
Fishing the Targets of Bioactive Compounds from Psidium guajava L. Leaves in the Context of Diabetes
 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Identification of the Phenolic Compositions
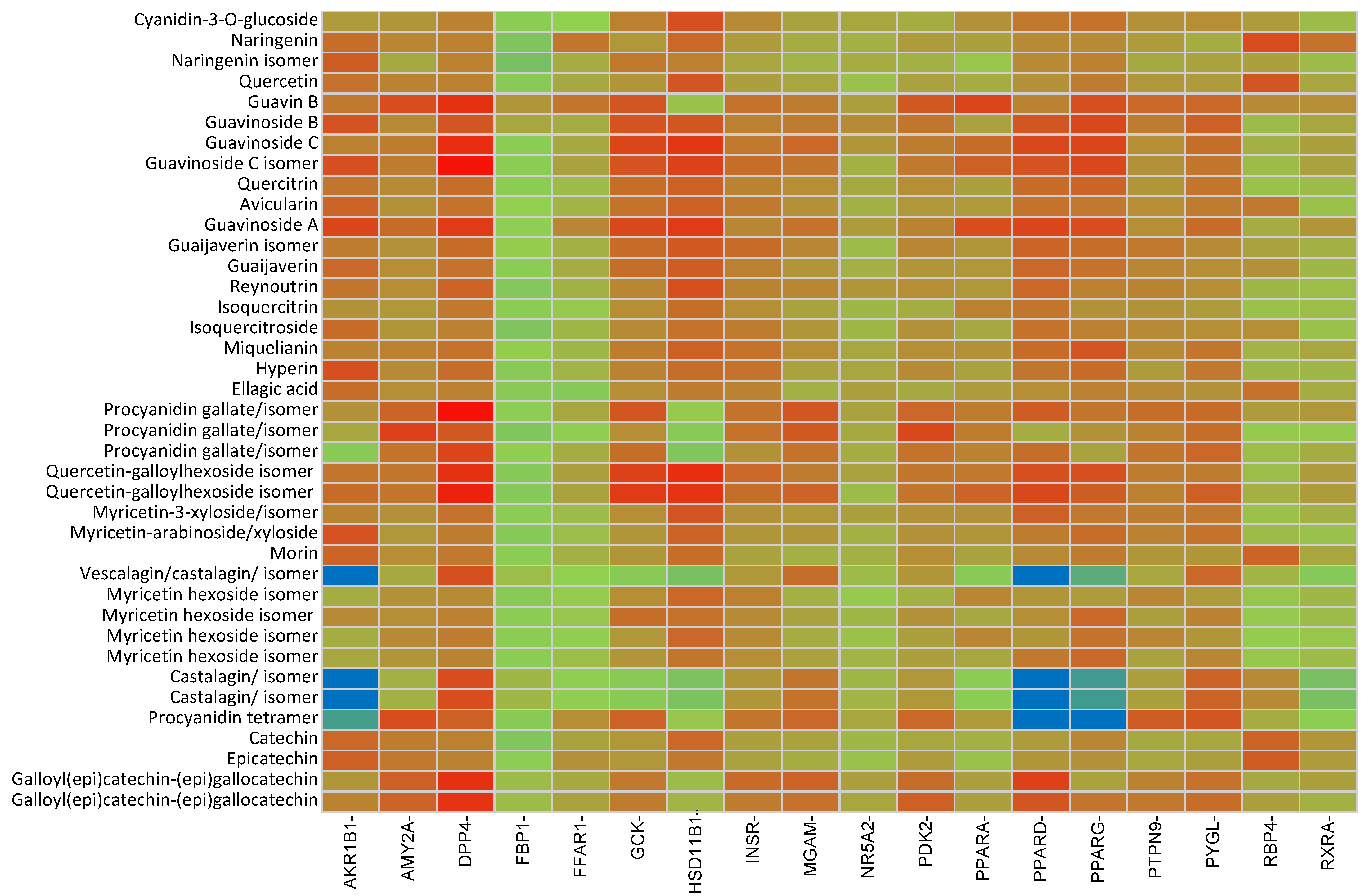
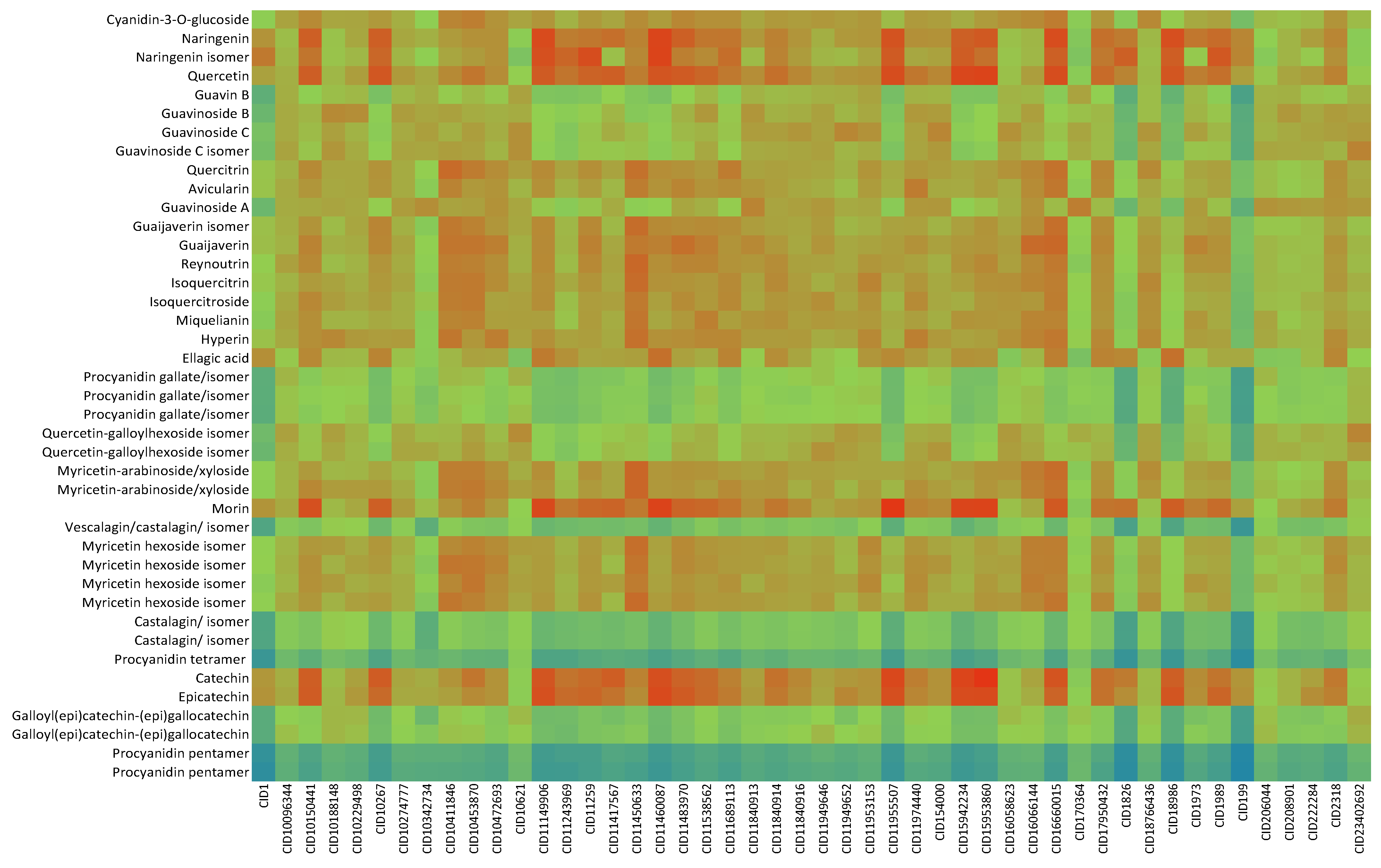
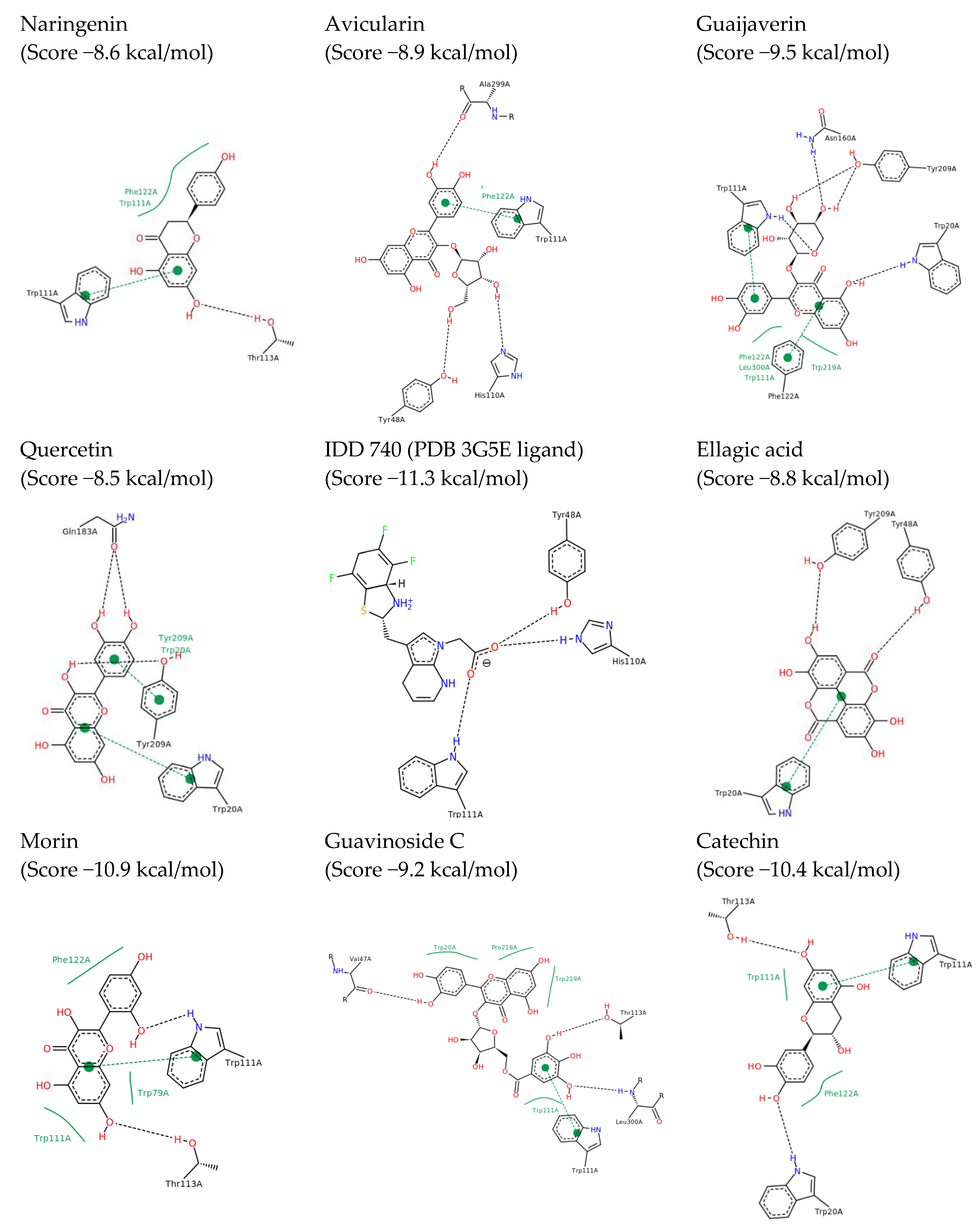
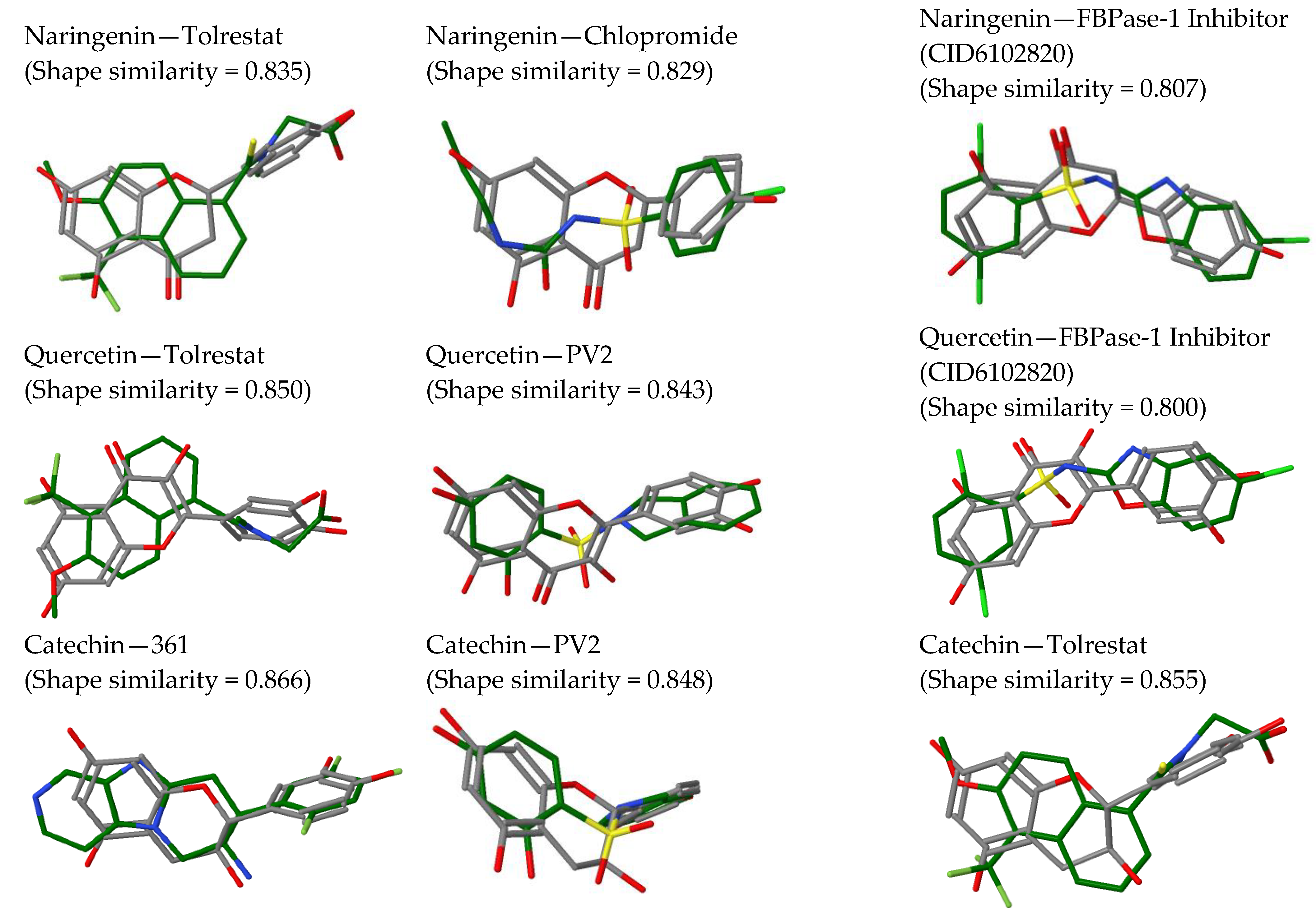
2.2. In Silico Results and Bibliography Searches
3. Discussion
4. Materials and Methods
4.1. Plant Material and Sample Preparation
4.2. HPLC-ESI-QTOF-MS Analyses
4.3. In Silico Approaches
4.4. Bibliography Searches
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035 for the IDF Diabetes Atlas. Diabetes Res. Clin. Pract. 2013, 103, 137–149. [Google Scholar] [CrossRef]
- Soltani, N. Prevention of Diabetes Complications. In Type 1 Diabetes Complications; Wagner, D., Ed.; InTech: Rijeka, Croatia, 2011; pp. 353–366. [Google Scholar]
- Tan, C.; Wang, Q.; Luo, C.; Chen, S.; Li, Q.; Li, P. Yeast α-glucosidase inhibitory phenolic compounds isolated from Gynura medica leaf. Int. J. Mol. Sci. 2013, 14, 2551–2558. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Kaur, N.; Kishore, L.; Gupta, G.K. Management of diabetic complications: A chemical constituents based approach. J. Ethnopharmacol. 2013, 150, 51–70. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, R.M.P.; Mitchell, S.; Solis, R.V. Psidium guajava: A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Díaz-de-Cerio, E.; Gómez-Caravaca, A.M.; Verardo, V.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Determination of guava (Psidium guajava L.) leaf phenolic compounds using HPLC-DAD-QTOF-MA. J. Funct. Foods. 2016, 22, 376–388. [Google Scholar] [CrossRef]
- Díaz-De-Cerio, E.; Verardo, V.; Gómez-Caravaca, A.M.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Exploratory characterization of phenolic compounds with demonstrated anti-diabetic activity in guava leaves at different Oxidation States. Int. J. Mol. Sci. 2016, 17, 699. [Google Scholar] [CrossRef] [Green Version]
- Gambari, R. Predictive analyses of biological effects of natural products: From plant extracts to biomolecular laboratory and computer modeling. Evid. Based Complement. Altern. Med. 2011, 2011, 383290. [Google Scholar] [CrossRef] [Green Version]
- Prathipati, P.; Ngai, L.M.; Manjunatha, U.H.; Bender, A. Fishing the target of antitubercular compounds: In silico target deconvolution model development and validation. J. Proteome Res. 2009, 8, 2788–2798. [Google Scholar] [CrossRef] [PubMed]
- de Rijke, E.; Out, P.; Niessen, W.M.A.; Ariese, F.; Gooijer, C.; Brinkman, U.A.T. Analytical separation and detection methods for flavonoids. J. Chromatogr. A 2006, 1112, 31–63. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.L. HPLC in natural product analysis: The detection issue. Planta Med. 2009, 75, 719–734. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Anand, S.; Arasakumari, M.; Prabu, P.; Amalraj, A.J. Anti-diabetic and aldose reductase inhibitory potential of Psidium guajava by in vitro analysis. Int. J. Pharm. Pharm. Sci. 2016, 8, 271–276. [Google Scholar] [CrossRef]
