

Treatment with the Glycosphingolipid Modulator THI Rescues Myelin Integrity in the Striatum of R6/2 HD Mice

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. THI Rescues Axonal Myelination Loss within Striatum of R6/2 Transgenic Mice
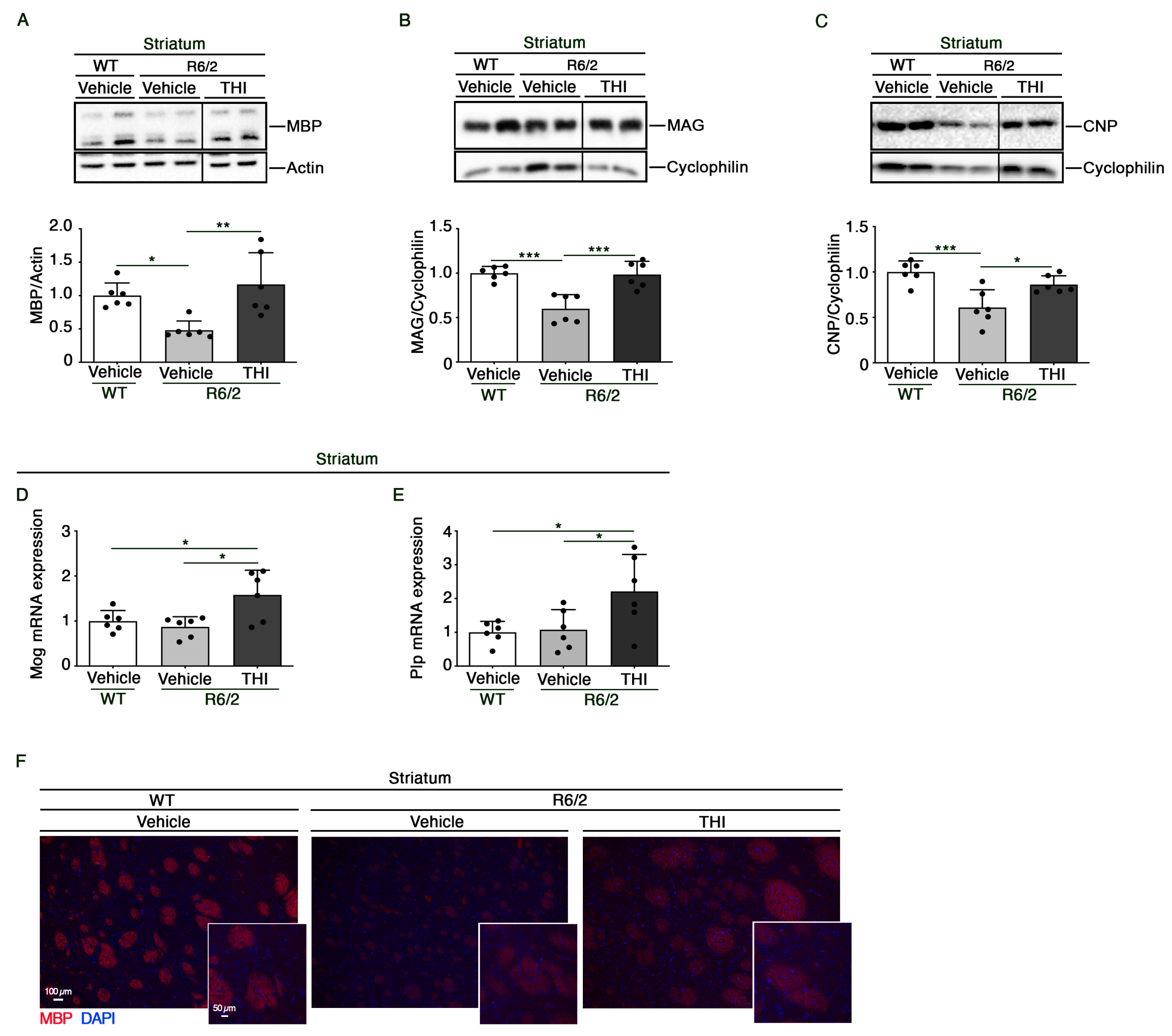
2.2. THI Treatment Preserves the Expression of Myelin-Related Proteins and Genes in the Striatum of R6/2 Mice
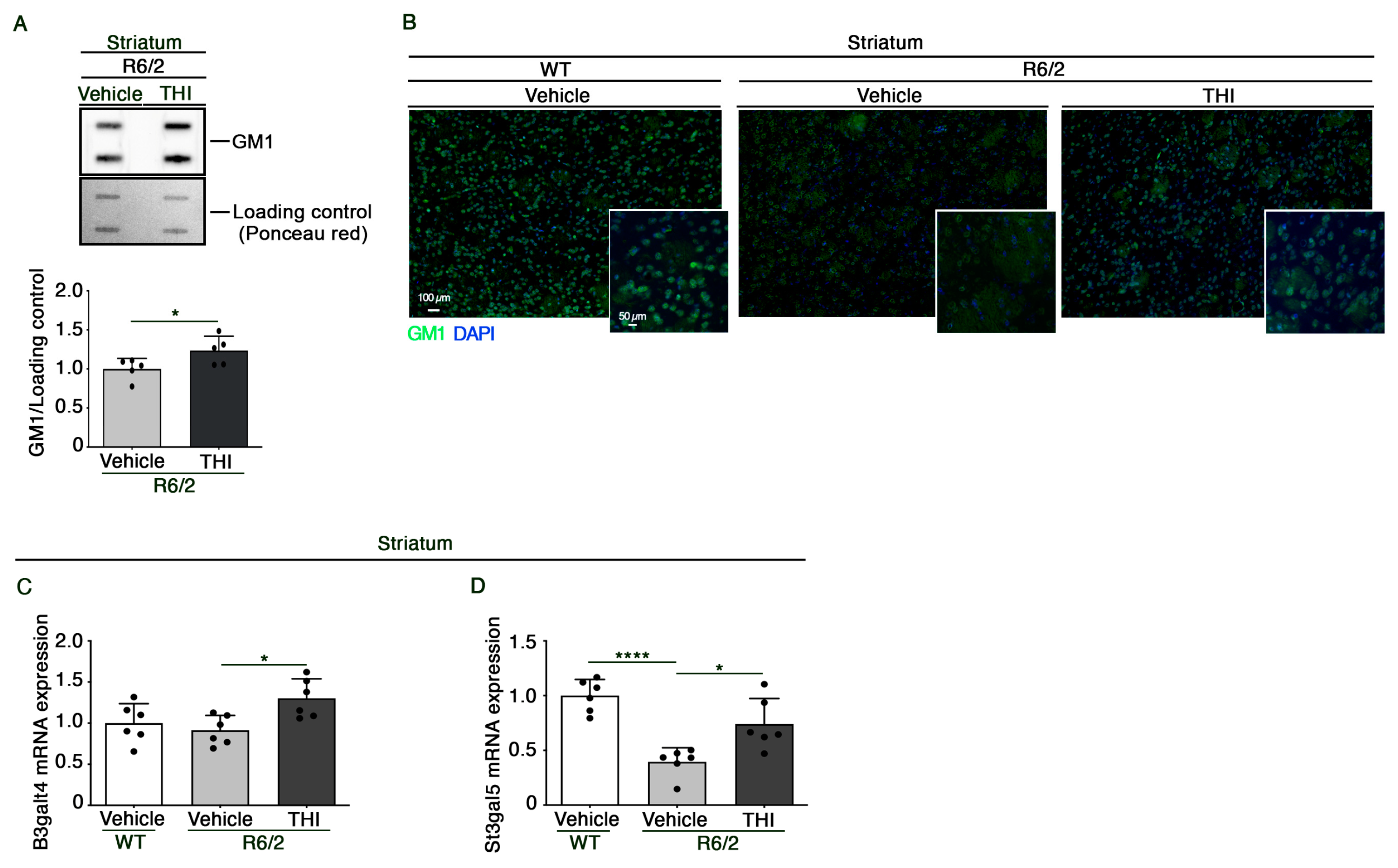
2.3. Administration of THI Modulates Metabolism of Gangliosides in the Striatum of R6/2 Mice
3. Discussion
4. Materials and Methods
4.1. Animal Model
4.2. In Vivo Drug Administration
4.3. Transmission Electron Microscopy (TEM)
4.4. Brain Lysate Preparation and Immunoblottings
4.5. Semiquantitative Analysis of Ganglioside GM1
4.6. Quantitative Real-Time PCR
4.7. Histochemical Analyses
4.8. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tabrizi, S.J.; Flower, M.D.; Ross, C.A.; Wild, E.J. Huntington Disease: New Insights into Molecular Pathogenesis and Therapeutic Opportunities. Nat. Rev. Neurol. 2020, 16, 529–546. [Google Scholar] [CrossRef]
- Saudou, F.; Humbert, S. The Biology of Huntingtin. Neuron 2016, 89, 910–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Agrawal, N. Metabolism in Huntington’s Disease: A Major Contributor to Pathology. Metab. Brain Dis. 2022, 37, 1757–1771. [Google Scholar] [CrossRef] [PubMed]
- De La Monte, S.M.; Vonsattel, J.P.; Richardson, E.P. Morphometric Demonstration of Atrophic Changes in the Cerebral Cortex, White Matter, and Neostriatum in Huntington’s Disease. J. Neuropathol. Exp. Neurol. 1988, 47, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Wilton, D.K.; Stevens, B. The Contribution of Glial Cells to Huntington’s Disease Pathogenesis. Neurobiol. Dis. 2020, 143, 104963. [Google Scholar] [CrossRef]
- Sun, Y.; Tong, H.; Yang, T.; Liu, L.; Li, X.-J.; Li, S. Insights into White Matter Defect in Huntington’s Disease. Cells 2022, 11, 3381. [Google Scholar] [CrossRef]
- Huang, B.; Wei, W.J.; Wang, G.; Gaertig, M.A.; Feng, Y.; Wang, W.; Li, X.J.; Li, S. Mutant Huntingtin Downregulates Myelin Regulatory Factor-Mediated Myelin Gene Expression and Affects Mature Oligodendrocytes. Neuron 2015, 85, 1212–1226. [Google Scholar] [CrossRef] [Green Version]
