

TRPM7-Mediated Ca2+ Regulates Mussel Settlement through the CaMKKβ-AMPK-SGF1 Pathway


Abstract
:1. Introduction
2. Results and Discussion
2.1. TRPM7 Channel Triggers Larval Settlement through Ca2+ Signal
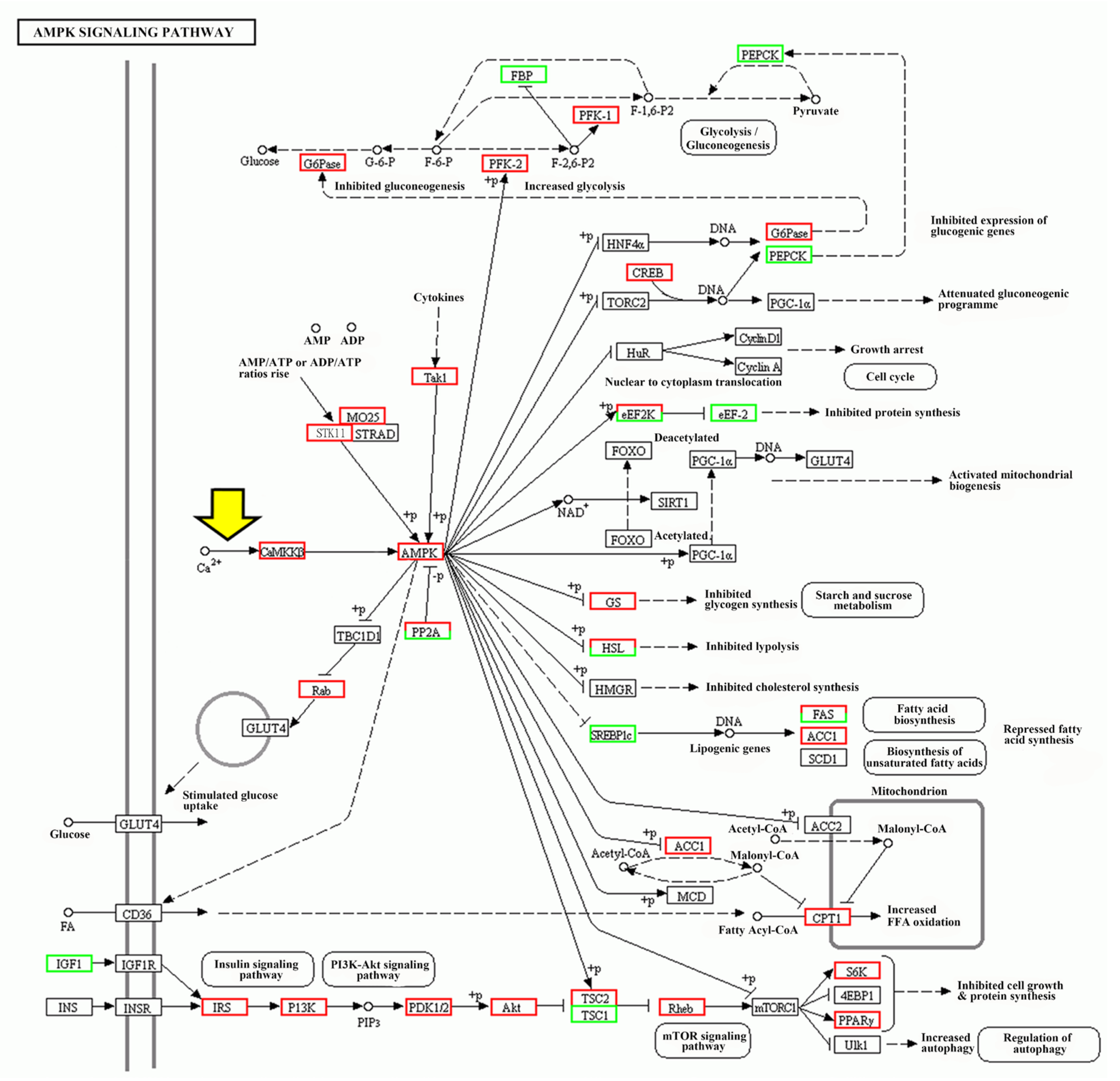
2.2. TRPM7-Mediated Ca2+ Signal Triggers Larval Settlement via CaMKKβ-AMPK Pathway
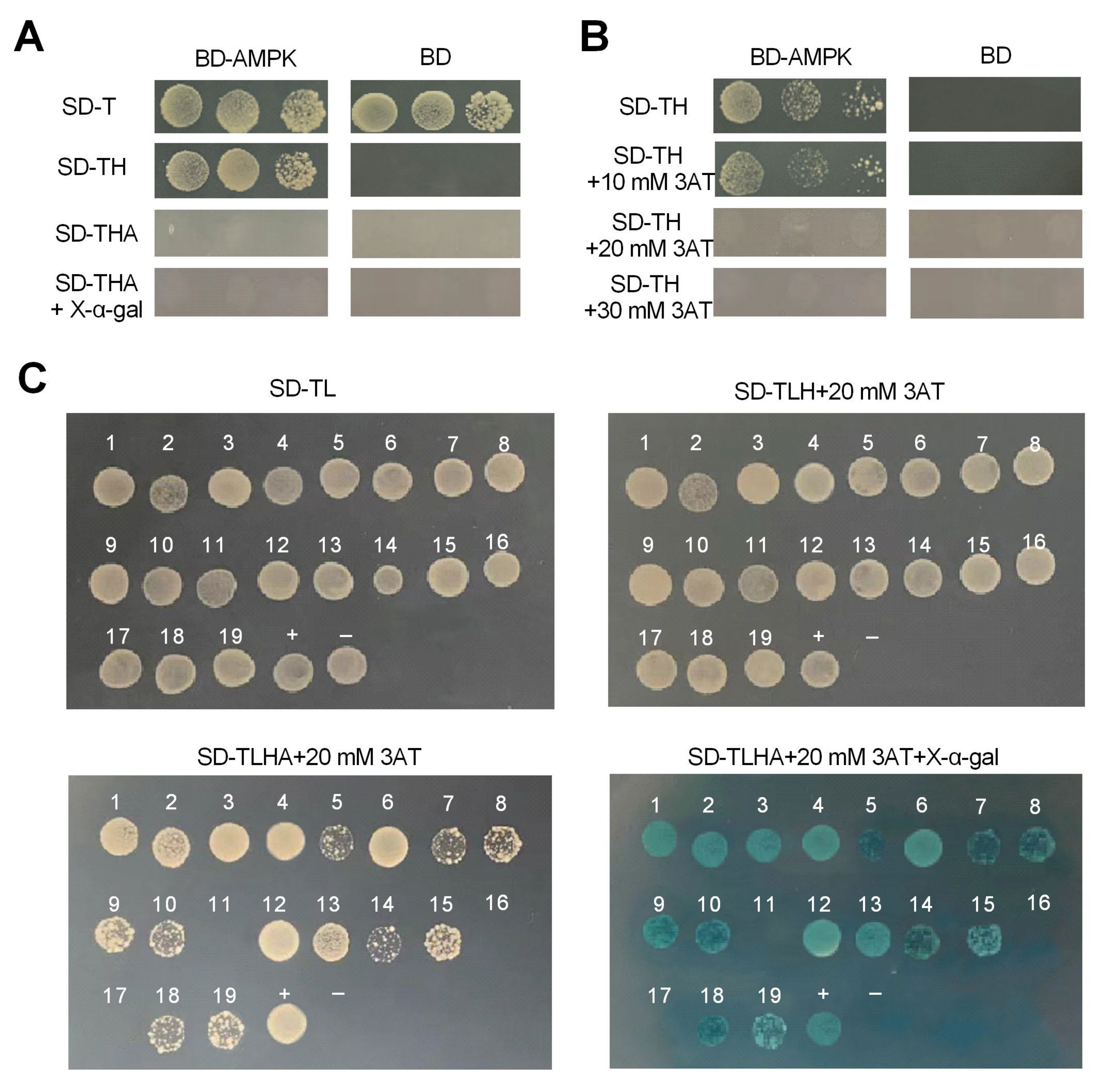
2.3. Yeast-Two-Hybrid Screening of AMPK-Interacting Proteins
2.4. AMPK-Interacting Protein SGF1 Is Responsible for Larval Settlement
2.5. TRPM7-AMPK Pathway Is Involved in Larval Responses to Substrates with Different Stiffness
3. Materials and Methods
3.1. Larval Culture of M. sallei
3.2. Measurement of Ca2+ Flux
3.3. Genes Expression Analysis by qRT-PCR
3.4. Screening and Identification of Proteins Interacted with AMPK by Y2H Assay
3.5. Tissue Localization of SGF1 Analysis by Immunofluorescence
3.6. Knockdown of TRPM7 and SGF1 Expression by siRNA Interference
3.7. Preparation of Substrates with Various Stiffness
3.8. Larval Settlement and Gene Expression in Response to Different Substrates
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, D.; Leys, S.P.; Hinman, V.F.; Woods, R.; Lavin, M.; Degnan, B. Ecological regulation of development: Induction of marine invertebrate metamorphosis. Int. J. Dev. Biol. 2004, 46, 679–686. [Google Scholar]
- Pechenik, J.A.; Wendt, D.E.; Jarrett, J.N. Metamorphosis is not a new beginning: Larval experience influences juvenile performance. Bioscience 1998, 48, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Cowen, R.K.; Sponaugle, S. Larval dispersal and marine population connectivity. Annu. Rev. Mar. Sci. 2009, 1, 443–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marechal, J.-P.; Hellio, C. Challenges for the development of new non-toxic antifouling solutions. Int. J. Mol. Sci. 2009, 10, 4623–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, N.L.; Banta, W.C.; Loeb, G.I. Aquatic biofouling larvae respond to differences in the mechanical properties of the surface on which they settle. Biofouling 2002, 18, 269–273. [Google Scholar] [CrossRef]
- Rosenhahn, A.; Sendra, G.H. Surface sensing and settlement strategies of marine biofouling organisms. Biointerphases 2012, 7, 63. [Google Scholar] [CrossRef] [Green Version]
