
Biofilm-Based Biocatalysis for Galactooligosaccharides Production by the Surface Display of β-Galactosidase in Pichia pastoris


Abstract
:1. Introduction
2. Results and Discussion
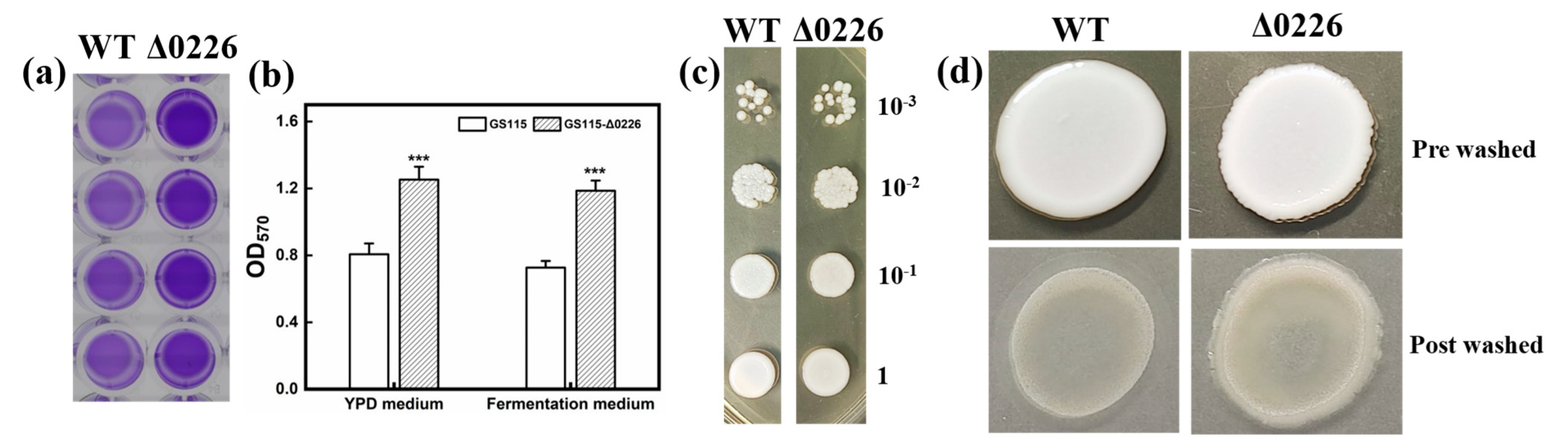
2.1. The Impact of PAS_chr1-3_0226 Gene Deletion on Biofilm Formation in P. pastoris
2.2. Biofilm Formation on Carriers and Cell Morphological Changes
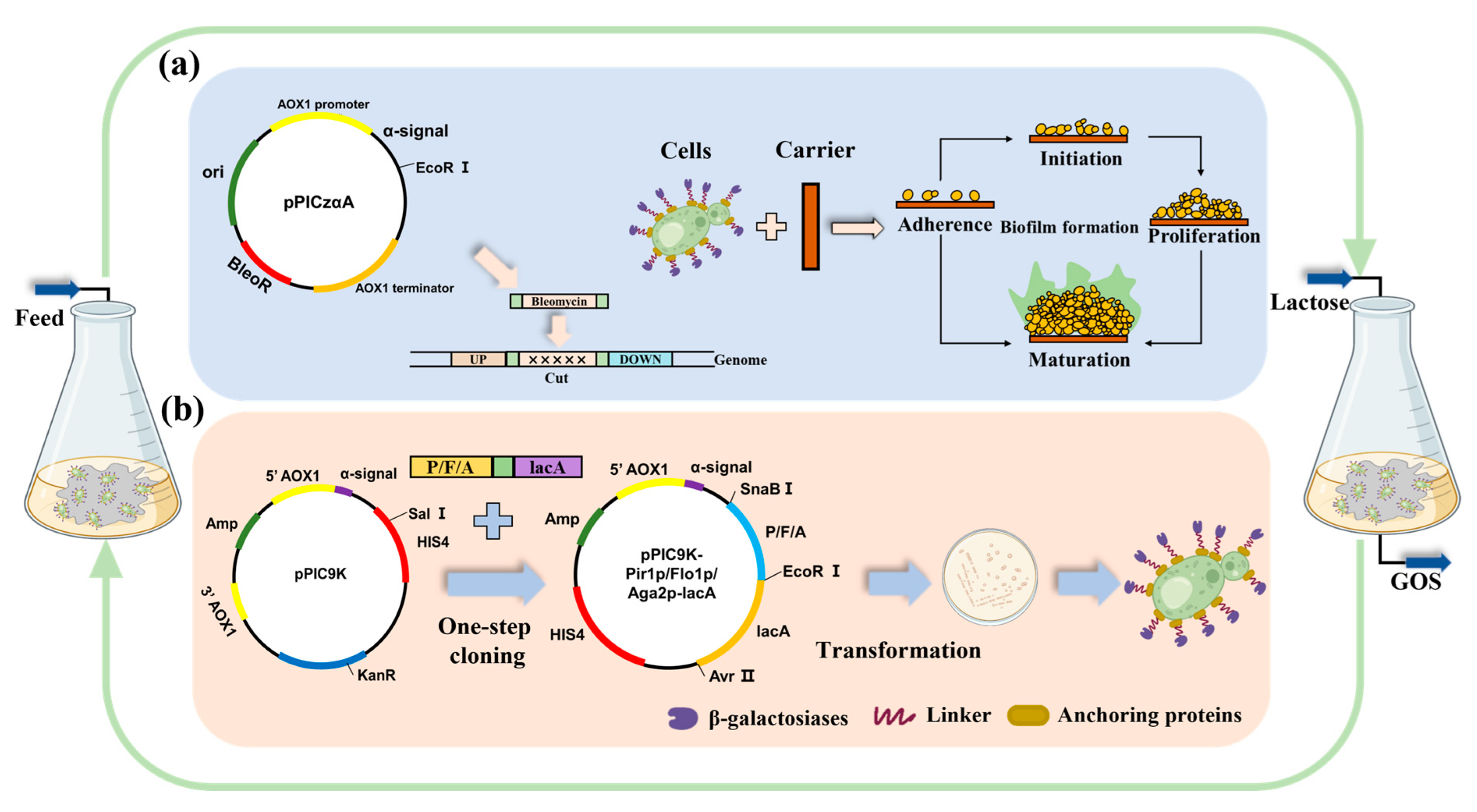
2.3. Construction of Strains for Surface Display
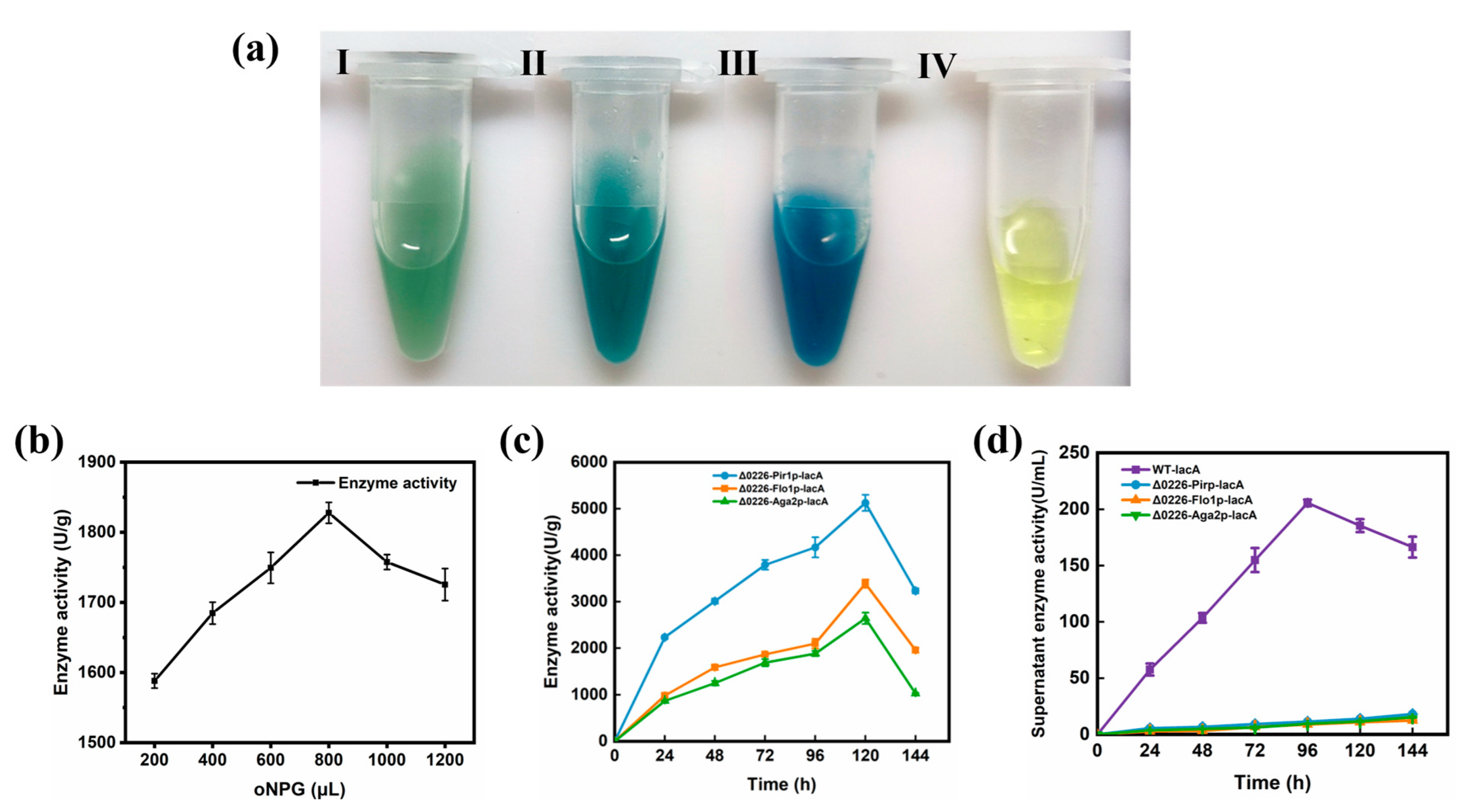
2.4. Expression of Surface-Displayed β-Galactosidase
2.5. Enzyme Activity of Surface-Displayed β-Galactosidase
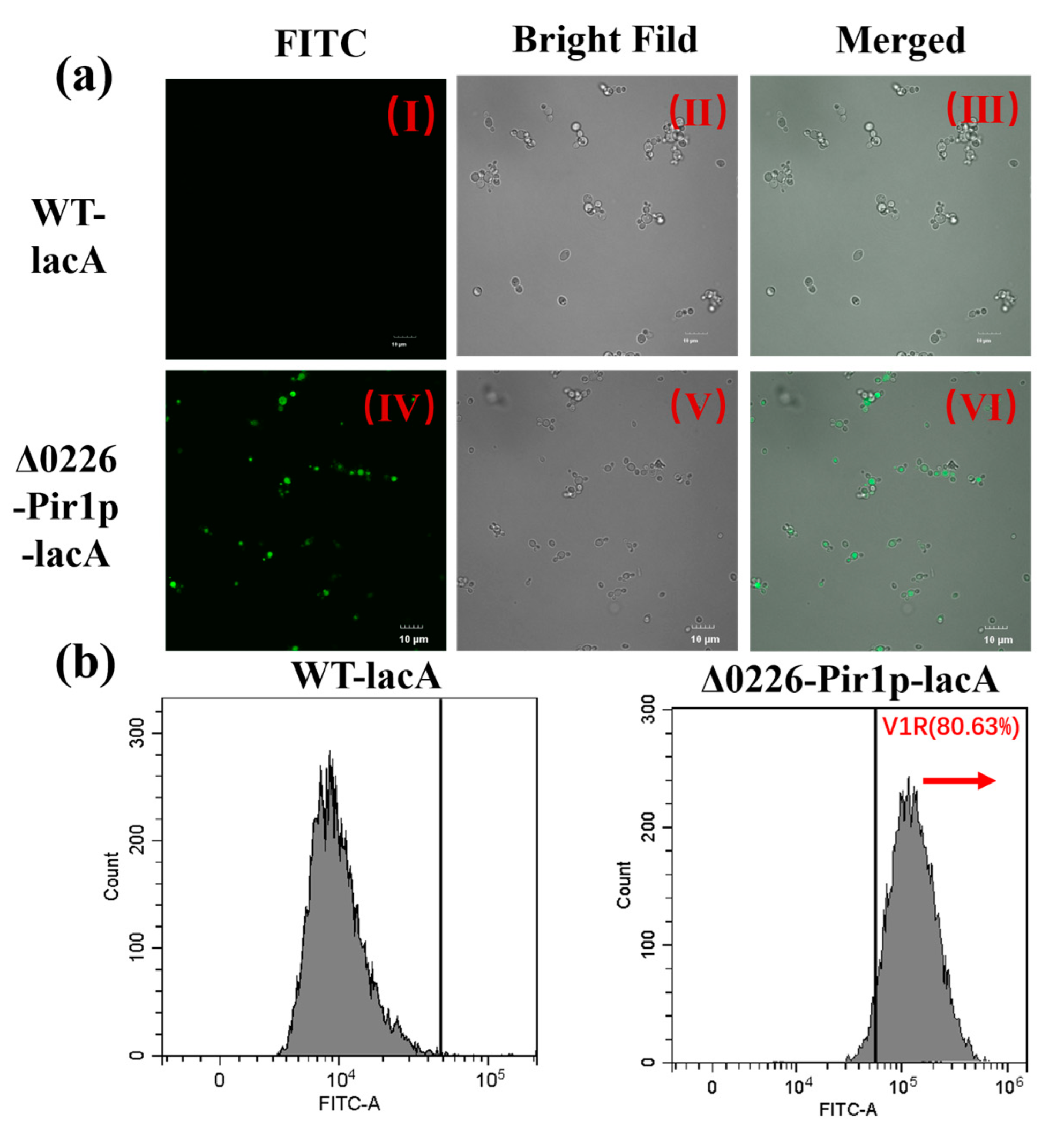
2.6. Verification of Surface-Displayed β-Galactosidase by Immunofluorescence Microscopy and Flow Cytometry
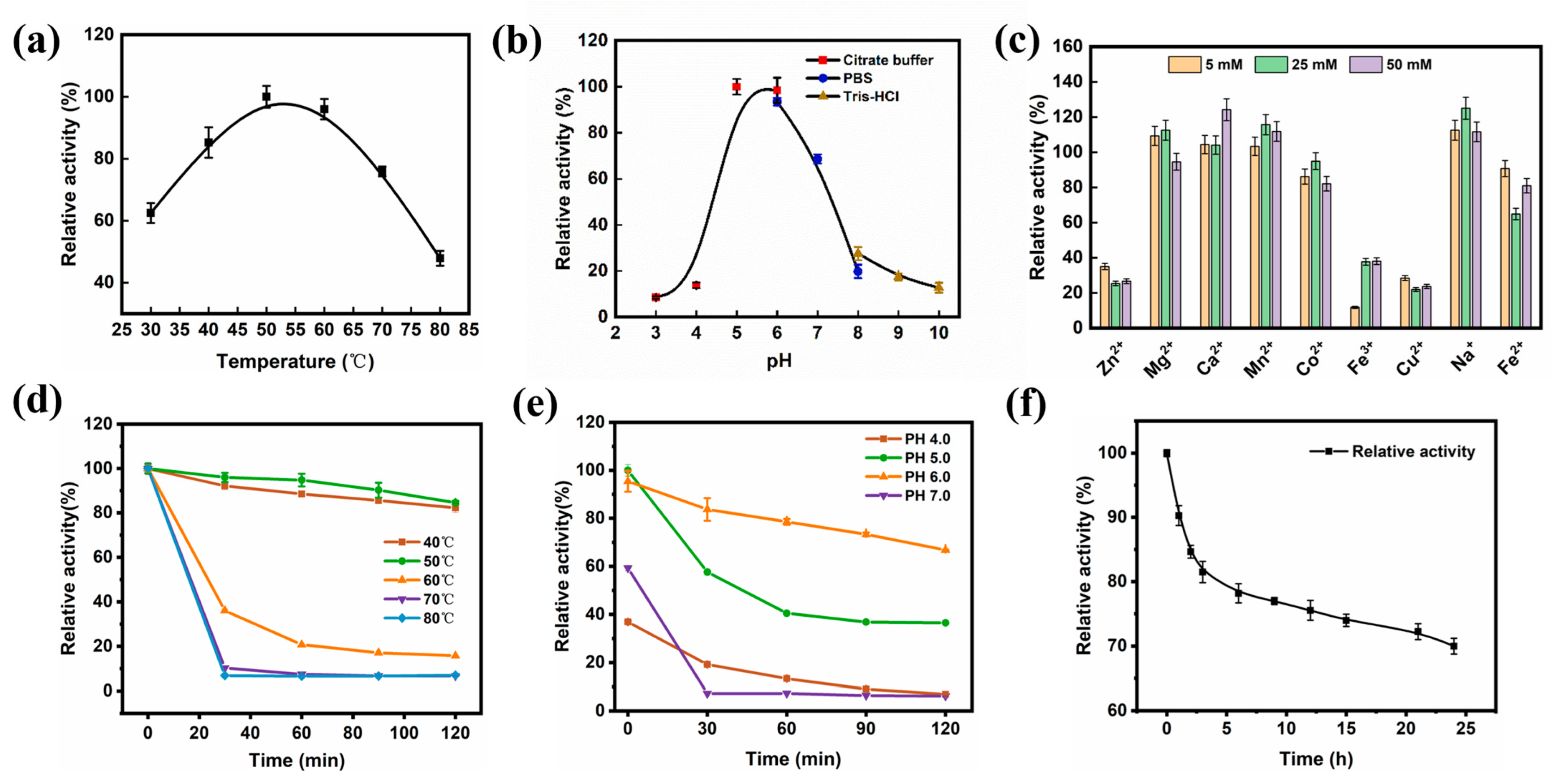