- Valenza, M.; Rigamonti, D.; Goffredo, D.; Zuccato, C.; Fenu, S.; Jamot, L.; Strand, A.; Tarditi, A.; Woodman, B.; Racchi, M.; et al. Dysfunction of the Cholesterol Biosynthetic Pathway in Huntington’s Disease. J. Neurosci. 2005, 25, 9932–9939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maglione, V.; Marchi, P.; Di Pardo, A.; Lingrell, S.; Horkey, M.; Tidmarsh, E.; Sipione, S. Impaired Ganglioside Metabolism in Huntington’s Disease and Neuroprotective Role of GM1. J. Neurosci. 2010, 30, 4072–4080. [Google Scholar] [CrossRef] [Green Version]
- Di Pardo, A.D.; Amico, E.; Maglione, V. Impaired Levels of Gangliosides in the Corpus Callosum of Huntington Disease Animal Models. Front. Neurosci. 2016, 10, 457. [Google Scholar] [CrossRef] [Green Version]
- Pardo, A.D.; Basit, A.; Armirotti, A.; Amico, E.; Castaldo, S.; Pepe, G.; Marracino, F.; Buttari, F.; Digilio, A.F.; Maglione, V. De Novo Synthesis of Sphingolipids Is Defective in Experimental Models of Huntington’s Disease. Front. Neurosci. 2017, 11, 698. [Google Scholar] [CrossRef] [PubMed]
- Di Pardo, A.; Amico, E.; Basit, A.; Armirotti, A.; Joshi, P.; Neely, D.M.; Vuono, R.; Castaldo, S.; Digilio, A.F.; Scalabrì, F.; et al. Defective Sphingosine-1-Phosphate Metabolism Is a Druggable Target in Huntington’s Disease. Sci. Rep. 2017, 7, 5280. [Google Scholar] [CrossRef] [PubMed]
- Di Pardo, A.; Maglione, V.; Alpaugh, M.; Horkey, M.; Atwal, R.S.; Sassone, J.; Ciammola, A.; Steffan, J.S.; Fouad, K.; Truant, R.; et al. Ganglioside GM1 Induces Phosphorylation of Mutant Huntingtin and Restores Normal Motor Behavior in Huntington Disease Mice. Proc. Natl. Acad. Sci. USA 2012, 109, 3528–3533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alpaugh, M.; Galleguillos, D.; Forero, J.; Morales, L.C.; Lackey, S.W.; Kar, P.; Di Pardo, A.; Holt, A.; Kerr, B.J.; Todd, K.G.; et al. Disease-Modifying Effects of Ganglioside GM1 in Huntington’s Disease Models. EMBO Mol. Med. 2017, 9, 1537–1557. [Google Scholar] [CrossRef]
- Di Pardo, A.; Castaldo, S.; Amico, E.; Pepe, G.; Marracino, F.; Capocci, L.; Giovannelli, A.; Madonna, M.; van Bergeijk, J.; Buttari, F.; et al. Stimulation of S1PR <inf>5</Inf> with A-971432, a Selective Agonist, Preserves Blood-Brain Barrier Integrity and Exerts Therapeutic Effect in an Animal Model of Huntington’s Disease. Hum. Mol. Genet. 2018, 27, 2490–2501. [Google Scholar] [CrossRef] [Green Version]