- Ortiz-Andrade, R.R.; Sánchez-Salgado, J.C.; Navarrete-Vázquez, G.; Webster, S.P.; Binnie, M.; García-Jiménez, S.; León-Rivera, I.; Cigarroa-Vázquez, P.; Villalobos-Molina, R.; Estrada-Soto, S. Antidiabetic and toxicological evaluations of naringenin in normoglycaemic and NIDDM rat models and its implications on extra-pancreatic glucose regulation. Diabetes Obes. Metab. 2008, 10, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Qais, F.A.; Rehman, M.T.; Ismail, M.H.; Alokail, M.S.; Altwaijry, N.; Alafaleq, N.O.; AlAjmi, M.F.; Salem, N.; Alqhatani, R. Mechanistic inhibition of non-enzymatic glycation and aldose reductase activity by naringenin: Binding, enzyme kinetics and molecular docking analysis. Int. J. Biol. Macromol. 2020, 159, 87–97. [Google Scholar] [CrossRef]
- Fan, J.; Johnson, M.H.; Lila, M.A.; Yousef, G.; De Mejia, E.G. Berry and citrus phenolic compounds inhibit dipeptidyl peptidase IV: Implications in diabetes management. Evid. Based Complement. Altern. Med. 2013, 2013, 479505. [Google Scholar] [CrossRef] [Green Version]
- Goldwasser, J.; Cohen, P.Y.; Yang, E.; Balaguer, P.; Yarmush, M.L.; Nahmias, Y. Transcriptional regulation of human and rat hepatic lipid metabolism by the grapefruit flavonoid naringenin: Role of PPARα, PPARγ and LXRα. PLoS ONE 2010, 5, e12399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chethan, S.; Dharmesh, S.M.; Malleshi, N.G. Inhibition of aldose reductase from cataracted eye lenses by finger millet (Eleusine coracana) polyphenols. Bioorg. Med. Chem. 2008, 16, 10085–10090. [Google Scholar] [CrossRef]
- De La Fuente, J.Á.; Manzanaro, S.; Martín, M.J.; De Quesada, T.G.; Reymundo, I.; Luengo, S.M.; Gago, F. Synthesis, activity, and molecular modeling studies of novel human aldose reductase inhibitors based on a marine natural product. J. Med. Chem. 2003, 46, 5208–5221. [Google Scholar] [CrossRef]
- Ueda, H.; Tachibana, Y.; Moriyasu, M.; Kawanishi, K.; Alves, S.M. Aldose reductase inhibitors from the fruits of Caesalpinia ferrea Mart. Phytomedicine 2001, 8, 377–381. [Google Scholar] [CrossRef]
- Torres-Piedra, M.; Ortiz-Andrade, R.; Villalobos-Molina, R.; Singh, N.; Medina-Franco, J.L.; Webster, S.P.; Binnie, M.; Navarrete-Vázquez, G.; Estrada-Soto, S. A comparative study of flavonoid analogues on streptozotocin–nicotinamide induced diabetic rats: Quercetin as a potential antidiabetic agent acting via 11β-Hydroxysteroid dehydrogenase type 1 inhibition. Eur. J. Med. Chem. 2010, 45, 2606–2612. [Google Scholar] [CrossRef]
