
Characterization and Clinical Relevance of Endometrial CAFs: Correlation between Post-Surgery Event and Resistance to Drugs

 ,
,
Abstract
:1. Introduction
Statement of Translational Relevance
2. Results
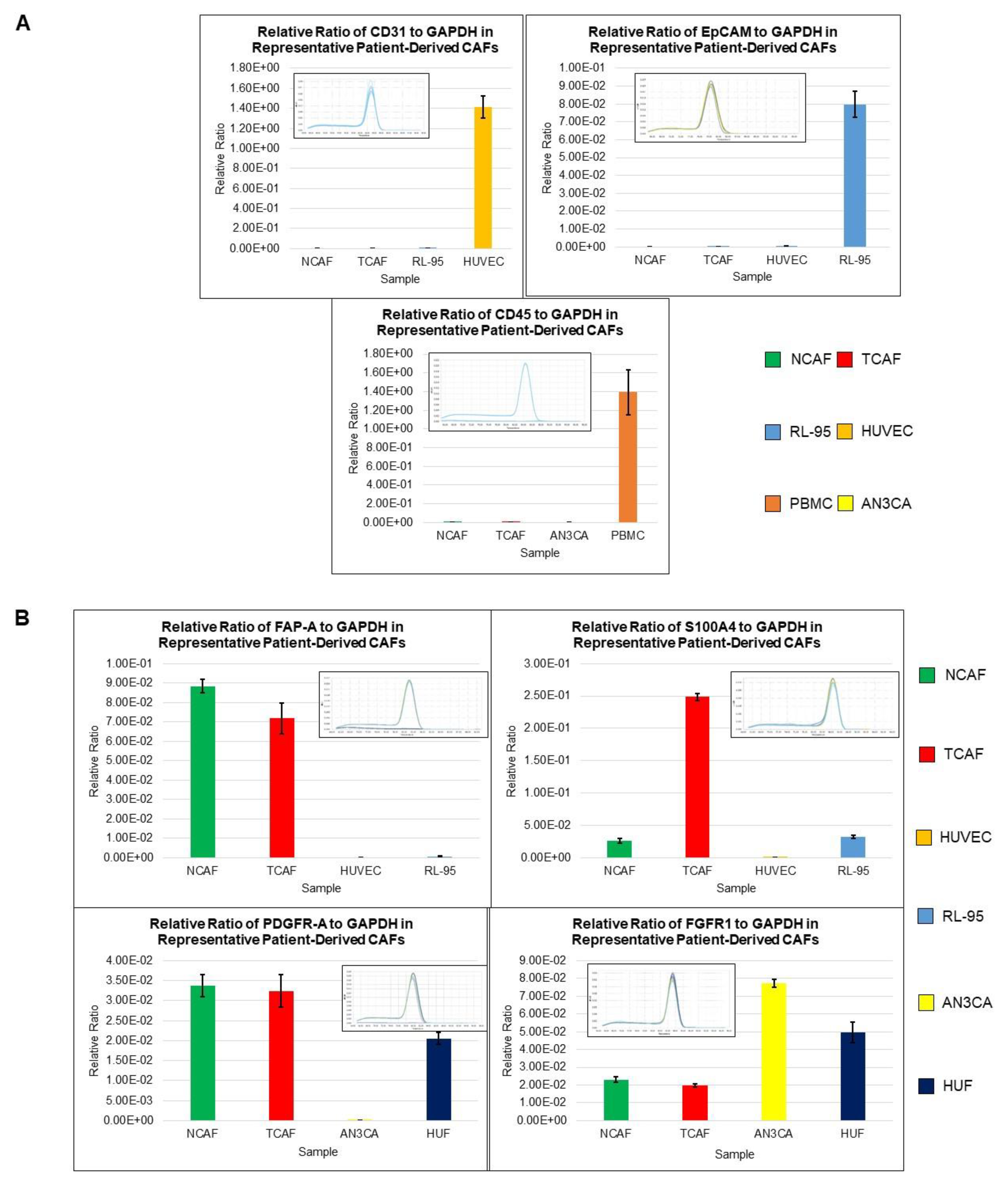
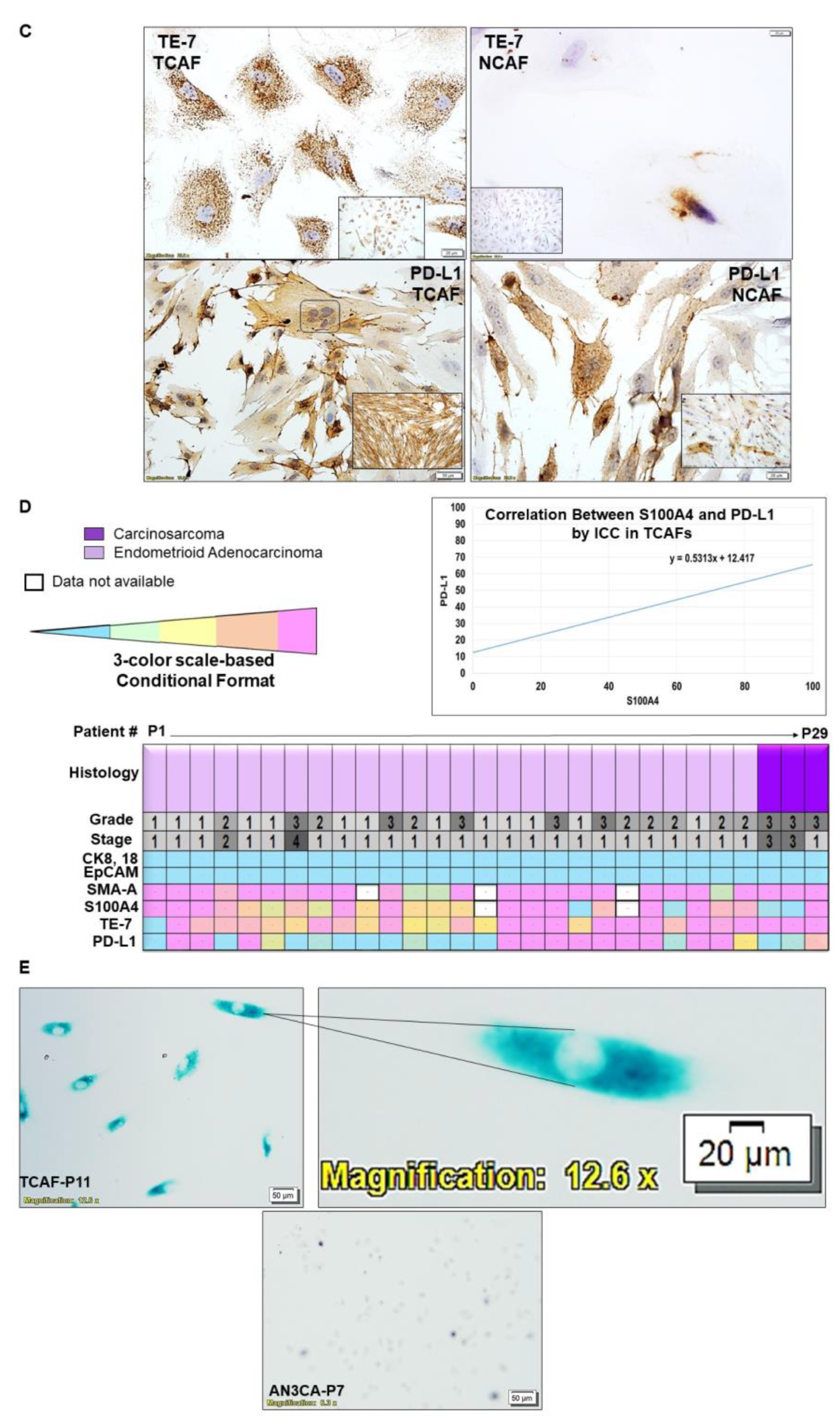
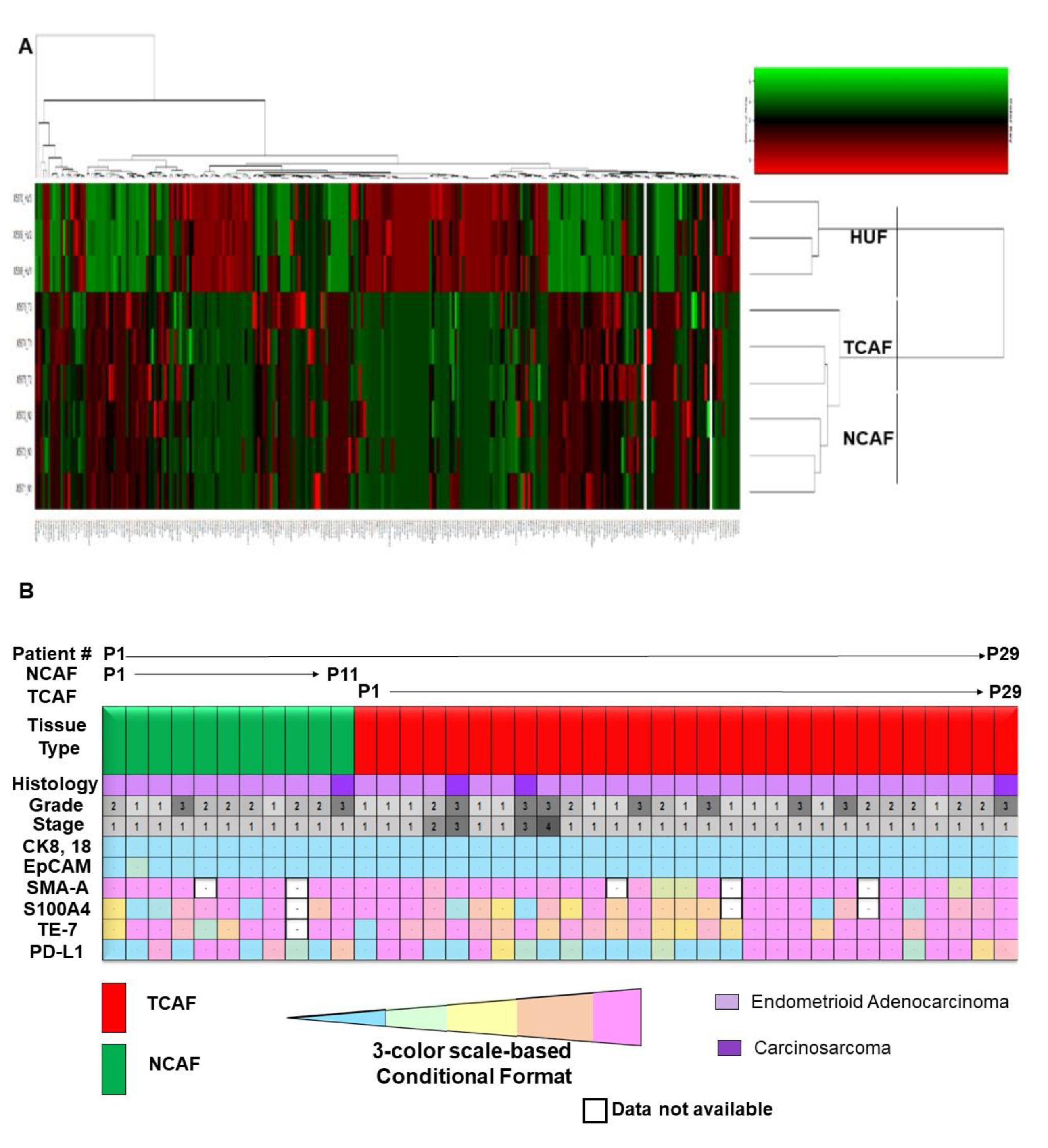
2.1. Characterization of Patient-Derived Endometrial CAFs
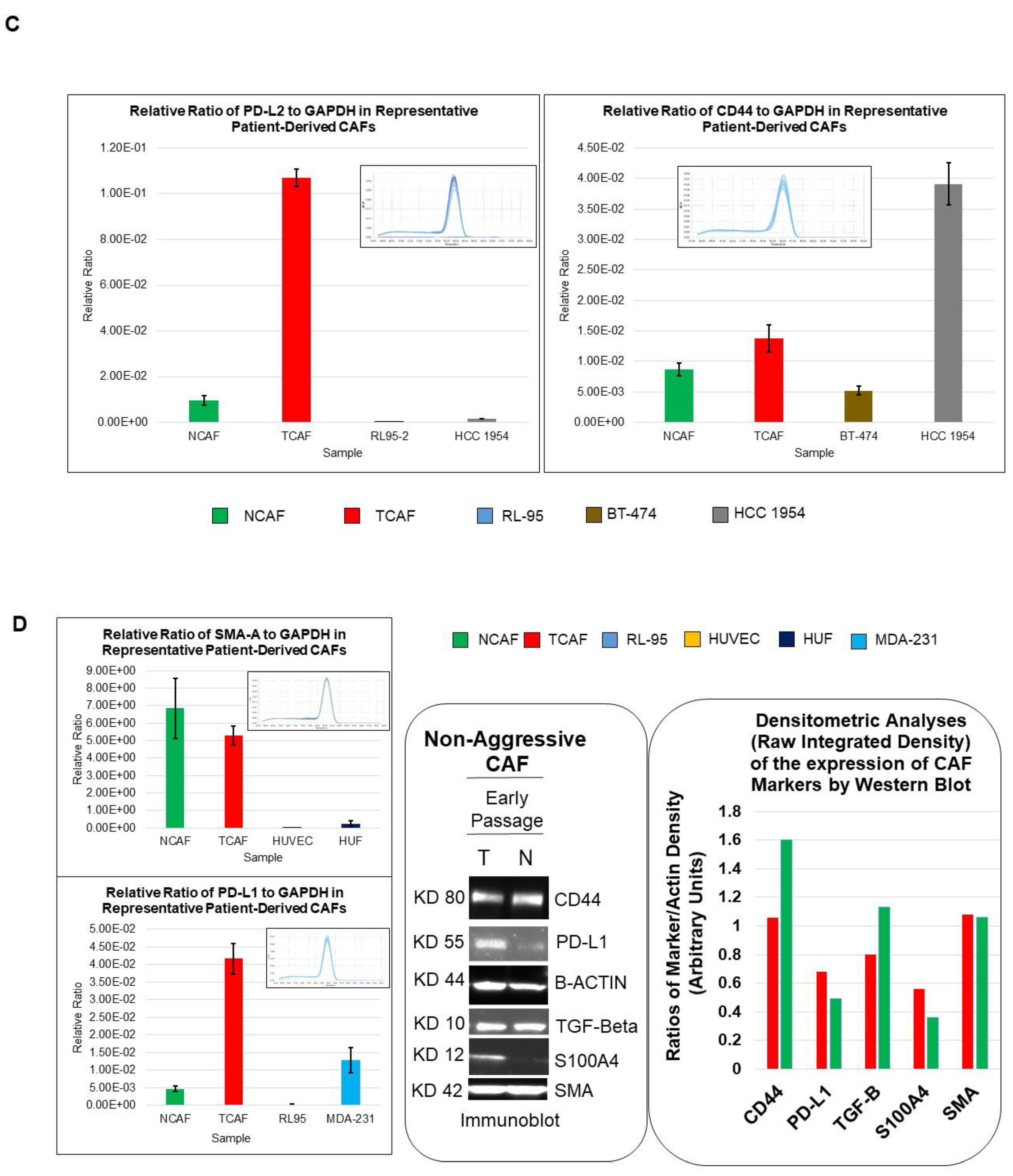
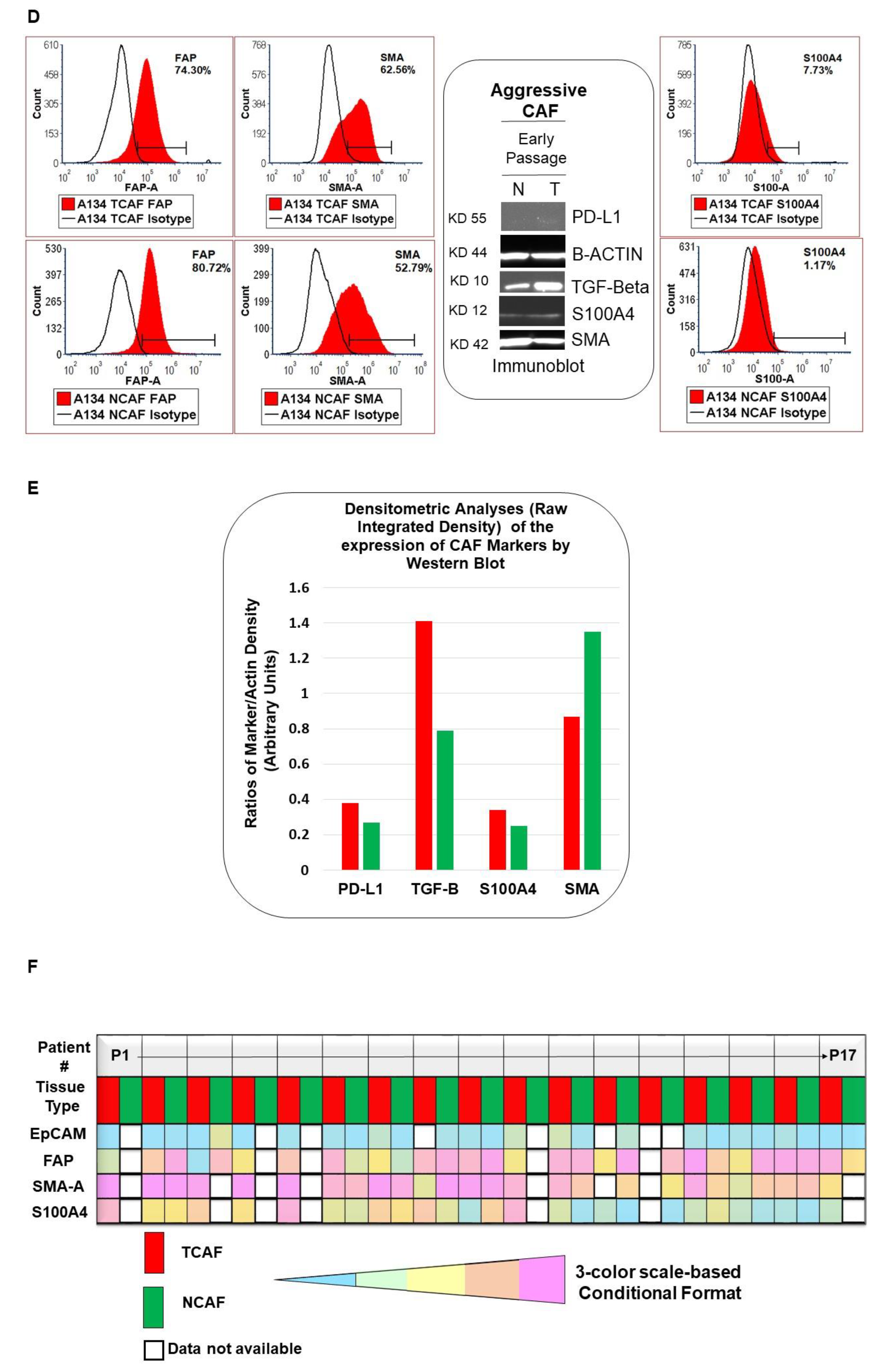
2.2. Designation of Aggressive and Non-Aggressive Endometrial CAFs
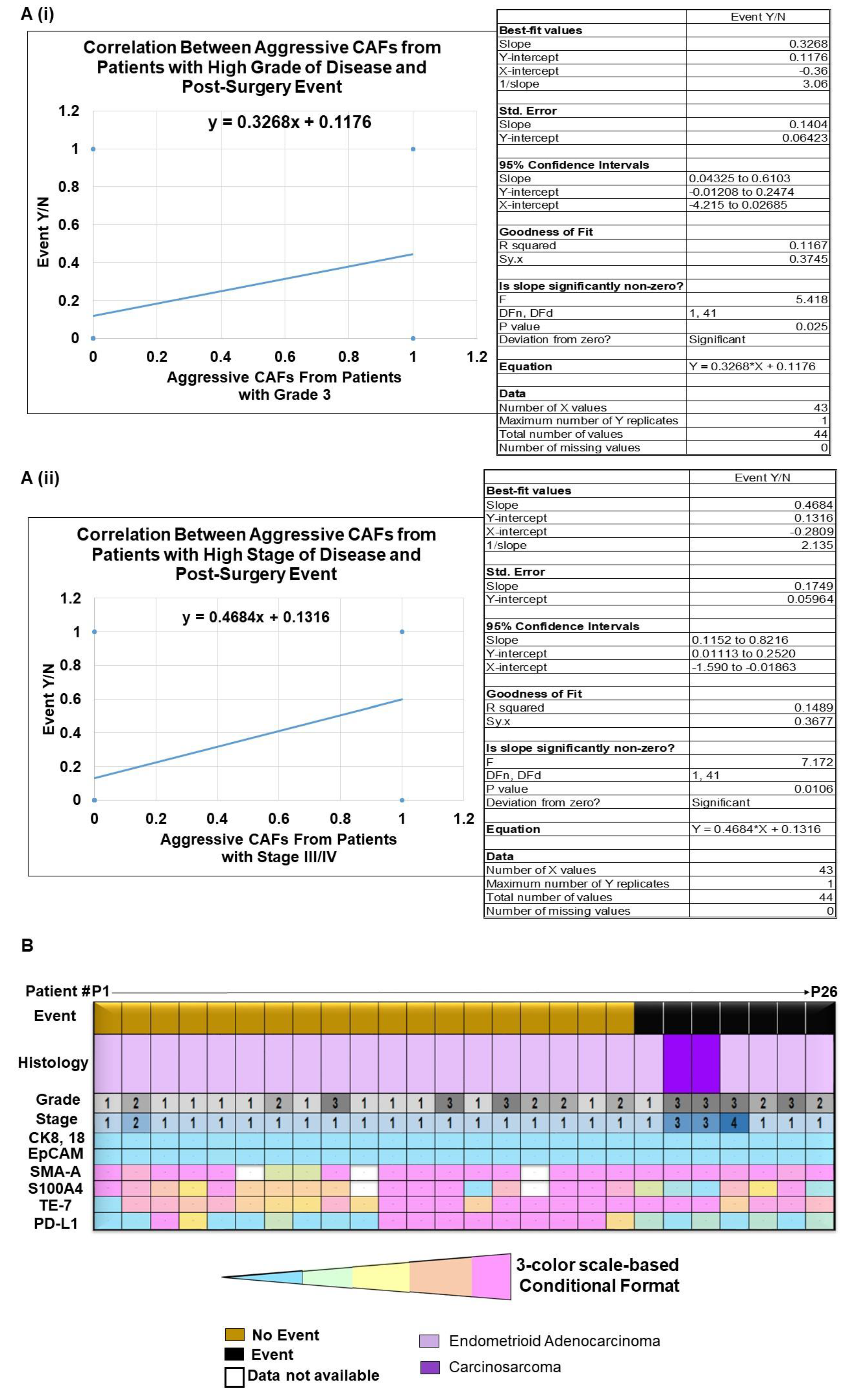
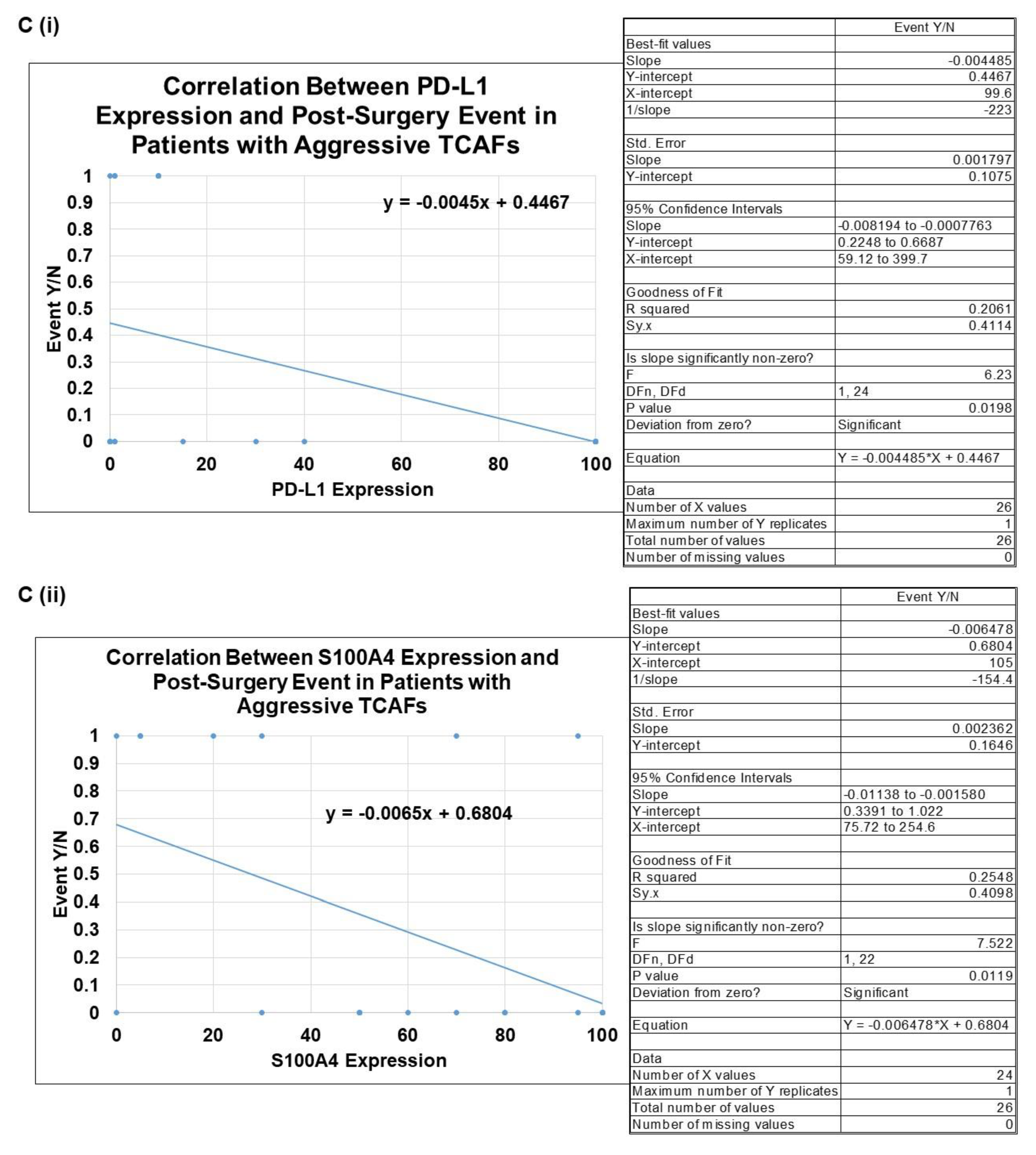
2.3. Clinical Relevance of Patient-Derived Aggressive Endometrial CAFs
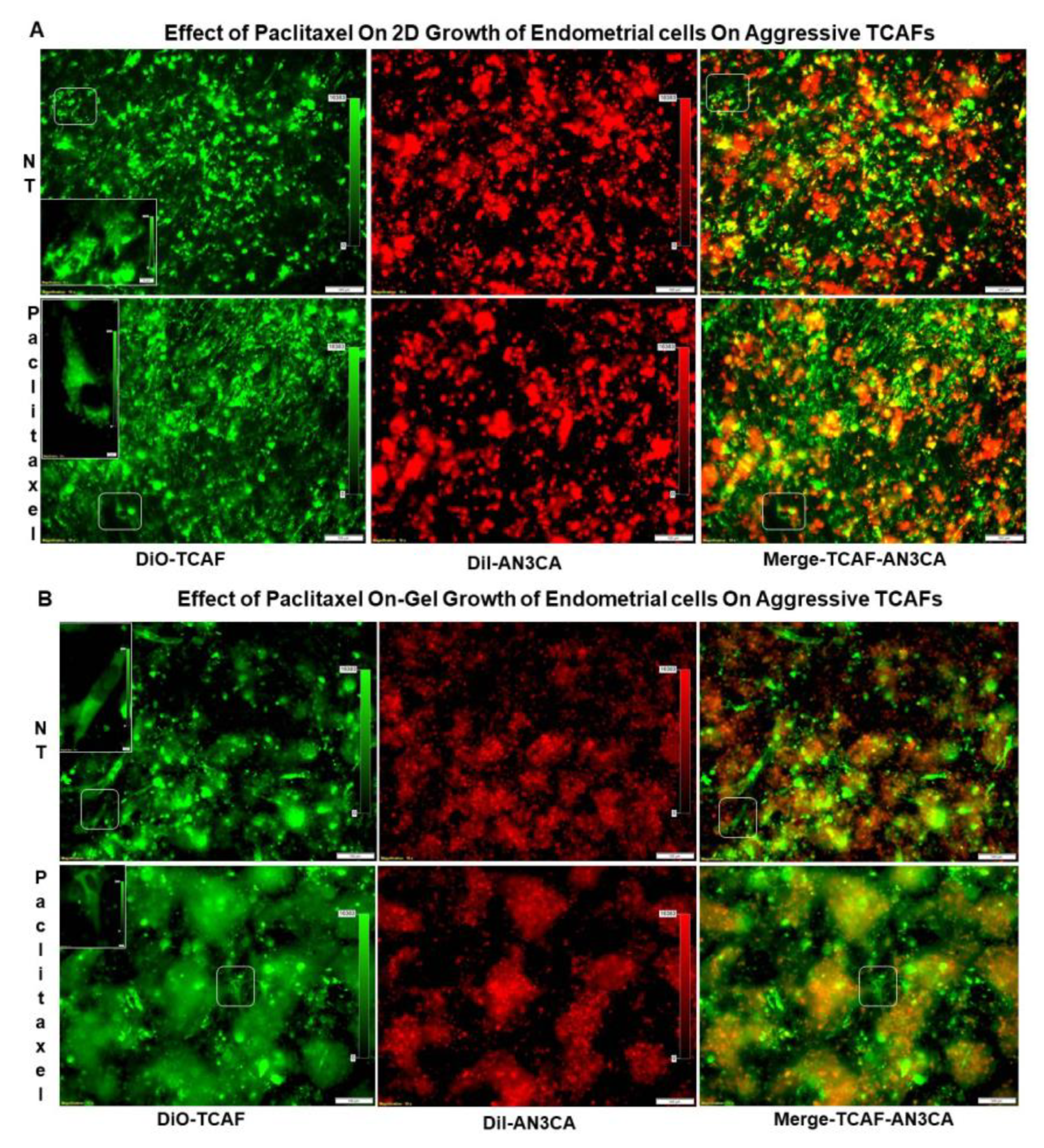
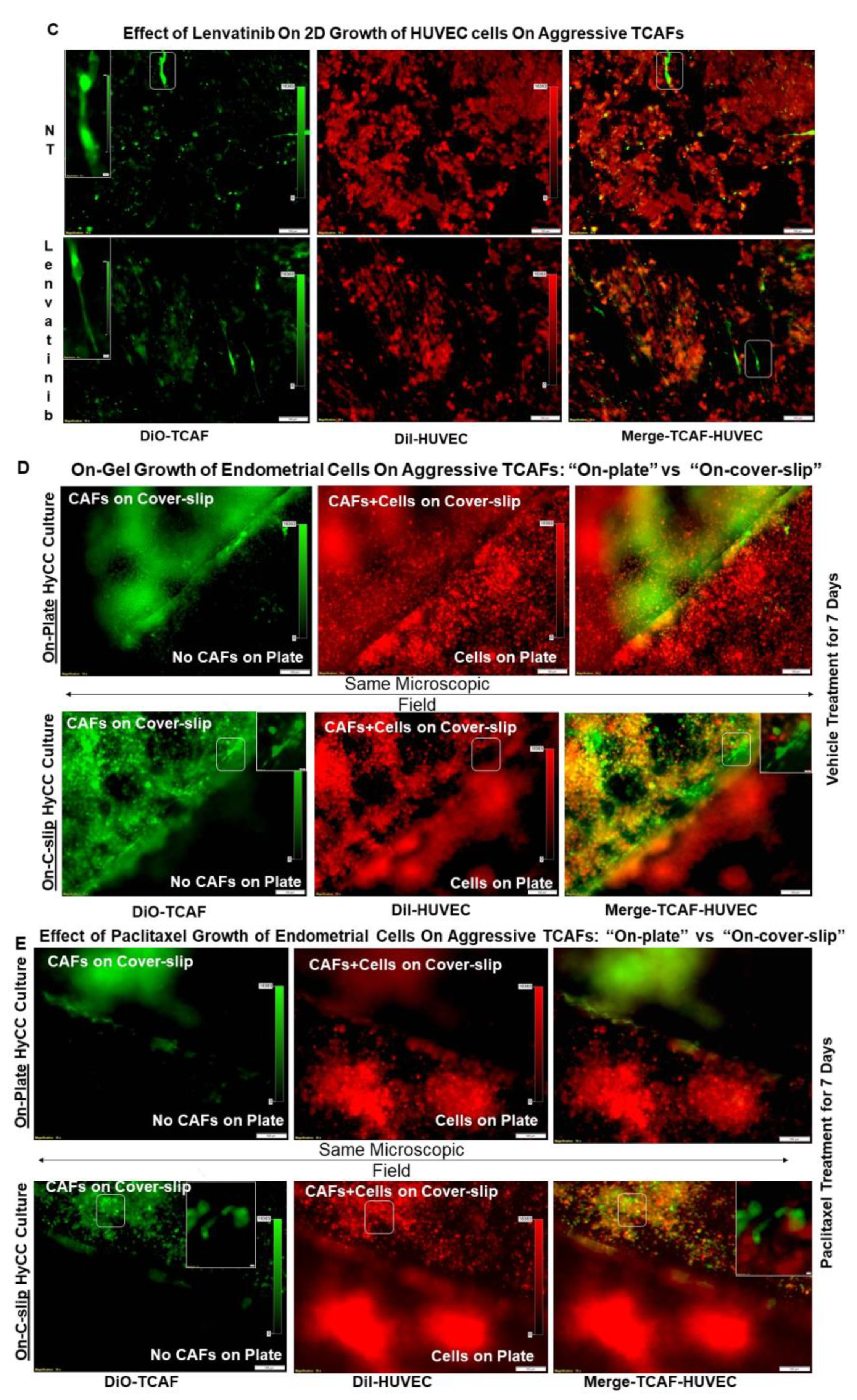
2.4. Growth Resistance Property of Aggressive Endometrial CAFs
3. Discussion
4. Methods and Materials
4.1. Tissue Collection at the Time of Surgery
4.2. Cell Lines and Reagents
4.3. Patient-Derived Primary Culture of Endometrial CAFs
4.4. Comprehensive Cancer Panel for Aggressive CAF Pair and HUF
4.5. Expression of mRNA for CAF Markers by qRT-PCR
4.6. Expression of Protein Markers of CAF by Flow-Cytometer and Western Blot
4.7. Cellular Localization of CAF Markers by ICC
4.8. Categorization of the Aggressiveness of CAFs
4.9. Post-Surgery Events in Endometrial Patients
4.10. Testing the Effect of Aggressive TCAFs from Patients with Post-Surgery Events on the 3D Clonogenic Growth of Endometrial Tumor Cells
4.11. Statistical Evaluation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koskas, M.; Amant, F.; Mirza, M.R.; Creutzberg, C.L. Cancer of the corpus uteri: 2021 update. Int. J. Gynaecol. Obstet. 2021, 155 (Suppl. S1), 45–60. [Google Scholar] [CrossRef] [PubMed]