- Jin, P.; Jan, L.Y.; Jan, Y.-N. Mechanosensitive ion channels: Structural features relevant to mechanotransduction mechanisms. Annu. Rev. Neurosci. 2020, 43, 207–229. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Lee, S.; Ohkawa, K.; Hwang, D.S. Counterplotting the mechanosensing-based gouling mechanism of mussels against fouling. ACS Nano 2021, 15, 18566–18579. [Google Scholar] [CrossRef]
- Amini, S.; Kolle, S.; Petrone, L.; Ahanotu, O.; Sunny, S.; Sutanto, C.N.; Hoon, S.; Cohen, L.; Weaver, J.C.; Aizenberg, J.; et al. Preventing mussel adhesion using lubricant-infused materials. Science 2017, 357, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Abramova, A.; Rosenblad, M.A.; Blomberg, A.; Larsson, T.A. Sensory receptor repertoire in cyprid antennules of the barnacle Balanus improvisus. PLoS ONE 2019, 14, e0216294. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Michalak, A.M. Environmental sciences: Troubled waters on the Great Lakes. Nature 2017, 543, 488–489. [Google Scholar] [CrossRef] [Green Version]
- Karatayev, A.Y.; Burlakova, L.E.; Mastitsky, S.E.; Padilla, D.K.; Mills, E.L. Contrasting rates of spread of two congeners, Dreissena polymorpha and Dreissena rostriformis bugensis, at different spatial scales. J. Shellfish. Res. 2011, 30, 923–931. [Google Scholar] [CrossRef]
- Kennedy, V.S. The invasive dark falsemussel Mytilopsis leucophaeata (Bivalvia: Dreissenidae): A literature review. Aquat. Ecol. 2011, 45, 163–183. [Google Scholar] [CrossRef]
- Wong, Y.T.; Meier, R.; Tan, K.S. High haplotype variability in established Asian populations of the invasive Carib-bean bivalve Mytilopsis sallei (Dreissenidae). Biol. Invasions 2011, 13, 341–348. [Google Scholar] [CrossRef]
- Tan, K.S.; Tay, T. The invasive Caribbean Mytilopsis sallei (Bivalvia: Dreissenidae): A short review. ASEAN J. Sci. Technol. Dev. 2018, 35, 133–139. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, M.; Liang, X.; Li, F.; Shi, Y.; Yang, X.; Jiang, C. Natural variation of an EF-hand Ca2+-binding-protein coding gene confers saline-alkaline tolerance in maize. Nat. Commun. 2020, 11, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Fan, X.; Gao, G.; Beardall, J.; Inaba, K.; Hall-Spencer, J.M.; Xu, D.; Zhang, X.; Han, W.; McMINN, A.; et al. Decreased motility of flagellated microalgae long-term acclimated to CO2-induced acidified waters. Nat. Clim. Chang. 2020, 10, 561–567. [Google Scholar] [CrossRef]