- Miranda, M.A.; Okamoto, A.K.; Ferreira, C.V.; Silva, T.L.; Granjeiro, J.M.; Aoyama, H. Differential effects of flavonoids on bovine kidney low molecular mass protein tyrosine phosphatase. J. Enzyme Inhib. Med. Chem. 2006, 21, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.; Martinez, K.; Xie, G.; Kennedy, A.; Bumrungpert, A.; Overman, A.; Jia, W.; McIntosh, M.M. Quercetin is equally or more effective than resveratrol in attenuating tumor necrosis factor-α-mediated inflammation and insulin resistance in primary human adipocytes. Am. J. Clin. Nutr. 2010, 92, 1511–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, M.; Nishida, N.; Shimoda, H.; Takada, M.; Kawahara, Y.; Matsuda, H. Polyphenol constituents from Salacia species: Quantitative analysis of mangiferin with alpha-glucosidase and aldose reductase inhibitory activities. Yakugaku Zasshi 2001, 121, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhagat, U.; Endo, S.; Hara, A.; El-Kabbani, O. Inhibition of 3(17)α-hydroxysteroid dehydrogenase (AKR1C21) by aldose reductase inhibitors. Bioorg. Med. Chem. 2008, 16, 3245–3254. [Google Scholar] [CrossRef]
- Kim, H.M.; Lee, D.G.; Lee, S. Plant-derived molecules from Saussurea grandifolia as inhibitors of aldose reductase. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 365–371. [Google Scholar] [CrossRef]
- Jung, H.A.; Islam, M.D.N.; Kwon, Y.S.; Jin, S.E.; Son, Y.K.; Park, J.J.; Sohn, H.S.; Choi, J.S. Extraction and identification of three major aldose reductase inhibitors from Artemisia montana. Food Chem. Toxicol. 2011, 49, 376–384. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Shimada, H.; Nishida, N.; Li, Y.; Toguchida, I.; Yamahara, J.; Matsuda, H. Antidiabetic principles of natural medicines. II. Aldose reductase and α-glucosidase inhibitors from Brazilian natural medicine, the leaves of Myrcia multiflora DC. (Myrtaceae): Structures of myrciacitrins I and II and myrciaphenones A and B. Chem. Pharm. Bull. 1998, 46, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-S.; Bae, J.-Y.; Kim, D.S.; Li, J.; Kim, J.-L.; Lee, Y.-J.; Kang, Y.-H. Dietary Compound Quercitrin Dampens VEGF Induction and PPARγ Activation in Oxidized LDL-Exposed Murine Macrophages: Association with Scavenger Receptor CD36. J. Agric. Food Chem. 2010, 58, 1333–1341. [Google Scholar] [CrossRef]
- Brindis, F.; González-Trujano, M.E.; González-Andrade, M.; Aguirre-Hernández, E.; Villalobos-Molina, R. Aqueous extract of Annona macroprophyllata: A potential α-glucosidase inhibitor. BioMed Res. Int. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Akileshwari, C.; Raghu, G.; Muthenna, P.; Mueller, N.H.; Suryanaryana, P.; Petrash, J.M.; Reddy, G.B. Bioflavonoid ellagic acid inhibits aldose reductase: Implications for prevention of diabetic complications. J. Funct. Foods 2014, 6, 374–383. [Google Scholar] [CrossRef]
- Hundsdörfer, C.; Hemmerling, H.J.; Götz, C.; Totzke, F.; Bednarski, P.; Le Borgne, M.; Jose, J. Indeno[1,2-b]indole derivatives as a novel class of potent human protein kinase CK2 inhibitors. Bioorg. Med. Chem. 2012, 20, 2282–2289. [Google Scholar] [CrossRef] [PubMed]
