I.; Voronkov, A.; Chizhova, S.; Kuznetsov, V.V.; Pojidaeva, E. The gene encoding the universal stress protein AtUSP is regulated by phytohormones and involved in seed germination of Arabidopsis thaliana. Dokl. Biochem. Biophys. 2018, 479, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Li, W.T.; Wei, Y.M.; Wang, J.R.; Liu, C.J.; Lan, X.J.; Jiang, Q.T.; Pu, Z.E.; Zheng, Y.L. Identification, localization, and characterization of putative USP genes in barley. Theor. Appl. Genet. 2010, 121, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.S.; Phan, K.A.T.; Jun, S.E.; Park, J.H.; Paeng, S.K.; Chae, H.B.; Wi, S.D.; Bae, S.B.; Kang, K.R.; Kim, G.T.; et al. Universal Stress Protein (USP) enhances plant growth and development by promoting cell expansion. J. Plant Biol. 2022, 65, 231–239. [Google Scholar] [CrossRef]
- Bhuria, M.; Goel, P.; Kumar, S.; Singh, A.K. The promoter of AtUSP is coregulated by phytohormones and abiotic stresses in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1957. [Google Scholar] [CrossRef] [Green Version]
- Gorshkova, D.; Getman, I.; Sergeeva, L.; Kuznetsov, V.V.; Pojidaeva, E. GRUSP, an universal stress protein, is involved in gibberellin-dependent induction of flowering in Arabidopsis thaliana. Dokl. Biochem. Biophys. 2021, 499, 233–237. [Google Scholar] [CrossRef]
- Bhuria, M.; Goel, P.; Kumar, S.; Singh, A.K. AtUSP17 negatively regulates salt stress tolerance through modulation of multiple signaling pathways in Arabidopsis. Physiol. Plant. 2022, 174, e13635. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Zhang, P.; Hu, Y.; Chen, C.; Liu, Q.; Guan, P.; Zhang, J. Genome-wide analysis of the universal stress protein A gene family in Vitis and expression in response to abiotic stress. Plant Physiol. Biochem. 2021, 165, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Biswas, P.; Banerjee, A.; Bose, A.; Siddiqui, N.; Nambi, S.; Saini, D.K.; Visweswariah, S.S. A universal stress protein in Mycobacterium smegmatis sequesters the cAMP-regulated lysine acyltransferase and is essential for biofilm formation. J. Biol. Chem. 2020, 295, 1500–1516. [Google Scholar] [CrossRef] [Green Version]
- Loukehaich, R.; Wang, T.; Ouyang, B.; Ziaf, K.; Li, H.; Zhang, J.; Lu, Y.; Ye, Z. SpUSP, an annexin-interacting universal stress protein, enhances drought tolerance in tomato. J. Exp. Bot. 2012, 63, 5593–5606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arabia, S.; Sami, A.A.; Akhter, S.; Sarker, R.H.; Islam, T. Comprehensive in silico characterization of universal stress proteins in rice (Oryza sativa L.) with insight into their stress-specific transcriptional modulation. Front. Plant Sci. 2021, 12, 712607. [Google Scholar] [CrossRef]
- Yang, M.; Che, S.; Zhang, Y.; Wang, H.; Wei, T.; Yan, G.; Song, W.; Yu, W. Universal stress protein in Malus sieversii confers enhanced drought tolerance. J. Plant Res. 2019, 132, 825–837. [Google Scholar] [CrossRef]
- Varshney, R.K.; Chen, W.; Li, Y.; Bharti, A.K.; Saxena, R.K.; Schlueter, J.A.; Donoghue, M.T.; Azam, S.; Fan, G.; Whaley, A.M. Draft genome sequence of pigeonpea (Cajanus cajan), an orphan legume crop of resource-poor farmers. Nat. Biotechnol. 2012, 30, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Sinha, P.; Pazhamala, L.T.; Singh, V.K.; Saxena, R.K.; Krishnamurthy, L.; Azam, S.; Khan, A.W.; Varshney, R.K. Identification and validation of selected universal stress protein domain containing drought responsive genes in pigeonpea (Cajanus cajan L.). Front. Plant Sci. 2016, 6, 1065. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.; Ahmad, A.; Batool, F.; Rashid, B.; Husnain, T. Genetic modification of Gossypium arboreum universal stress protein (GUSP1) improves drought tolerance in transgenic cotton (Gossypium hirsutum). Physiol. Mol. Biol. Plants 2021, 27, 1779–1794. [Google Scholar] [CrossRef]