- Feng, K.; Ni, C.; Yu, L.; Zhou, W.; Li, X. Synthesis and antifouling evaluation of indole derivatives. Ecotoxicol. Environ. Saf. 2019, 182, 109423. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Wu, Z.; Chen, L.; Dai, Q.; Hao, H.; Su, P.; Ke, C.; Feng, D. Adenosine triggers larval settlement and metamorphosis in the mussel Mytilopsis sallei through the ADK-AMPK-FoxO pathway. ACS Chem. Biol. 2021, 16, 1390–1400. [Google Scholar] [CrossRef] [PubMed]
- Tamás, P.; Hawley, S.A.; Clarke, R.G.; Mustard, K.J.; Green, K.; Hardie, G.; Cantrell, D.A. Regulation of the energy sensor AMP-activated protein kinase by antigen receptor and Ca2+ in T lymphocytes. J. Exp. Med. 2006, 203, 1665–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Zhang, W.; Zhang, Y.; Tang, X.; Ling, J.; Zhang, Y.; Dong, J. Promoting larval settlement of coral Pocillopora damicornis by calcium. Coral Reefs 2022, 41, 223–235. [Google Scholar] [CrossRef]
- Matsumura, K.; Mori, S.; Fusetani, N. Induction of larval metamorphosis in the ascidian, Halocynthia roretzi by excess potassium ion and by reduced calcium ion. J. Mar. Bio. Assoc. UK 1999, 79, 1143–1144. [Google Scholar] [CrossRef]
- Clare, A.S. Signal transduction in barnacle settlement: Calcium re-visited. Biofouling 1996, 10, 141–159. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Yan, Y.; Li, H. The Effect of chemical cues onlarval settlement of the abalone, Haliotis diversicolor supertexta. J. World Aquac. Soc. 2010, 41, 626–632. [Google Scholar] [CrossRef]
- Chen, Z.-F.; Wang, H.; Qian, P.-Y. Characterization and expression of calmodulin gene during larval settlement and metamorphosis of the polychaete Hydroides elegans. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2012, 162, 113–119. [Google Scholar] [CrossRef]
- Reyes-Bermudez, A.; DeSalvo, M.K.; Voolstra, C.R.; Sunagawa, S.; Szmant, A.M.; Iglesias-Prieto, R.; Medina, M. Gene expression microarray analysis encompassing metamorphosis and the onset of calcification in the scleractinian coral Montastraea faveolata. Mar. Genom. 2009, 2, 149–159. [Google Scholar] [CrossRef]
- Ueda, N.; Richards, G.S.; Degnan, B.M.; Kranz, A.; Adamska, M.; Croll, R.P.; Degnan, S.M. An ancient role for nitric oxide in regulating the animal pelagobenthic life cycle: Evidence from a marine sponge. Sci. Rep. 2016, 6, 37546. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, Y.; Hu, X.; Zhou, Z.; Zhu, Y.; Liang, X.; Yang, J.-L. AMPK promotes larval metamorphosis of Mytilus coruscus. Genes 2022, 13, 2384. [Google Scholar] [CrossRef]