- Naeem, S.; Hylands, P.; Barlow, D. Construction of an Indonesian herbal constituents database and its use in Random Forest modelling in a search for inhibitors of aldose reductase. Bioorg. Med. Chem. 2012, 20, 1251–1258. [Google Scholar] [CrossRef]
- Sawant, L.; Singh, V.K.; Dethe, S.; Bhaskar, A.; Balachandran, J.; Mundkinajeddu, D.; Agarwal, A. Aldose reductase and protein tyrosine phosphatase 1B inhibitory active compounds from Syzygium cumini seeds. Pharm. Biol. 2015, 53, 1176–1182. [Google Scholar] [CrossRef] [Green Version]
- Kawanishi, K.; Moriyasu, M.; Kuroiwa, E.; Tachibana, Y.; Ayala, F.; Ueda, H. Aldose reductase inhibitors from the leaves of Myrciaria dubia (H. B. & K.) McVaugh. Phytomedicine 2004, 11, 652–656. [Google Scholar]
- Chatzileontiadou, D.S.M.; Skamnaki, V.T.; Kyriakis, E.; Kantsadi, A.L.; Stravodimos, G.A.; Leonidas, D.D. Natural flavonoids as antidiabetic agents. The binding of gallic and ellagic acids to glycogen phosphorylase b. FEBS Lett. 2015, 589, 1787–1794. [Google Scholar]
- Zoechling, A.; Liebner, F.; Jungbauer, A. Red wine: A source of potent ligands for peroxisome proliferator-activated receptor γ. Food Funct. 2011, 2, 28–38. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Chanathong, B. α-Glucosidase inhibitory activity and lipid-lowering mechanisms of Moringa oleifera leaf extract. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 803–808. [Google Scholar]
- Toma, A.; Makonnen, E.; Mekonnen, Y.; Debella, A.; Addisakwattana, S. Intestinal α-glucosidase and some pancreatic enzymes inhibitory effect of hydroalcholic extract of Moringa stenopetala leaves. BMC Complement. Altern. Med. 2014, 14, 180. [Google Scholar] [CrossRef] [Green Version]
- Palanisamy, U.D.; Ling, L.T.; Manaharan, T.; Appleton, D. Rapid isolation of geraniin from Nephelium lappaceum rind waste and its anti-hyperglycemic activity. Food Chem. 2011, 127, 21–27. [Google Scholar] [CrossRef]
- Abbas, G.; Hussain, H.; Hamaed, A.; Supuran, C.T. The management of diabetes mellitus-imperative role of natural products against dipeptidyl peptidase-4, α-glucosidase and sodium-dependent glucose co-transporter 2 (SGLT2). Bioorg. Chem. 2019, 86, 305–315. [Google Scholar] [CrossRef]
- Zhu, Q.; Ge, F.; Dong, Y.; Sun, W.; Wang, Z.; Shan, Y.; Chen, R.; Sun, J.; Ge, R.-S. Comparison of flavonoids and isoflavonoids to inhibit rat and human 11β-hydroxysteroid dehydrogenase 1 and 2. Steroids 2018, 132, 25–32. [Google Scholar] [CrossRef]
- Shen, S.C.; Cheng, F.C.; Wu, N.J. Effect of guava (Psidium guajava Linn.) leaf soluble solids on glucose metabolism in type 2 diabetic rats. Phytother. Res. 2008, 22, 1458–1464. [Google Scholar] [CrossRef]