- Crosbie, E.J.; Kitson, S.J.; McAlpine, J.N.; Mukhopadhyay, A.; Powell, M.E.; Singh, N. Endometrial cancer. Lancet 2022, 399, 1412–1428. [Google Scholar] [CrossRef] [PubMed]
- Brooks, R.A.; Fleming, G.F.; Lastra, R.R.; Lee, N.K.; Moroney, J.W.; Son, C.H.; Tatebe, K.; Veneris, J.L. Current recommendations and recent progress in endometrial cancer. CA Cancer J. Clin. 2019, 69, 258–279. [Google Scholar] [CrossRef] [PubMed]
- Connor, E.V.; Rose, P.G. Management Strategies for Recurrent Endometrial Cancer. Expert Rev. Anticancer Ther. 2018, 18, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Winterhoff, B.; Konecny, G.E. Targeting fibroblast growth factor pathways in endometrial cancer. Curr. Probl. Cancer 2017, 41, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K.; Dandara, C. Broadening Drug Design and Targets to Tumor Microenvironment? Cancer-Associated Fibroblast Marker Expression in Cancers and Relevance for Survival Outcomes. OMICS 2020, 24, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Pradip, D.; Jennifer, A.; Nandini, D. Cancer-Associated Fibroblasts in Conversation with Tumor Cells in Endometrial Cancers: A Partner in Crime. Int. J. Mol. Sci. 2021, 22, 9121. [Google Scholar] [CrossRef] [PubMed]
- De, P.; Aske, J.; Dey, N. Cancer-Associated Fibroblast Functions as a Road-Block in Cancer Therapy. Cancers 2021, 13, 5246. [Google Scholar] [CrossRef] [PubMed]
- Kennel, K.B.; Bozlar, M.; De Valk, A.F.; Greten, F.R. Cancer-associated fibroblasts in inflammation and anti-tumor immunity. Clin. Cancer Res. 2022, 29, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, R.; De, P.; Aske, J.C.; Lin, X.; Dale, A.; Vaselaar, E.; Koirala, N.; Ageton, C.; Gaster, K.; Plorde, J.; et al. A Laboratory-Friendly CTC Identification: Comparable Double-Immunocytochemistry with Triple-Immunofluorescence. Cancers 2022, 14, 2871. [Google Scholar] [CrossRef] [PubMed]