- Hassan, S.; Samiullah, T.R.; Ansari, M.R.; Rashid, B.; Husnain, T. Cloning, antibody production, expression and cellular localization of universal stress protein gene (USP1-GFP) in transgenic cotton. J. Plant Biochem. Biotechnol. 2018, 27, 175–185. [Google Scholar] [CrossRef]
- Hafeez, M.N.; Khan, M.A.; Sarwar, B.; Hassan, S.; Ali, Q.; Husnain, T.; Rashid, B. Mutant Gossypium universal stress protein-2 (GUSP-2) gene confers resistance to various abiotic stresses in E. coli BL-21 and CIM-496-Gossypium hirsutum. Sci. Rep. 2021, 11, 20466. [Google Scholar] [CrossRef] [PubMed]
- Udawat, P.; Jha, R.K.; Sinha, D.; Mishra, A.; Jha, B. Overexpression of a cytosolic abiotic stress responsive universal stress protein (SbUSP) mitigates salt and osmotic stress in transgenic tobacco plants. Front. Plant Sci. 2016, 7, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udawat, P.; Mishra, A.; Jha, B. Heterologous expression of an uncharacterized universal stress protein gene (SbUSP) from the extreme halophyte, Salicornia brachiata, which confers salt and osmotic tolerance to E. coli. Gene 2014, 536, 163–170. [Google Scholar] [CrossRef]
- Shokry, A.M.; Karim, S.A.; Ramadan, A.; Gadallah, N.; Attas, S.G.; Sabir, J.S.; Hassan, S.M.; Madkour, M.A.; Bressan, R.; Mahfouz, M. Detection of a Usp-like gene in Calotropis procera plant from the de novo assembled genome contigs of the high-throughput sequencing dataset. C. R. Biol. 2014, 337, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.F.; Su, J.; Na, Y.; Hui, Z.; Cao, X.Y.; Kang, J.F. Functional characterization of selected universal stress protein from Salvia miltiorrhiza (SmUSP) in Escherichia coli. Genes 2017, 8, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanyalakshmi, K.H.; Nataraja, K.N. Universal stress protein-like gene from mulberry enhances abiotic stress tolerance in Escherichia coli and transgenic tobacco cells. Plant Biol. 2021, 23, 1190–1194. [Google Scholar] [CrossRef]
- Zarembinski, T.I.; Hung, L.W.; Mueller, H.J.; Kim, K.K.; Yokota, H.; Kim, R.; Kim, S.H. Structure-based assignment of the biochemical function of a hypothetical protein: A test case of structural genomics. Proc. Natl. Acad. Sci. USA 1998, 95, 15189–15193. [Google Scholar] [CrossRef] [Green Version]
- Sousa, M.C.; McKay, D.B. Structure of the Universal Stress Protein of Haemophilus influenzae. Structure 2001, 9, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Adolph, R.S.; Gopalakrishnapai, J.; Kleinboelting, S.; Emmerich, C.; Steegborn, C.; Visweswariah, S.S. A universal stress protein (USP) in Mycobacteria binds cAMP. J. Biol. Chem. 2015, 290, 12731–12743. [Google Scholar] [CrossRef] [Green Version]
- Bangera, M.; Panigrahi, R.; Sagurthi, S.R.; Savithri, H.S.; Murthy, M. Structural and functional analysis of two universal stress proteins YdaA and YnaF from Salmonella typhimurium: Possible roles in microbial stress tolerance. J. Struct. Biol. 2015, 189, 238–250. [Google Scholar] [CrossRef]