- Di Pardo, A.; Maglione, V. The S1P Axis: New Exciting Route for Treating Huntington’s Disease. Trends Pharmacol. Sci. 2018, 39, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Di Pardo, A.; Pepe, G.; Castaldo, S.; Marracino, F.; Capocci, L.; Amico, E.; Madonna, M.; Giova, S.; Jeong, S.K.; Park, B.-M.; et al. Stimulation of Sphingosine Kinase 1 (SPHK1) Is Beneficial in a Huntington’s Disease Pre-Clinical Model. Front. Mol. Neurosci. 2019, 12, 100. [Google Scholar] [CrossRef] [PubMed]
- Pepe, G.; Capocci, L.; Marracino, F.; Realini, N.; Lenzi, P.; Martinello, K.; Bovier, T.F.; Bichell, T.J.; Scarselli, P.; Di Cicco, C.; et al. Treatment with THI, an Inhibitor of Sphingosine-1-Phosphate Lyase, Modulates Glycosphingolipid Metabolism and Results Therapeutically Effective in Experimental Models of Huntington’s Disease. Mol. Ther. 2023, 31, 282–299. [Google Scholar] [CrossRef]
- Giussani, P.; Prinetti, A.; Tringali, C. The Role of Sphingolipids in Myelination and Myelin Stability and Their Involvement in Childhood and Adult Demyelinating Disorders. J. Neurochem. 2021, 156, 403–414. [Google Scholar] [CrossRef]
- Pan, B.; Fromholt, S.E.; Hess, E.J.; Crawford, T.O.; Griffin, J.W.; Sheikh, K.A.; Schnaar, R.L. Myelin-Associated Glycoprotein and Complementary Axonal Ligands, Gangliosides, Mediate Axon Stability in the CNS and PNS: Neuropathology and Behavioral Deficits in Single- and Double-Null Mice. Exp. Neurol. 2005, 195, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Schnaar, R.L. Brain Gangliosides in Axon-Myelin Stability and Axon Regeneration. FEBS Lett. 2010, 584, 1741–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, O.R.; Joshi, S.H.; Squitieri, F.; Sanchez-Castaneda, C.; Narr, K.; Shattuck, D.W.; Caltagirone, C.; Sabatini, U.; Di Paola, M. Major Superficial White Matter Abnormalities in Huntington’s Disease. Front. Neurosci. 2016, 10, 197. [Google Scholar] [CrossRef] [Green Version]