- Jiang, Z.; Ping, S.; Jin, C.; Tu, C.; Zhou, X. Transcriptome analysis provides insights into a molecular mechanism of histamine response in the cyprid larvae of Amphibalanus amphitrite. Mar. Ecol. Prog. Ser. 2022, 681, 1–12. [Google Scholar] [CrossRef]
- Hardie, D.G.; Schaffer, B.E.; Brunet, A. AMPK: An energy-sensing pathway with multiple inputs and outputs. Trends Cell Biol. 2016, 26, 190–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of cellular energy sensing and restoration of metabolic balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-F.; Zhang, H.; Wang, H.; Matsumura, K.; Wong, Y.H.; Ravasi, T.; Qian, P.-Y. Quantitative proteomics study of larval settlement in the Barnacle Balanus amphitrite. PLoS ONE 2014, 9, e88744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandramouli, K.H.; Sun, J.; Mok, F.S.; Liu, L.; Qiu, J.-W.; Ravasi, T.; Qian, P.-Y. Transcriptome and quantitative proteome analysis reveals molecular processes associated with larval metamorphosis in the polychaete Pseudopolydora vexillosa. J. Proteome Res. 2013, 12, 1344–1358. [Google Scholar] [CrossRef] [PubMed]
- Lam, E.W.-F.; Brosens, J.; Gomes, A.R.; Koo, C.Y. Forkhead box proteins: Tuning forks for transcriptional harmony. Nat. Rev. Cancer 2013, 13, 482–495. [Google Scholar] [CrossRef] [PubMed]
- Julien, E.; Bordeaux, M.-C.; Garel, A.; Couble, P. Fork head alternative binding drives stage-specific gene expression in the silk gland of Bombyx mori. Insect Biochem. Mol. Biol. 2002, 32, 377–387. [Google Scholar] [CrossRef]
- Ma, L.; Li, K.; Guo, Y.; Sun, X.; Deng, H.; Li, K.; Feng, Q.; Li, S. Ras-Raf-MAPK signaling promotes nuclear localization of FOXA transcription factor SGF1 via Ser91 phosphorylation. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2018, 1865, 560–571. [Google Scholar] [CrossRef]
- McCartney, M.A. Structure, function and parallel evolution of the bivalve byssus, with insights from proteomes and the zebra mussel genome. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200155. [Google Scholar] [CrossRef]
- Croll, R.P.; Jackson, D.L.; Voronezhskaya, E.E. Catecholamine-containing cells in larval and postlarval bivalve molluscs. Biol. Bull. 1997, 193, 116–124. [Google Scholar] [CrossRef]