- Dutta, A.; Batish, M.; Parashar, V. Structural basis of KdpD histidine kinase binding to the second messenger c-di-AMP. J. Biol. Chem. 2021, 296, 100771. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Quan, C.-S.; Jin, X.; Zhao, J.; Li, X.; Zheng, W.; Jin, L.; Liu, D.; Fan, S.; Ha, N.-C. Crystallization and preliminary X-ray diffraction analysis of UspE from Escherichia coli. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2014, 70, 1640–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Guo, J.; Jin, X.; Kim, J.S.; Ji, Y.; Fan, S.; Ha, N.C.; Quan, C.S. Crystal structure and functional implications of the tandem-type universal stress protein UspE from Escherichia coli. BMC Struct. Biol. 2016, 16, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.; Eduard, K.; Craig, B.; Bingman, A.; Kim, H.J. Crystal structure of the protein At3g01520, a eukaryotic universal stress protein-like protein from Arabidopsis thalianain complex with AMP. Proteins Struct. Funct. Bioinform. 2015, 83, 1368–1373. [Google Scholar]
- Havis, S.; Bodunrin, A.; Rangel, J.; Zimmerer, R.; Murphy, J.; Storey, J.D.; Duong, T.D.; Mistretta, B.; Gunaratne, P.; Widger, W.R. A universal stress protein that controls bacterial stress survival in Micrococcus luteus. J. Bacteriol. 2019, 201, e00497-19. [Google Scholar] [CrossRef]
- Iino, H.; Shimizu, N.; Goto, M.; Ebihara, A.; Fukui, K.; Hirotsu, K.; Kuramitsu, S. Crystal structure of the tandem-type universal stress protein TTHA0350 from Thermus thermophilus HB8. J. Biochem. 2011, 150, 295–302. [Google Scholar] [CrossRef]
- Averaimo, S.; Milton, R.H.; Duchen, M.R.; Mazzanti, M. Chloride intracellular channel 1 (CLIC1): Sensor and effector during oxidative stress. FEBS Lett. 2010, 584, 2076–2084. [Google Scholar] [CrossRef] [Green Version]
- Suh, K.S.; Mutoh, M.; Nagashima, K.; Fernandez-Salas, E.; Edwards, L.E.; Hayes, D.D.; Crutchley, J.M.; Marin, K.G.; Dumont, R.A.; Levy, J.M. The organellular chloride channel protein CLIC4/mtCLIC translocates to the nucleus in response to cellular stress and accelerates apoptosis. J. Biol. Chem. 2004, 279, 4632–4641. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, R.; Williams, H.D. Universal stress proteins and Mycobacterium tuberculosis. Res. Microbiol. 2003, 154, 387–392. [Google Scholar] [CrossRef]
- Chakraborti, S.; Chakraborty, M.; Bose, A.; Srinivasan, N.; Visweswariah, S.S. Identification of potential binders of Mtb universal stress protein (Rv1636) through an in silico approach and insights into compound selection for experimental validation. Front. Mol. Biosci. 2021, 8, 177. [Google Scholar] [CrossRef]
- Glass, L.N.; Swapna, G.; Chavadi, S.S.; Tufariello, J.M.; Mi, K.; Drumm, J.E.; Lam, T.T.; Zhu, G.; Zhan, C.; Vilcheze, C. Mycobacterium tuberculosis universal stress protein Rv2623 interacts with the putative ATP binding cassette (ABC) transporter Rv1747 to regulate mycobacterial growth. PLoS Pathog. 2017, 13, e1006515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Q.; Hu, X.; Shi, D.; Zhang, Y.; Sun, M.; Wang, J.; Mi, K.; Zhu, G. Universal stress protein Rv2624c alters abundance of arginine and enhances intracellular survival by ATP binding in mycobacteria. Sci. Rep. 2016, 6, 35462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Li, X.; Huang, L.; Chan, J.; Chen, Y.; Deng, H.; Mi, K. Quantitative proteomics reveals novel insights into isoniazid susceptibility in mycobacteria mediated by a universal stress protein. J. Proteome Res. 2015, 14, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Isokpehi, R.D.; McInnis, D.S.; Destefano, A.M.; Johnson, G.S.; Walker, A.D.; Hall, Y.A.; Mapp, B.W.; Johnson, M.O.; Simmons, S.S. Bioinformatics investigations of universal stressproteins from mercury methylating desulfovibrionaceae. Microorganisms 2021, 9, 1780. [Google Scholar] [CrossRef]