- Gatto, R.G.; Chu, Y.; Ye, A.Q.; Price, S.D.; Tavassoli, E.; Buenaventura, A.; Brady, S.T.; Magin, R.L.; Kordower, J.H.; Morfini, G.A. Analysis of YFP(J16)-R6/2 Reporter Mice and Postmortem Brains Reveals Early Pathology and Increased Vulnerability of Callosal Axons in Huntington’s Disease. Hum. Mol. Genet. 2015, 24, 5285–5298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatto, R.G.; Ye, A.Q.; Colon-Perez, L.; Mareci, T.H.; Lysakowski, A.; Price, S.D.; Brady, S.T.; Karaman, M.; Morfini, G.; Magin, R.L. Detection of Axonal Degeneration in a Mouse Model of Huntington’s Disease: Comparison between Diffusion Tensor Imaging and Anomalous Diffusion Metrics. Magn. Reson. Mater. Phys. Biol. Med. 2019, 32, 461–471. [Google Scholar] [CrossRef]
- Di Pardo, A.; Amico, E.; Favellato, M.; Castrataro, R.; Fucile, S.; Squitieri, F.; Maglione, V. FTY720 (Fingolimod) Is a Neuroprotective and Disease-Modifying Agent in Cellular and Mouse Models of Huntington Disease. Hum. Mol. Genet. 2014, 23, 2251–2265. [Google Scholar] [CrossRef] [Green Version]
- Lim, R.G.; Al-Dalahmah, O.; Wu, J.; Gold, M.P.; Reidling, J.C.; Tang, G.; Adam, M.; Dansu, D.K.; Park, H.J.; Casaccia, P.; et al. Huntington Disease Oligodendrocyte Maturation Deficits Revealed by Single-Nucleus RNAseq Are Rescued by Thiamine-Biotin Supplementation. Nat. Commun. 2022, 13, 7791. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Peng, Q.; Hou, Z.; Jiang, M.; Wang, X.; Langseth, A.J.; Tao, M.; Barker, P.B.; Mori, S.; Bergles, D.E.; et al. Early White Matter Abnormalities, Progressive Brain Pathology and Motor Deficits in a Novel Knock-in Mouse Model of Huntington’s Disease. Hum. Mol. Genet. 2015, 24, 2508–2527. [Google Scholar] [CrossRef]
- Yi Teo, R.T.; Hong, X.; Yu-Taeger, L.; Huang, Y.; Tan, L.J.; Xie, Y.; To, X.V.; Guo, L.; Rajendran, R.; Novati, A.; et al. Structural and Molecular Myelination Deficits Occur Prior to Neuronal Loss in the YAC128 and BACHD Models of Huntington Disease. Hum. Mol. Genet. 2016, 25, 2621–2632. [Google Scholar] [CrossRef]
- Casella, C.; Lipp, I.; Rosser, A.; Jones, D.K.; Metzler-Baddeley, C. A Critical Review of White Matter Changes in Huntington’s Disease. Mov. Disord. 2020, 35, 1302–1311. [Google Scholar] [CrossRef]
- Harry, G.J.; Toews, A.D. Myelination, dysmyelination, and demyelination. Handb. Dev. Neurotoxicol. 1998, 87–115. [Google Scholar] [CrossRef]