2.7. Characterization of Surface-Displayed β-Galactosidase Activity
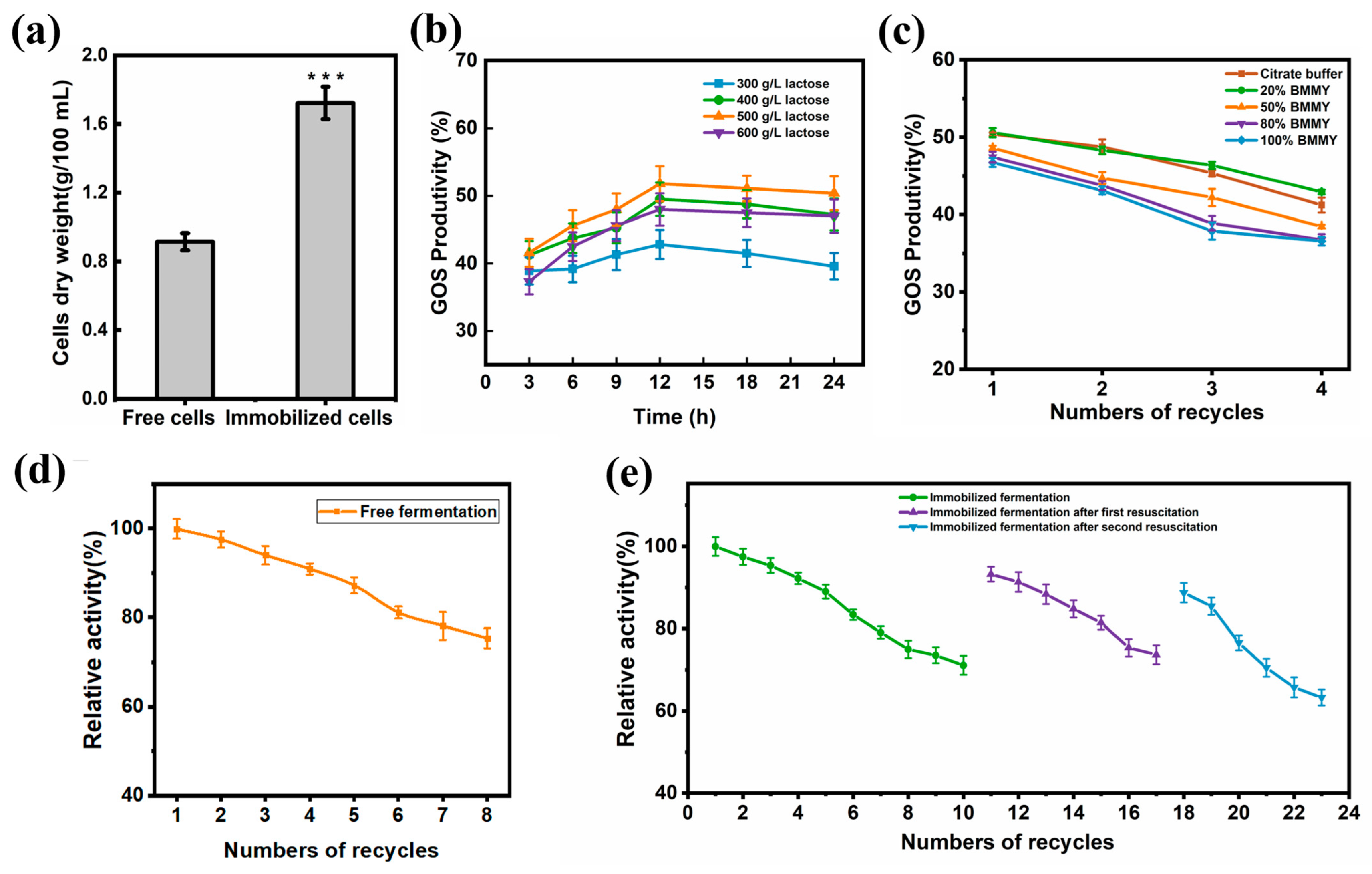
2.8. Optimizing the Process of Increasing GOS Yields
2.9. Free and Immobilized Cell Biocatalytic GOS Production
3. Materials and Methods
3.1. Strains and Plasmids
3.2. Media and Growth Conditions
3.3. Construction of Gene Knock-Out and Surface-Displayed Strains
3.4. Characterization of Biofilm Formation
3.4.1. Biofilm Formation on Plastics
3.4.2. Cell Growth and Infiltration Capacity Analysis
3.4.3. Characterization of Scanning Electron Microscopy and Transmission Electron Microscopy
3.5. Surface-Displayed β-Galactosidase Assay
3.5.1. β-Galactosidase In Situ Staining Assay
3.5.2. β-Galactosidase Enzyme Activity
3.5.3. Immunofluorescence Microscopy and Flow Cytometry Analysis