- O’Connor, A.; Berisio, R.; Lucey, M.; Schaffer, K.; Clean, S.M. A Universal Stress Protein upregulated by hypoxia may contribute to chronic lung colonisation and intramacrophage survival in cystic fibrosis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Tremonte, P.; Succi, M.; Coppola, R.; Sorrentino, E.; Tipaldi, L.; Picariello, G.; Pannella, G.; Fraternali, F. Homology-based modeling of universal stress protein from Listeria innocua up-regulated under acid stress conditions. Front. Microbiol. 2016, 7, 1998. [Google Scholar] [CrossRef]
- De Souza, C.S.; Torres, A.G.; Caravelli, A.; Silva, A.; Polatto, J.M.; Piazza, R.M.F. Characterization of the universal stress protein F from atypical enteropathogenic Escherichia coli and its prevalence in Enterobacteriaceae. Protein Sci. 2016, 25, 2142–2151. [Google Scholar] [CrossRef] [Green Version]
- Masamba, P.; Kappo, A.P. Parasite survival and disease persistence in cystic fibrosis, schistosomiasis and pathogenic bacterial diseases: A role for universal Stress Proteins? Int. J. Mol. Sci. 2021, 22, 10878. [Google Scholar] [CrossRef]
- Bandyopadhyay, D.; Singh, G.; Mukherjee, M.; Akhter, Y. Computational approach towards the design of novel inhibitor against universal stress protein A to combat multidrug resistant uropathogenic Escherichia coli. J. Mol. Struct. 2021, 1238, 130379. [Google Scholar] [CrossRef]
- Fang, Q.j.; Han, Y.X.; Shi, Y.J.; Huang, H.Q.; Fang, Z.G.; Hu, Y.H. Universal stress proteins contribute Edwardsiella piscicida adversity resistance and pathogenicity and promote blocking host immune response. Fish Shellfish. Immunol. 2019, 95, 248–258. [Google Scholar] [CrossRef]
- Heermann, R.; Weber, A.; Mayer, B.; Ott, M.; Hauser, E.; Gabriel, G.; Pirch, T.; Jung, K. The universal stress protein UspC scaffolds the KdpD/KdpE signaling cascade of Escherichia coli under salt stress. J. Mol. Biol. 2009, 386, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Heermann, R.; Jung, K. The complexity of the ‘simple’two-component system KdpD/KdpE in Escherichia coli. FEMS Microbiol. Lett. 2010, 304, 97–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isokpehi, R.D.; Mahmud, O.; Awofolu, O.R.; Mbah, A.N. Inferences on the biochemical and environmental regulation of Universal Stress Proteins from Schistosomiasis parasites. Adv. Appl. Bioinform. Chem. 2013, 6, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masamba, P.; Adenowo, A.F.; Oyinloye, B.E.; Kappo, A.P. Universal stress proteins as new targets for environmental and therapeutic interventions of Schistosomiasis. Int. J. Environ. Res. Public Health 2016, 13, 972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.J.; Melencion, S.M.; Lee, E.S.; Park, J.H.; Alinapon, C.V.; Oh, H.T.; Yun, D.J.; Chi, Y.H.; Lee, S.Y. Universal stress protein exhibits a redox-dependent chaperone function in arabidopsis and enhances plant tolerance to heat shock and oxidative stress. Front. Plant Sci. 2015, 6, 1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verslues, P.E.; Lasky, J.R.; Juenger, T.E.; Liu, T.W.; Kumar, M.N. Genome-wide association mapping combined with reverse genetics identifies new effectors of low water potential-induced proline accumulation in Arabidopsis. Plant Physiol. 2014, 164, 144–159. [Google Scholar] [CrossRef] [Green Version]
- Gonzali, S.; Loreti, E.; Cardarelli, F.; Novi, G.; Parlanti, S.; Pucciariello, C.; Bassolino, L.; Banti, V.; Licausi, F.; Perata, P. Universal stress protein HRU1 mediates ROS homeostasis under anoxia. Nat. Plants 2015, 1, 15151. [Google Scholar] [CrossRef] [PubMed]