- Osipovitch, M.; Asenjo Martinez, A.; Mariani, J.N.; Cornwell, A.; Dhaliwal, S.; Zou, L.; Chandler-Militello, D.; Wang, S.; Li, X.; Benraiss, S.J.; et al. Human ESC-Derived Chimeric Mouse Models of Huntington’s Disease Reveal Cell-Intrinsic Defects in Glial Progenitor Cell Differentiation. Cell Stem Cell 2019, 24, 107–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benraiss, A.; Wang, S.; Herrlinger, S.; Li, X.; Chandler-Militello, D.; Mauceri, J.; Burm, H.B.; Toner, M.; Osipovitch, M.; Jim Xu, Q.; et al. Human Glia Can Both Induce and Rescue Aspects of Disease Phenotype in Huntington Disease. Nat. Commun. 2016, 7, 11758. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Cho, E.S.; Sha, Q.; Peng, J.; Oksov, Y.; Kam, S.Y.; Ho, M.; Walker, R.H.; Lee, S. Giant Axon Formation in Mice Lacking Kell, XK, or Kell and XK: Animal Models of McLeod Neuroacanthocytosis Syndrome. Am. J. Pathol. 2014, 184, 800–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganay, T.; Boizot, A.; Burrer, R.; Chauvin, J.P.; Bomont, P. Sensory-Motor Deficits and Neurofilament Disorganization in Gigaxonin-Null Mice. Mol. Neurodegener. 2011, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Reading, S.A.J.; Yassa, M.A.; Bakker, A.; Dziorny, A.C.; Gourley, L.M.; Yallapragada, V.; Rosenblatt, A.; Margolis, R.L.; Aylward, E.H.; Brandt, J.; et al. Regional White Matter Change in Pre-Symptomatic Huntington’s Disease: A Diffusion Tensor Imaging Study. Psychiatry Res. 2005, 140, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, M.; Phillips, O.R.; Sanchez-Castaneda, C.; Di Pardo, A.; Maglione, V.; Caltagirone, C.; Sabatini, U.; Squitieri, F. MRI Measures of Corpus Callosum Iron and Myelin in Early Huntington’s Disease. Hum. Brain Mapp. 2014, 35, 3143–3151. [Google Scholar] [CrossRef]
- Sipione, S.; Monyror, J.; Galleguillos, D.; Steinberg, N.; Kadam, V. Gangliosides in the Brain: Physiology, Pathophysiology and Therapeutic Applications. Front. Neurosci. 2020, 14, 572965. [Google Scholar] [CrossRef] [PubMed]