- Lee, Y.; Lim, Y.; Kwon, O. Selected phytochemicals and culinary plant extracts inhibit fructose uptake in caco-2 cells. Molecules 2015, 20, 17393–17404. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.J.; Yan, W.L.; Yu, S.C.; Gunawan, G.; Lin, C.Y.; Huang, C.Y.; Chang, C.T.; Chen, H.W.; Lii, C.K.; Yu, A.L.; et al. Inactivation of Protein Tyrosine Phosphatase 1B (PTP1B) Activity by the Aqueous Partition of Guava Leaf Extract. J. Pharm. Pharmacol. 2018, 6, 890–906. [Google Scholar]
- Cheng, F.C.; Shen, S.C.; Wu, J.S.B. Effect of guava (Psidium guajava L.) leaf extract on glucose uptake in rat hepatocytes. J. Food Sci. 2009, 74, H132–H138. [Google Scholar] [CrossRef]
- Yoon, S.-Y.; Lee, J.H.; Kwon, S.J.; Kang, H.J.; Chung, S.J. Ginkgolic acid as a dual-targeting inhibitor for protein tyrosine phosphatases relevant to insulin resistance. Bioorg. Chem. 2018, 81, 264–269. [Google Scholar] [CrossRef]
- Qiang, G.; Xue, S.; Yang, J.J.; Du, G.; Pang, X.; Li, X.; Goswami, D.; Griffin, P.R.; Ortlund, E.A.; Chan, C.B. Identification of a small molecular insulin receptor agonist with potent antidiabetes activity. Diabetes 2014, 63, 1394–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.W.; Wang, Y.C.; Lu, H.C.; Chiang, W.D. Optimization of ultrasound-assisted extraction conditions for total phenols with anti-hyperglycemic activity from Psidium guajava leaves. Process. Biochem. 2014, 49, 1601–1605. [Google Scholar] [CrossRef]
- Oghogho, O.O.; Nimenibo-Uadia, R. Phytochemical screening, GC-MS analysis and in vitro inhibition of alpha-amylase and alpha-glucosidase activities by methanol extract of Psidium guajava leaves and fractions. J. Pharmacogn. Phytochem. 2019, 8, 634–640. [Google Scholar]
- Wang, H.; Du, Y.J.; Song, H.C. α-Glucosidase and α-amylase inhibitory activities of guava leaves. Food Chem. 2010, 123, 6–13. [Google Scholar] [CrossRef]
- Banu, M.; Sujatha, K.; Kamala, M.; Kumar, K.V. Hypoglycaemic and hypolipidaemic potentials of isolated fraction of Psidium guajava leaf in alloxan-induced diabetic rats. Int. J. Pharm. Innov. 2012, 2, 16–22. [Google Scholar]
- Tella, T.; Masola, B.; Mukaratirwa, S. The effect of Psidium guajava aqueous leaf extract on liver glycogen enzymes, hormone sensitive lipase and serum lipid profile in diabetic rats. Biomed. Pharmacother. 2019, 109, 2441–2446. [Google Scholar] [CrossRef] [PubMed]
- Vinayagam, R.; Jayachandran, M.; Chung, S.S.M.; Xu, B. Guava leaf inhibits hepatic gluconeogenesis and increases glycogen synthesis via AMPK/ACC signaling pathways in streptozotocin-induced diabetic rats. Biomed. Pharmacother. 2018, 103, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Caravaca, A.M.; Verardo, V.; Toselli, M.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Caboni, M.F. Determination of the major phenolic compounds in pomegranate juices by HPLC-DAD-ESI-MS. J. Agric. Food Chem. 2013, 61, 5328–5337. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Pérez, A.; Muñoz, A.; Peña-García, J.; Den-Haan, H.; Bekas, N.; Katsikoudi, A.; Tzakos, A.G.; Péréz-Sánchez, H. DIA-DB: A Web-Accessible Database for the Prediction of Diabetes Drugs. In Bioinformatics and Biomedical Engineering, Proceedings of the Third International Conference, IWBBIO 2015, Granada, Spain, 15–17 April 2015; Ortuño, F., Rojas, I., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Ge, H.; Wang, Y.; Zhao, W.; Lin, W.; Yan, X.; Xu, J. Scaffold hopping of potential anti-tumor agents by WEGA: A shape-based approach. MedChemComm 2014, 5, 737–741. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A. Autodock vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | rt (min) | m/z Exp | m/z Calc | Molecular Formula | Score | Error (ppm) |
|---|---|---|---|---|---|---|---|
| Negative mode | |||||||
| 1 | Galloyl-Hexahydroxydiphenoyl (HHDP) glucose isomer 1 | 1.93 | 481.06 | 481.34 | C20H18O14 | 96.51 | −2.55 |
| 2 | HHDP glucose isomer 2 | 2.14 | 481.06 | 481.34 | C20H18O14 | 99.09 | −0.19 |
| 3 | HHDP glucose isomer 3 | 2.52 | 481.06 | 481.34 | C20H18O14 | 97.21 | −2.24 |
| 4 | Prodelphinidin B isomer | 3.85 | 609.13 | 609.51 | C30H26O14 | 97.84 | −1.7 |
| 5 | Gallic acid | 4.02 | 169.01 | 169.11 | C7H6O5 | 99.27 | 0.37 |
| 6 | Pedunculagin/Casuariin isomer 1 | 5.87 | 783.07 | 783.53 | C34H24O22 | 98.57 | −1.29 |
| 7 | Prodelphinidin Dimer isomer 1 | 7.27 | 593.13 | 593.51 | C30H26O13 | 96.51 | −2.35 |
| 8 | (epi)-gallocatechin isomer 1 | 7.81 | 305.07 | 305.26 | C15H14O7 | 95.55 | −3.32 |
| 9 | Vescalagin/castalagin isomer | 7.95 | 933.07 | 933.62 | C41H26O26 | 99.19 | −0.79 |
| 10 | Prodelphinidin Dimer isomer 2 | 8.12 | 593.13 | 593.51 | C30H26O13 | 96.51 | −2.35 |
| 11 | Uralenneoside | 9.39 | 285.06 | 285.23 | C12H14O8 | 97.80 | −2.69 |
| 12 | Geraniin isomer 1 | 9.50 | 951.08 | 951.64 | C41H28O27 | 99.56 | −0.20 |
| 13 | Pedunculagin/Casuariin isomer 2 | 9.54 | 783.07 | 783.53 | C34H24O22 | 98.39 | −1.36 |
| 14 | Geraniin isomer 2 | 9.65 | 951.08 | 951.64 | C41H28O27 | 99.56 | −0.20 |
| 15 | Procyanidin B isomer 1 | 10.02 | 577.14 | 577.51 | C30H26O12 | 95.68 | −2.55 |
| 16 | Galloyl(epi)catechin-(epi)gallocatechin | 10.35 | 745.14 | 745.62 | C37H30O17 | 96.90 | −0.62 |
| 17 | Procyanidin B isomer 2 | 10.36 | 577.14 | 577.51 | C30H26O13 | 99.41 | −0.61 |
| 18 | Tellimagrandin I isomer | 10.74 | 785.09 | 785.55 | C34H26O22 | 99.13 | −0.96 |
| 19 | Pterocarinin A isomer 1 | 11.00 | 1067.12 | 1067.75 | C46H36O30 | 99.82 | −0.11 |
| 20 | Pterocarinin A isomer 2 | 11.21 | 1067.12 | 1067.75 | C46H36O30 | 98.39 | −1.26 |
| 21 | Stenophyllanin A | 11.25 | 1207.15 | 1207.89 | C56H40O31 | 98.64 | −1.08 |
| 22 | Procyanidin trimer isomer 1 | 11.25 | 865.20 | 865.77 | C45H38O18 | 97.53 | −1.59 |
| 23 | (epi)-catechin | 11.26 | 289.07 | 289.26 | C15H14O6 | 96.76 | −3.18 |
| 24 | Procyanidin tetramer | 11.34 | 1153.26 | 1153.03 | C60H50O24 | 99.60 | −0.50 |
| 25 | Procyanidin trimer isomer 2 | 11.41 | 865.20 | 865.77 | C45H38O18 | 97.53 | −1.59 |