- Hoskins, P.J.; Swenerton, K.D.; Pike, J.A.; Wong, F.; Lim, P.; Acquino-Parsons, C.; Lee, N. Paclitaxel and carboplatin, alone or with irradiation, in advanced or recurrent endometrial cancer: A phase II study. J. Clin. Oncol. 2001, 19, 4048–4053. [Google Scholar] [CrossRef] [PubMed]
- Makker, V.; Colombo, N.; Casado Herraez, A.; Santin, A.D.; Colomba, E.; Miller, D.S.; Fujiwara, K.; Pignata, S.; Baron-Hay, S.; Ray-Coquard, I.; et al. Lenvatinib plus Pembrolizumab for Advanced Endometrial Cancer. N. Engl. J. Med. 2022, 386, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Kailasam, A.; Langstraat, C. Contemporary Use of Hormonal Therapy in Endometrial Cancer: A Literature Review. Curr. Treat. Options Oncol. 2022, 23, 1818–1828. [Google Scholar] [CrossRef] [PubMed]
- Kalampokas, E.; Giannis, G.; Kalampokas, T.; Papathanasiou, A.A.; Mitsopoulou, D.; Tsironi, E.; Triantafyllidou, O.; Gurumurthy, M.; Parkin, D.E.; Cairns, M.; et al. Current Approaches to the Management of Patients with Endometrial Cancer. Cancers 2022, 14, 4500. [Google Scholar] [CrossRef] [PubMed]
- De, P.; Carlson, J.H.; Jepperson, T.; Willis, S.; Leyland-Jones, B.; Dey, N. RAC1 GTP-ase signals Wnt-beta-catenin pathway mediated integrin-directed metastasis-associated tumor cell phenotypes in triple negative breast cancers. Oncotarget 2017, 8, 3072–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Patients | |
|---|---|
| Age group | # Patients (n = 53) |
| Below 55 | 7 |
| Above 55 | 46 |
| Pathology Parameters | |
| Histology | |
| Carcinosarcoma | 3 |
| Endometrioid Adenocarcinoma | 42 |
| Endometrial Cancer Not Otherwise Specified | 1 |
| High-Grade Papillary Serous Carcinoma | 1 |
| Mixed High-Grade Carcinoma | 3 |
| High-Grade Serous Carcinoma | 3 |
| Stage | |
| I | 40 |
| II | 2 |
| III | 8 |
| IV | 3 |
| Grade | |
| 1 | 27 |