- Desplats, P.A.; Denny, C.A.; Kass, K.E.; Gilmartin, T.; Head, S.R.; Sutcliffe, J.G.; Seyfried, T.N.; Thomas, E.A. Glycolipid and Ganglioside Metabolism Imbalances in Huntington’s Disease. Neurobiol. Dis. 2007, 27, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Miguez, A.; Barriga, G.G.D.; Brito, V.; Straccia, M.; Giralt, A.; Ginés, S.; Canals, J.M.; Alberch, J. Fingolimod (FTY720) Enhances Hippocampal Synaptic Plasticity and Memory in Huntington’s Disease by Preventing P75NTR up-Regulation and Astrocyte-Mediated Inflammation. Hum. Mol. Genet. 2015, 24, 4958–4970. [Google Scholar] [CrossRef] [Green Version]
- Phillips, G.R.; Saville, J.T.; Hancock, S.E.; Brown, S.H.J.; Jenner, A.M.; Mclean, C.; Fuller, M.; Newell, K.A.; Mitchell, T.W. The Long and the Short of Huntington’s Disease: How the Sphingolipid Profile Is Shifted in the Caudate of Advanced Clinical Cases. Brain Commun. 2021, 4, fcab303. [Google Scholar] [CrossRef]
- Hunter, M.; Demarais, N.J.; Faull, R.L.M.; Grey, A.C.; Curtis, M.A. An Imaging Mass Spectrometry Atlas of Lipids in the Human Neurologically Normal and Huntington’s Disease Caudate Nucleus. J. Neurochem. 2021, 157, 2158–2172. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pepe, G.; Lenzi, P.; Capocci, L.; Marracino, F.; Pizzati, L.; Scarselli, P.; Di Pardo, A.; Fornai, F.; Maglione, V. Treatment with the Glycosphingolipid Modulator THI Rescues Myelin Integrity in the Striatum of R6/2 HD Mice. Int. J. Mol. Sci. 2023, 24, 5956. https://doi.org/10.3390/ijms24065956
Pepe G, Lenzi P, Capocci L, Marracino F, Pizzati L, Scarselli P, Di Pardo A, Fornai F, Maglione V. Treatment with the Glycosphingolipid Modulator THI Rescues Myelin Integrity in the Striatum of R6/2 HD Mice. International Journal of Molecular Sciences. 2023; 24(6):5956. https://doi.org/10.3390/ijms24065956
Chicago/Turabian StylePepe, Giuseppe, Paola Lenzi, Luca Capocci, Federico Marracino, Ludovica Pizzati, Pamela Scarselli, Alba Di Pardo, Francesco Fornai, and Vittorio Maglione. 2023. "Treatment with the Glycosphingolipid Modulator THI Rescues Myelin Integrity in the Striatum of R6/2 HD Mice" International Journal of Molecular Sciences 24, no. 6: 5956. https://doi.org/10.3390/ijms24065956
APA StylePepe, G., Lenzi, P., Capocci, L., Marracino, F., Pizzati, L., Scarselli, P., Di Pardo, A., Fornai, F., & Maglione, V. (2023). Treatment with the Glycosphingolipid Modulator THI Rescues Myelin Integrity in the Striatum of R6/2 HD Mice. International Journal of Molecular Sciences, 24(6), 5956. https://doi.org/10.3390/ijms24065956