- Melencion, S.M.B.; Chi, Y.H.; Pham, T.T.; Paeng, S.K.; Wi, S.D.; Lee, C.; Ryu, S.W.; Koo, S.S.; Lee, S.Y. RNA chaperone function of a universal stress protein in Arabidopsis confers enhanced cold stress tolerance in plants. Int. J. Mol. Sci. 2017, 18, 2546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Park, S.J.; Kwak, K.J. Plant RNA chaperones in stress response. Trends Plant Sci. 2013, 18, 100–106. [Google Scholar] [CrossRef]
- Chi, Y.H.; Koo, S.S.; Oh, H.T.; Lee, E.S.; Park, J.H.; Phan, K.A.T.; Wi, S.D.; Bae, S.B.; Paeng, S.K.; Chae, H.B. The physiological functions of universal stress proteins and their molecular mechanism to protect plants from environmental stresses. Front. Plant Sci. 2019, 10, 750. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.H.; Paeng, S.K.; Kim, M.J.; Hwang, G.Y.; Melencion, S.M.; Oh, H.T.; Lee, S.Y. Redox-dependent functional switching of plant proteins accompanying with their structural changes. Front. Plant Sci. 2013, 4, 277. [Google Scholar] [CrossRef] [Green Version]
- Park, S.C.; Jung, Y.J.; Lee, Y.; Kim, I.R.; Seol, M.-A.; Kim, E.-J.; Jang, M.-K.; Lee, J.R. Functional characterization of the Arabidopsis universal stress protein AtUSP with an antifungal activity. Biochem. Biophys. Res. Commun. 2017, 486, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Geigenberger, P.; Thormählen, I.; Daloso, D.M.; Fernie, A.R. The unprecedented versatility of the plant thioredoxin system. Trends Plant Sci. 2017, 22, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Bosch, C.G. Priming plant resistance by activation of redox-sensitive genes. Free. Radic. Biol. Med. 2018, 122, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 8, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Mata-Pérez, C.; Spoel, S.H. Thioredoxin-mediated redox signalling in plant immunity. Plant Sci. 2019, 279, 27–33. [Google Scholar] [CrossRef]
- Hinoux, V.D.; Bangash, S.A.; Meyer, A.J.; Reichheld, J.P. Nuclear thiol redox systems in plants. Plant Sci. 2016, 243, 84–95. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Gutsche, N.; Thurow, C.; Zachgo, S.; Gatz, C. Plant specific CC-type glutaredoxins: Functions in developmental processes and stress responses. Biol. Chem. 2015, 396, 495–509. [Google Scholar] [CrossRef]
- Nikkanen, L.; Toivola, J.; Diaz, M.G.; Rintamäki, E. Chloroplast thioredoxin systems: Prospects for improving photosynthesis. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160474. [Google Scholar] [CrossRef] [Green Version]
- Beltrán, E.G.; Personat, J.M.; Torre, F.D.; Pozo, O.D. A universal stress protein involved in oxidative stress is a phosphorylation target for protein kinase CIPK6. Plant Physiol. 2017, 173, 836–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Does, C.V.; Albers, S.V. SaUspA, the universal stress protein of Sulfolobus acidocaldarius stimulates the activity of the PP2A phosphatase and is involved in growth at high salinity. Front. Microbiol. 2020, 11, 2872. [Google Scholar] [CrossRef] [PubMed]
- Moghanloo, M.; Iranbakhsh, A.; Ebadi, M.; Ardebili, Z.O. Differential physiology and expression of phenylalanine ammonia lyase (PAL) and universal stress protein (USP) in the endangered species Astragalus fridae following seed priming with cold plasma and manipulation of culture medium with silica nanoparticles. 3 Biotech 2019, 9, 288. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, D.; Mukherjee, M. Systematic comparison of the protein-protein interaction network of bacterial universal stress protein A (UspA): An insight into its discrete functions. Biologia 2022, 77, 2631–2642. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Protein | Locus | CDS Length (bp) | Protein Length (aa) | Domain | Pfam ID |