| 26 | Guavin A | 11.50 | 1223.14 | 1223.89 | C56H40O32 | 99.05 | 0.85 |
| 27 | Casuarinin/Casuarictin isomer | 11.90 | 935.08 | 935.64 | C41H28O26 | 97.67 | −1.43 |
| 28 | Galloyl(epi)catechin-(epi)gallocatechin | 12.10 | 745.14 | 745.62 | C37H30O17 | 96.90 | −0.62 |
| 29 | Procyanidin pentamer | 12.14 | 1441.32 | 1441.27 | C75H62O30 | 95.66 | 1.97 |
| 30 | Galloyl-(epi)catechin trimer isomer 1 | 12.17 | 1017.21 | 1017.87 | C52H42O22 | 99.72 | −0.01 |
| 31 | (epi)-gallocatechin isomer 2 | 12.33 | 305.07 | 305.26 | C15H14O7 | 95.55 | −3.32 |
| 32 | Tellimagrandin I isomer | 12.50 | 785.09 | 785.55 | C34H26O22 | 98.44 | −1.38 |
| 33 | Vescalagin | 12.76 | 933.07 | 933.62 | C41H26O26 | 96.33 | −0.80 |
| 34 | Stenophyllanin A isomer | 12.93 | 1207.15 | 1207.89 | C56H40O31 | 98.37 | 0.89 |
| 35 | Galloyl-(epi)catechin trimer isomer 2 | 12.99 | 1017.21 | 1017.87 | C52H42O22 | 98.17 | −1.35 |
| 36 | Myricetin hexoside isomer 1 | 13.28 | 479.08 | 479.37 | C21H20O13 | 98.36 | −0.92 |
| 37 | Stachyuranin A | 13.41 | 1225.16 | 1225.91 | C56H42O32 | 95.54 | 1.35 |
| 38 | Procyanidin gallate isomer | 13.52 | 729.15 | 729.62 | C37H30O16 | 96.89 | −1.91 |
| 39 | Myricetin hexoside isomer 2 | 13.68 | 479.08 | 479.37 | C21H20O13 | 97.89 | −0.08 |
| 40 | Vescalagin/castalagin isomer | 13.84 | 933.07 | 933.62 | C41H26O26 | 88.32 | −1.57 |
| 41 | Myricetin-arabinoside/xylopyranoside isomer 1 | 13.99 | 449.07 | 449.34 | C20H18O12 | 98.39 | −1.65 |
| 42 | Myricetin-arabinoside/xylopyranoside isomer 2 | 14.21 | 449.07 | 449.34 | C20H18O12 | 98.02 | −1.65 |
| 43 | Procyanidin gallate isomer | 14.56 | 729.64 | 577.51 | C30H26O12 | 98.17 | −1.73 |
| 44 | Myricetin-arabinoside/xylopyranoside isomer 3 | 14.99 | 449.07 | 449.34 | C20H18O12 | 98.66 | −1.65 |
| 45 | Myricetin hexoside isomer 3 | 15.03 | 479.08 | 479.37 | C21H20O13 | 97.08 | −1.92 |
| 46 | Myricetin hexoside isomer 4 | 15.22 | 479.08 | 479.37 | C21H20O13 | 97.08 | −1.92 |
| 47 | Myricetin-arabinoside/xylopyranoside Isomer 4 | 15.60 | 449.07 | 449.34 | C20H18O12 | 98.39 | −1.65 |
| 48 | Quercetin-galloylhexoside isomer | 15.63 | 615.10 | 615.47 | C28H24O16 | 99.16 | −0.98 |
| 49 | Ellagic acid deoxyhexoside | 15.84 | 447.06 | 447.33 | C20H16O12 | 91.25 | −3.19 |
| 50 | Quercetin-galloylhexoside isomer | 16.04 | 615.10 | 615.47 | C28H24O16 | 99.16 | −0.98 |
| 51 | Myricetin-arabinoside/xylopyranoside isomer 5 | 16.19 | 449.07 | 449.34 | C20H18O12 | 98.39 | −1.65 |
| 52 | Morin | 16.28 | 301.04 | 301.23 | C15H10O7 | 97.46 | −2.50 |
| 53 | Myricetin-arabinoside/xylopyranoside isomer 6 | 16.46 | 449.07 | 449.34 | C20H18O12 | 98.39 | −1.65 |
| 54 | Ellagic acid | 16.51 | 301.00 | 301.19 | C14H6O8 | 98.88 | −1.71 |
| 55 | Hyperin | 16.62 | 463.09 | 463.37 | C21H20O12 | 96.41 | −2.65 |
| 56 | Quercetin glucoronide | 16.72 | 477.07 | 477.35 | C21H18O13 | 98.10 | −1.83 |