| 2 | 11 |
| 3 | 15 |
| Myometrial Invasion % | |
| 0–25 | 24 |
| 26–50 | 19 |
| 51–75 | 4 |
| 76–100 | 6 |
| Lymphovascular Invasion | |
| + | 13 |
| − | 39 |
| Indeterminate | 1 |
| Lymph Node Positivity | |
| + | 11 |
| − | 35 |
| None Submitted | 7 |
| Patients with Aggressive CAF | |
|---|---|
| Histology | |
| Carcinosarcoma | 2 |
| Endometrioid Adenocarcinoma | 26 |
| Endometrial Cancer Not Otherwise Specified | 1 |
| High-Grade Papillary Serous Carcinoma | 0 |
| Mixed High-Grade Carcinoma | 1 |
| High Grade Serous Carcinoma | 1 |
| Stage | |
| I | 25 |
| II | 1 |
| III | 3 |
| IV | 2 |
| Grade | |
| 1 | 14 |
| 2 | 8 |
| 3 | 9 |
| Myometrial Invasion % | |
| 0–25 | 11 |
| 26–50 | 14 |
| 51–75 | 2 |
| 76–100 | 4 |
| Lymphovascular Invasion | |
| + | 5 |
| − | 25 |
| Indeterminate | 1 |
| Lymph Node Positivity | |
| + | 5 |
| − | 23 |
| None Submitted | 3 |
| Patients with Non-Aggressive CAF | |
| Histology | |
| Carcinosarcoma | 1 |
| Endometrioid Adenocarcinoma | 9 |
| Endometrial Cancer Not Otherwise Specified | 0 |
| High Grade Papillary Serous Carcinoma | 1 |
| Mixed High-Grade Carcinoma | 1 |
| High-Grade Serous Carcinoma | 1 |
| Stage | |
| I | 9 |
| II | 0 |
| III | 3 |
| IV | 1 |
| Grade | |
| 1 | 8 |
| 2 | 1 |
| 3 | 4 |
| Myometrial Invasion % | |
| 0–25 | 7 |
| 26–50 | 3 |
| 51–75 | 1 |
| 76–100 | 2 |
| Lymphovascular Invasion | |
| + | 5 |
| − | 8 |
| Indeterminate | 0 |
| Lymph Node Positivity | |
| + | 4 |
| − | 5 |
| None Submitted | 4 |
| Characteristics of Patients with Aggressive CAF and Post-Surgery Events | |||||||
|---|---|---|---|---|---|---|---|
| # | Treatment | Post-Surgery Events | Pathology Parameter(s) | Genomics | |||
| Neo-Adjuvant Treatment | Adjuvant Treatment | Specific Event | Time of Event (Months) | OS (Overall Survival in Months) | |||