3.5.4. Enzymatic Properties of β-Galactosidase
3.6. Free and Immobilized Fermentation for GOS Synthesis
- ① Free cells (g/L/h) =
- ② Immobilized cells (g/L/h) =
3.7. The Methods of Product Detection
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tzortzis, G.; Vulevic, J. Galacto-Oligosaccharide Prebiotics. In Prebiotics and Probiotics Science and Technology; Charalampopoulos, D., Rastall, R.A., Eds.; Springer: New York, NY, USA, 2009; pp. 207–244. [Google Scholar]
- Nawaz, A.; Bakhsh Javaid, A.; Irshad, S.; Hoseinifar, S.H.; Xiong, H. The functionality of prebiotics as immunostimulant: Evidences from trials on terrestrial and aquatic animals. Fish Shellfish. Immunol. 2018, 76, 272–278. [Google Scholar] [CrossRef]
- Panesar, P.S.; Kaur, R.; Singh, R.S.; Kennedy, J.F. Biocatalytic strategies in the production of galacto-oligosaccharides and its global status. Int. J. Biol. Macromol. 2018, 111, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.; Lee, V.; Sørensen, K.I.; Gänzle, M.G. Production of galactooligosaccharides and heterooligosaccharides with disrupted cell extracts and whole cells of lactic acid bacteria and bifidobacteria. Int. Dairy J. 2011, 21, 748–754. [Google Scholar] [CrossRef]
- Güleç, H.A.; Gürdaş, S.; Albayrak, N.; Mutlu, M. Immobilization of Aspergillus oryzae β-galactosidase on low-pressure plasma-modified cellulose acetate membrane using polyethyleneimine for production of galactooligosaccharide. Biotechnol. Bioprocess Eng. 2010, 15, 1006–1015. [Google Scholar] [CrossRef]