- Joyce, A.; Vogeler, S. Molluscan bivalve settlement and metamorphosis: Neuroendocrine inducers and morphogenetic responses. Aquaculture 2018, 487, 64–82. [Google Scholar] [CrossRef]
- He, J.; Qi, J.F.; Feng, D.Q.; Ke, C.H. Embryonic and larval development of the invasive biofouler Mytilopsis sallei (Récluz, 1849) (Bivalvia: Dreissenidae). J. Mollus. Stud. 2016, 82, 23–30. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Protein Name | Accession | No. | Protein Name | Accession |
|---|---|---|---|---|---|
| 1 | Mitochondrial ATP synthase lipid-binding protein-like protein (ATPS) | MZ592789.1 | 9 | Ferritin | JQ959539.1 |
| 2 | E3 ubiquitin-protein ligase TRIM56 (E3UPL) | XM_045315934.1 | 10 | Tubulin beta-1 chain (Tubulin β-1) | XM_003979430.2 |
| 3 | Elongation factor 2 (EF-2) | XM_012821713.1 | 12 | 6-phosphofructo-2-kinase (PFK-2) | XM_011454180.2 |
| 4 | 60S acidic ribosomal protein P2 (RPP2) | XM_005102789.3 | 13 | Adenine nucleotide translocator (ANT) | KX361239.1 |
| 5 | Silk gland factor 1-like (SGF1) | XP_021361791.1 | 14 | Calcium/calmodulin-dependent protein kinase kinase 2 (CaMKKβ) | XR_001204123.1 |
| 6 | Cytochrome P450 4B1 (P450) | XM_016429472.1 | 15 | Serine/threonine kinase 11 (STK11) | XM_012645487.1 |
| 7 | Glycogen synthase (GS) | XM_014826310.1 | 18 | Acetyl-CoA carboxylase (ACACA) | XM_014254957.1 |
| 8 | Uncharacterized protein | _ | 19 | Uncharacterized protein | _ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Wang, P.; Wang, Z.; Feng, D.; Zhang, D. TRPM7-Mediated Ca2+ Regulates Mussel Settlement through the CaMKKβ-AMPK-SGF1 Pathway. Int. J. Mol. Sci. 2023, 24, 5399. https://doi.org/10.3390/ijms24065399
He J, Wang P, Wang Z, Feng D, Zhang D. TRPM7-Mediated Ca2+ Regulates Mussel Settlement through the CaMKKβ-AMPK-SGF1 Pathway. International Journal of Molecular Sciences. 2023; 24(6):5399. https://doi.org/10.3390/ijms24065399
Chicago/Turabian StyleHe, Jian, Peng Wang, Zhixuan Wang, Danqing Feng, and Dun Zhang. 2023. "TRPM7-Mediated Ca2+ Regulates Mussel Settlement through the CaMKKβ-AMPK-SGF1 Pathway" International Journal of Molecular Sciences 24, no. 6: 5399. https://doi.org/10.3390/ijms24065399
APA StyleHe, J., Wang, P., Wang, Z., Feng, D., & Zhang, D. (2023). TRPM7-Mediated Ca2+ Regulates Mussel Settlement through the CaMKKβ-AMPK-SGF1 Pathway. International Journal of Molecular Sciences, 24(6), 5399. https://doi.org/10.3390/ijms24065399