| 1 | None | None | Developed “squamous cell carcinoma of the skin” | 25–37 | NA | Endometrioid Adenocarcinoma | Not performed |
| Grade 1 | |||||||
| Stage I | |||||||
| LVI Negative | |||||||
| Lymph Node: Negative | |||||||
| Myometrial Invasion: 30% | |||||||
| Mismatch Repair Competent | |||||||
| 2 | None | Carboplatinum + Paclitaxel x6. Radiation post-chemo. | Vaginal bleeding, biopsy demonstrated recurrent disease. | 10 | 14 | Carcino-sarcoma | PTEN: p.L186fs Frameshift. Variant Allele Fraction 52.1% CTNNB1: p.S33F Missense variant. Variant Allele Fraction 34.2% PTEN: p.R130G Missense variant. Variant Allele Fraction 22.9% ARID1A: p.P109fs Frameshift. Variant Allele Fraction 10.4% |
| Grade 3 | |||||||
| Stage III | |||||||
| LVI Positive | |||||||
| Lymph Node: Positive | |||||||
| Myometrial Invasion: 72% | |||||||
| Mismatch Repair Competent | |||||||
| 3 | None | Carboplatinum + Paclitaxel x6. Radiation post-chemo. | CT scan demonstrated lymph nodes concerning for metastatic disease, biopsy confirmed recurrence. | 19 | NA | Carcino-sarcoma | PIK3CA: p.Q546K Missense Variant. Variant Allele Fraction 68% CDKN2A: p.P81L Missense Variant. Variant Allele Fraction 4% TP53: p.K132R Missense Variant. Variant Allele Fraction 53% FBXW7: p.R505C Missense Variant. Variant Allele Fraction 20% KMT2D: p.Q2811Sfs*40 Frameshift. Variant Allele Fraction 16% SLX4: p.Y255* Nonsense variant. Variant Allele Fraction 17% |
| Grade 3 | |||||||
| Stage III | |||||||
| LVI Negative | |||||||
| Lymph Node: Positive | |||||||
| Myometrial Invasion: 38% | |||||||
| Mismatch Repair Competent | |||||||