|---|---|---|---|---|---|---|
| AtUSP1 | AtUSP1 | At1G01680 | 927 | 308 | USP | PF00582 |
| AtUSP2 | AtUSP2 | At1G09740 | 516 | 171 | USP | PF00582 |
| AtUSP3 | AtUSP3 | At1G11360 | 729 | 242 | USP | PF00582 |
| AtUtyK1 | AtUtyK1 | At1G16760 | 2277 | 758 | USP | PF00582 |
| Pkinase-Tyr | PF07714 | |||||
| AtUtyK2 | AtUtyK2 | At1G17540 | 2187 | 728 | USP | PF00582 |
| Pkinase-Tyr | PF00069 | |||||
| AtUK1 | AtUK1 | At1G21590 | 2271 | 756 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP4 | AtUSP4 | At1G44760 | 642 | 213 | USP | PF00582 |
| AtUSP5 | AtUSP5 | At1G48960 | 660 | 219 | USP | PF00582 |
| AtUSP6 | AtUSP6 | At1G68300 | 483 | 160 | USP | PF00582 |
| AtUSP7 | AtUSP7.1 | At1G69080.1 | 672 | 223 | USP | PF00582 |
| AtUSP7.2 | At1G69080.2 | 630 | 209 | USP | PF00582 | |
| AtUK2 | AtUK2 | At1G77280 | 2385 | 753 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUtyK3 | AtUtyK3 | At1G78940.2 | 2265 | 754 | USP | PF00582 |
| Pkinase-Tyr | PF07714 | |||||
| AtUSP8 | AtUSP8 | At2G03720 | 498 | 165 | USP | PF00582 |
| AtUK3 | AtUK3 | At2G07020 | 2103 | 700 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP9 | AtUSP9.1 | At2G21620.1 | 564 | 187 | USP | PF00582 |
| AtUSP9.2 | At2G21620.2 | 582 | 193 | USP | PF00582 | |
| AtUK4 | AtUK4 | At2G24370 | 2367 | 788 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP10 | AtUSP10 | At2G47710 | 489 | 162 | USP | PF00582 |
| AtUSP11 | AtUSP11 | At3G01520 | 528 | 175 | USP | PF00582 |
| AtUSP12 | AtUSP12 | At3G03270.1 | 606 | 201 | USP | PF00582 |
| AtUSP13 | AtUSP13 | At3G03290 | 825 | 274 | USP | PF00582 |
| AtUSPUSP1 | AtUSPUSP1.1 | At3G11930.1 | 600 | 199 | USP | PF00582 |
| AtUSPUSP1.2 | At3G11930.2 | 603 | 200 | USP | PF00582 | |
| AtUSPUSP1.3 | At3G11930.3 | 681 | 226 | USP, USP | PF00582 | |
| AtUSPUSP1.4 | At3G11930.4 | 606 | 201 | USP | PF00582 | |
| AtUSP14 | AtUSP14 | At3G17020 | 492 | 163 | USP | PF00582 |
| AtUtyK4 | AtUtyK4 | At3G20200 | 2343 | 780 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP15 | AtUSP15 | At3G21210 | 2415 | 804 | USP | PF00582 |
| AtUSP16 | AtUSP16 | At3G25930 | 465 | 154 | USP | PF00582 |
| AtUSP17 | AtUSP17.1 | At3G53990.1 | 483 | 160 | USP | PF00582 |
| AtUSP17.2 | At3G53990.2 | 381 | 126 | USP | PF00582 | |
| AtUSP18 | AtUSP18.1 | At3G58450.1 | 615 | 204 | USP | PF00582 |
| AtUSP18.2 | At3G58450.2 | 594 | 197 | USP | PF00582 | |
| AtUSP19 | AtUSP19 | At3G62550 | 489 | 162 | USP | PF00582 |
| AtUtyK5 | AtUtyK5 | At4G25160 | 2508 | 835 | USP | PF00582 |
| Pkinase-Tyr | PF07714 | |||||
| AtUSP20 | AtUSP20 | AT4G13450.1 | 660 | 219 | USP | PF00582 |
| AtUSP21 | AtUSP21 | At4G27320 | 783 | 260 | USP | PF00582 |
| AtUK5 | AtUK5 | At4G31230 | 2295 | 764 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUtyK6 | AtUtyK6 | At5G12000 | 2106 | 701 | USP | PF00582 |
| Pkinase-Tyr | PF07714 | |||||
| AtUSP22 | AtUSP22 | At5G14680 | 528 | 175 | USP | PF00582 |
| AtUSP23 | AtUSP23 | At5G17390 | 858 | 285 | USP | PF00582 |
| AtUSP24 | AtUSP24 | At5G20310 | 1185 | 394 | USP | PF00582 |
| AtUK6 | AtUK6 | At5G35380 | 2196 | 731 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP25 | AtUSP25.1 | At5G47740.1 | 735 | 244 | USP | PF00582 |
| AtUSP25.2 | At5G47740.2 | 741 | 246 | USP | PF00582 | |