| 57 | Isoquercitrin | 16.95 | 463.09 | 463.37 | C21H20O12 | 97.04 | −2.33 |
| 58 | Procyanidin gallate isomer | 17.04 | 729.15 | 729.62 | C37H30O16 | 96.89 | −1.91 |
| 59 | Reynoutrin | 17.50 | 433.08 | 433.34 | C20H18O11 | 95.94 | −2.90 |
| 60 | Guajaverin | 17.80 | 433.08 | 433.34 | C20H18O11 | 97.99 | −1.91 |
| 61 | Guavinoside A isomer 1 | 17.96 | 543.12 | 544.46 | C26H24O13 | 98.10 | −1.77 |
| 62 | Avicularin | 18.21 | 433.08 | 433.34 | C20H18O11 | 96.70 | −2.20 |
| 63 | Quercitrin | 19.19 | 447.10 | 447.37 | C21H20O11 | 95.23 | −3.02 |
| 64 | Myrciaphenone B | 19.21 | 481.10 | 481.38 | C21H22O13 | 97.20 | −2.23 |
| 65 | Guavinoside C | 19.77 | 585.09 | 585.45 | C27H22O15 | 97.19 | −1.92 |
| 66 | Guavinoside B isomer 1 | 20.77 | 571.15 | 571.51 | C28H28O13 | 97.26 | −2.05 |
| 67 | Guavinoside A isomer 2 | 20.70 | 543.12 | 543.45 | C26H24O13 | 98.10 | −1.77 |
| 68 | Guavinoside B isomer 2 | 21.67 | 571.15 | 571.51 | C28H28O13 | 97.26 | −2.05 |
| 69 | 2,6-dihydroxy-3-methyl-4-O-(6″-O-galloyl-β-D-glucopyranosyl)-benzophenone | 21.97 | 557.13 | 557.48 | C27H26O13 | 96.93 | −2.12 |
| 70 | Guavin B | 22.24 | 693.11 | 693.54 | C33H26O17 | 97.82 | −1.67 |
| 71 | Quercetin | 22.31 | 301.04 | 301.23 | C15H10O7 | 98.90 | −1.34 |
| 72 | Naringenin isomer | 26.74 | 271.06 | 271.25 | C15H12O5 | 96.09 | −3.67 |
| Positive mode | |||||||
| 73 | Cyanidin-3-O-glucoside | 3.66 | 449.11 | 449.39 | C21H21O11 | 96.97 | −2.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-de-Cerio, E.; Girón, F.; Pérez-Garrido, A.; Pereira, A.S.P.; Gabaldón-Hernández, J.A.; Verardo, V.; Segura Carretero, A.; Pérez-Sánchez, H. Fishing the Targets of Bioactive Compounds from Psidium guajava L. Leaves in the Context of Diabetes. Int. J. Mol. Sci. 2023, 24, 5761. https://doi.org/10.3390/ijms24065761
Díaz-de-Cerio E, Girón F, Pérez-Garrido A, Pereira ASP, Gabaldón-Hernández JA, Verardo V, Segura Carretero A, Pérez-Sánchez H. Fishing the Targets of Bioactive Compounds from Psidium guajava L. Leaves in the Context of Diabetes. International Journal of Molecular Sciences. 2023; 24(6):5761. https://doi.org/10.3390/ijms24065761
Chicago/Turabian StyleDíaz-de-Cerio, Elixabet, Francisco Girón, Alfonso Pérez-Garrido, Andreia S. P. Pereira, José Antonio Gabaldón-Hernández, Vito Verardo, Antonio Segura Carretero, and Horacio Pérez-Sánchez. 2023. "Fishing the Targets of Bioactive Compounds from Psidium guajava L. Leaves in the Context of Diabetes" International Journal of Molecular Sciences 24, no. 6: 5761. https://doi.org/10.3390/ijms24065761
APA StyleDíaz-de-Cerio, E., Girón, F., Pérez-Garrido, A., Pereira, A. S. P., Gabaldón-Hernández, J. A., Verardo, V., Segura Carretero, A., & Pérez-Sánchez, H. (2023). Fishing the Targets of Bioactive Compounds from Psidium guajava L. Leaves in the Context of Diabetes. International Journal of Molecular Sciences, 24(6), 5761. https://doi.org/10.3390/ijms24065761