| 4 | None | Carboplatinum + Paclitaxel x6. | Patient had worsening symptoms and was found to have moderate volume of ascites and peritoneal nodularity. CT scan showed changes concerning for progression. | 10 | 13 | Endome-trioid Adeno-carcinoma | PIK3CA: p.H1047R Missense variant (exon 20). Variant Allele Fraction 52.8% PTEN: p.E288* Stop gain. Variant Allele Fraction 71% TP53: p.Y234C Missense Variant. Variant Allele Fraction 68.5% NF1: Copy Number Loss KHDRBS3-TP63: Chromosomal rearrangement |
| Grade 3 | |||||||
| Stage IV | |||||||
| LVI Negative | |||||||
| Lymph Node: None Submitted | |||||||
| Myometrial Invasion: 50% | |||||||
| Mismatch Repair Competent | |||||||
| 5 | None | None | Patient presented with hematuria and worsening peripheral edema. There was concern for recurrent disease but patient passed away before definitive diagnosis. | 11 | 11 | Endome-trioid Adeno-carcinoma | Not performed |
| Grade 2 | |||||||
| Stage I | |||||||
| LVI Negative | |||||||
| Lymph Node: Negative | |||||||
| Myometrial Invasion: 44% | |||||||
| Mismatch Repair Competent | |||||||
| 6 | None | Radiation. | Patient presented with vaginal spotting, biopsy revealed recurrent disease. CT scan demonstrated widespread metastatic recurrence. | 27 | NA | Endome-trioid Adeno-carcinoma | PIK3CA: p.Q546H Missense variant. Variant Allele Fraction 55% 17q12q21.2 (Focal—ERBB2): Gain, Copies = 3.5 SMARCA4: p.L410Tfs*90 Frameshift. Variant Allele Fraction 28% SPOP: p.R121Q Missense variant. Variant Allele Fraction 59% TP53: p.J195F Missense variant. Variant Allele Fraction 46% 17p13.3q12 (whole 17p, part 17q—TP53): Loss, Copies = 1.5 10q23.1q23.32 (Sub-arm—PTEN): Loss, Copies = 1.5 |
| Grade 3 | |||||||
| Stage I | |||||||
| LVI Negative | |||||||
| Lymph Node: Negative | |||||||
| Myometrial Invasion: 95% | |||||||
| Mismatch Repair Competent | |||||||
| 7 | None | Radiation. | Patient had lesion on top of vagina, biopsy revealed recurrent disease. | 8 | NA | Endome-trioid Adeno-carcinoma | Invitae—Negative |
| Grade 2 | |||||||
| Stage I | |||||||