| AtUSP26 | AtUSP26 | At5G49050 | 453 | 150 | USP | PF00582 |
| AtUSP27 | AtUSP27 | At5G54430 | 729 | 242 | USP | PF00582 |
| AtUK7 | AtUK7 | At5G63940 | 2118 | 705 | USP | PF00582 |
| Pkinase | PF00069 |
| Organism | PDB Code | Chains | Mutation(s) | Small Molecules | References |
|---|---|---|---|---|---|
| Methanocaldococcus jannaschii | 1MJH | 2 | 0 | ATP, Mn2+ | [57] |
| Haemophilus influenzae | 1JMV | 4 | 0 | SO42− | [58] |
| Methanocaldococcus jannaschii | 5AHW | 6 | 0 | CMP, SO42−, POG, Cl− | [59] |
| Salmonella enterica subsp. enterica serovar Typhimurium str. LT2 | 4R2J | 1 | 0 | GLC, PO43−, EDO, Zn2+ | [60] |
| 4R2K | 1 | 1 (4R2J Mutation) | SO42−, EDO, OXD | ||
| 4R2L | 2 | 0 | EDO, Cl−, ATP, Mg2+ | ||
| 4R2M | 2 | 1 (4R2L Mutation) | ANP, Mg2+ | ||
| Nitrosomonas europaea ATCC 19718 | 3TNJ | 1 | 0 | AMP | [61] |
| 2PFS | 1 | 3 | MSE, Cl− | ||
| Archaeoglobus fulgidus | 3QTB | 2 | 0 | D5M, ACT, MSE | |
| 6HCD | 4 | 0 | ACT, MSE, Cl− | ||
| Escherichia coli K-12 | 5CB0 | 2 | 0 | Z6X | [63] |
| Arabidopsis thaliana | 2GM3 | 6 | 3 | AMP, MSE | [64] |
| Plant Species | Name of USPs | Functions | References |
|---|---|---|---|
| Arabidopsis thaliana | AtUSP | Protein chaperone | [85] |
| RNA chaperone | [90] | ||
| Inherently antifungal activity | [55] | ||
| AT5G35380 | Related to resistance to flooding | [86] | |
| GRUSP | Promote seed germination | [36] | |
| Novel regulatory components of the flowering signal transduction pathway | [40] | ||
| HRU1 | Coordination of oxygen sensing and ROS signaling under hypoxic conditions | [87] | |
| AtUSP17 | Related to the salt tolerance of the plant | [42] | |
| Gossypium arboretum | GUSP1 | Activates downstream genes in response to drought | [49,50] |
| GUSP2 | Directly involved in stress tolerance or as signaling molecules to activate stress adaptation mechanisms | [51] | |
| Malus sieversii | MsUSPA | Reducing transpiration and retaining water by altering the cellular structure of the leaves to improve drought resistance | [46] |
| Increase the activity of antioxidant enzymes, reduce the accumulation of Ros and improve the antioxidant capacity | |||
| Solanum lycoperiscus | SlRd2 | Regulation of SlCipk6-mediated ROS generation | [102] |
| Solanum pennellii | SpUSP | Improving tolerance to oxidative stress | [44] |
| Salicornia brachiata | SbUSP | Participate in tolerance mechanisms or act as molecular switches (signaling molecules) to activate stress adaptation mechanisms | [53] |
| Sulfolobus acidocaldarius | SaUspA | Binds phosphatase and alters phosphatase activity | [103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, D.; Wu, Z.; Bai, Q.; Zhang, Y.; Huang, M.; Huang, Y.; Li, X. Universal Stress Proteins: From Gene to Function. Int. J. Mol. Sci. 2023, 24, 4725. https://doi.org/10.3390/ijms24054725
Luo D, Wu Z, Bai Q, Zhang Y, Huang M, Huang Y, Li X. Universal Stress Proteins: From Gene to Function. International Journal of Molecular Sciences. 2023; 24(5):4725. https://doi.org/10.3390/ijms24054725
Chicago/Turabian StyleLuo, Dan, Zilin Wu, Qian Bai, Yong Zhang, Min Huang, Yajiao Huang, and Xiangyang Li. 2023. "Universal Stress Proteins: From Gene to Function" International Journal of Molecular Sciences 24, no. 5: 4725. https://doi.org/10.3390/ijms24054725
APA StyleLuo, D., Wu, Z., Bai, Q., Zhang, Y., Huang, M., Huang, Y., & Li, X. (2023). Universal Stress Proteins: From Gene to Function. International Journal of Molecular Sciences, 24(5), 4725. https://doi.org/10.3390/ijms24054725