| LVI Negative | |||||||
| Lymph Node: Negative | |||||||
| Myometrial Invasion: 6% | |||||||
| Mismatch Repair Deficient | |||||||
| Primers for qRT-PCR | ||
|---|---|---|
| Gene | Primer Sequence (Sequences Listed 5′–3′) | |
| ACTA-2/SMA | F: CGT TAC TAC TGC TGA GCG TGA | |
| R: GCC CAT CAG GCA ACT CGT AA | ||
| CD31 | F: ATT GCA GTG GTT ATC ATC GGA GTG | |
| R: CTG GTT GTT GGA GTT CAG AAG TGG | ||
| CD44 | F: AGC ACT TCA GGA GGT TAC ATC T | |
| R: CTT GCC TCT TGG TTG CTG TCT | ||
| CD45 | F: CTTCAGTGGTCCCATTGTGGTG | |
| R: CCACTTTGTTCTCGGCTTCCAG | ||
| CD90/THY1 | F: GAAGGTCCTCTACTTATCCGCC | |
| R: TGATGCCCTCACACTTGACCAG | ||
| EpCAM | F: AGC GAG TGA GAA CCT ACT GGA | |
| R: CGC GTT GTG ATC TCC TTC TGA | ||
| FAP-A | F: GGA AGT GCC TGT TCC AGC AAT G | |
| R: TGT CTG CCA GTC TTC CCT GAA G | ||
| GAPDH | F: TCA AGG CTG AGA ACG GGA AG | |
| R: CGC CCC ACT TGA TTT TGG AG | ||
| FGFR1 | F: GAC ACC ACC TAC TTC TCC GTC AA | |
| R: CAA TAT GGA GCT ACG GGC ATA CG | ||
| PDGFRA | F: TGG CAG TAC CCC ATG TCT GAA | |
| R: CCA AGA CCG TCA CAA AAA GGC | ||
| PD-L1 | F: ACC TAC TGG CAT TTG CTG AAC G | |
| R: ATA GAC AAT TAG TGC AGC CAG GT | ||
| S100A4 | F: CAG AAC TAA AGG AGC TGC TGA CC | |
| R: CTT GGA AGT CCA CCT CGT TGT C | ||
| Antibodies for ICC | ||
| Antibody | Manufacturer | Cat.# |
| Cytokeratin 8 & 18 (B22.1 & B23.1) | Cell Marque | 818M-94 |
| Ep-CAM/Epithelial Specific Antigen (Ber-EP4) | Cell Marque | 248M-94 |
| Fibroblasts Antibody (TE-7) | NOVUS | NBP2-50082 |
| Actin, Smooth Muscle (1A4) | Cell Marque | 202M-94 |
| Recombinant Anti-S100A4 Antibody | Abcam (Waltham, MA, USA) | ab124805 |
| PD-L1 [Clone 22C3] | Agilent-Dako | M365329-1 |
| PD-L2 (D7U8C) | Cell Signaling | 82723 |
| CD31 | Cell Signaling | 3528 |
| Vimentin (SP20) Rabbit Monoclonal Antibody | Cell Marque | 347R-14 |
| Antibodies for Flow Cytometry | ||
| Antibody | Manufacturer | Cat.# |
| CD31-FITC | Miltenyi (Waltham, MA, USA) | 130-117-390 |
| CD155-PE | Miltenyi | 130-105-846 |
| CD90 PE-Vio615 | Miltenyi | 130-114-909 |
| S100A4-PerCP | NOVUS | NBP2-36431APCCY7 |
| SMA-FITC | NOVUS | NBP2-34522F |
| FAP-PE | R&D Systems (McKinley Place NE, Minneapolis, MN, USA) | FAB3715P-025 |
| PD-L1-APC | Miltenyi | 130-122-816 |
| EpCAM-APC | Miltenyi | 130-133-260 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulaiman, R.; De, P.; Aske, J.C.; Lin, X.; Dale, A.; Gaster, K.; Espaillat, L.R.; Starks, D.; Dey, N. Characterization and Clinical Relevance of Endometrial CAFs: Correlation between Post-Surgery Event and Resistance to Drugs. Int. J. Mol. Sci. 2023, 24, 6449. https://doi.org/10.3390/ijms24076449
Sulaiman R, De P, Aske JC, Lin X, Dale A, Gaster K, Espaillat LR, Starks D, Dey N. Characterization and Clinical Relevance of Endometrial CAFs: Correlation between Post-Surgery Event and Resistance to Drugs. International Journal of Molecular Sciences. 2023; 24(7):6449. https://doi.org/10.3390/ijms24076449
Chicago/Turabian StyleSulaiman, Raed, Pradip De, Jennifer C. Aske, Xiaoqian Lin, Adam Dale, Kris Gaster, Luis Rojas Espaillat, David Starks, and Nandini Dey. 2023. "Characterization and Clinical Relevance of Endometrial CAFs: Correlation between Post-Surgery Event and Resistance to Drugs" International Journal of Molecular Sciences 24, no. 7: 6449. https://doi.org/10.3390/ijms24076449
APA StyleSulaiman, R., De, P., Aske, J. C., Lin, X., Dale, A., Gaster, K., Espaillat, L. R., Starks, D., & Dey, N. (2023). Characterization and Clinical Relevance of Endometrial CAFs: Correlation between Post-Surgery Event and Resistance to Drugs. International Journal of Molecular Sciences, 24(7), 6449. https://doi.org/10.3390/ijms24076449