- Onishi, N.; Tanaka, T. Purification and Properties of a Galacto- and Gluco-Oligosaccharide-Producing P-Glycosidase from Rhodotorula minuta IF0879. J. Ferment. Bioeng. 1996, 82, 439–443. [Google Scholar] [CrossRef]
- Kaczyński, L.K.; Cais-Sokolińska, D.; Szwengiel, A. Kinetics of lactose hydrolysis and galactooligosaccharides formation in beverages based on goat’s milk and its permeate. Food Sci. Biotechnol. 2019, 28, 1529–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loo, J.; Cummings, J.; Delzenne, N.; Englyst, H.; Franck, A.; Hopkins, M.; Kok, N.; Macfarlane, G.; Newton, D.; Quigley, M.; et al. Functional food properties of non-digestible oligosaccharides: A consensus report from the ENDO project (DGXII AIRII-CT94-1095). Br. J. Nutr. 1999, 81, 121–132. [Google Scholar] [PubMed] [Green Version]
- Vera, C.; Córdova, A.; Aburto, C.; Guerrero, C.; Suárez, S.; Illanes, A. Synthesis and purification of galacto-oligosaccharides: State of the art. World J. Microbiol. Biotechnol. 2016, 32, 197. [Google Scholar] [CrossRef]
- Guidini, C.Z.; Fischer, J.; Santana, L.N.S.; Cardoso, V.L.; Ribeiro, E.J. Immobilization of Aspergillus oryzae β-galactosidase in ion exchange resins by combined ionic-binding method and cross-linking. Biochem. Eng. J. 2010, 52, 137–143. [Google Scholar] [CrossRef]
- Tanaka, T.; Yamada, R.; Ogino, C.; Kondo, A. Recent developments in yeast cell surface display toward extended applications in biotechnology. Appl. Microbiol. Biotechnol. 2012, 95, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Xu, Y.; Wang, K.; Zheng, S. Synthesis of Isomalto-Oligosaccharides by Pichia pastoris Displaying the Aspergillus niger α-Glucosidase. J. Agric. Food Chem. 2017, 65, 9468–9474. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Y.; Liu, D.; Zhao, N.; Cheng, H.; Ren, H.; Guo, T.; Niu, H.; Zhuang, W.; Wu, J.; et al. Involvement of glycolysis/gluconeogenesis and signaling regulatory pathways in Saccharomyces cerevisiae biofilms during fermentation. Front. Microbiol. 2015, 6, 139. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, J.R.; Mitchell, A.P. How to build a biofilm: A fungal perspective. Curr. Opin. Microbiol. 2006, 9, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Ren, H.; Li, Z.; Zhao, T.; Shi, X.; Cheng, H.; Zhuang, W.; Chen, Y.; Ying, H. Enhancement of nuclease P1 production by Penicillium citrinum YL104 immobilized on activated carbon filter sponge. Appl. Microbiol. Biotechnol. 2015, 99, 1145–1153. [Google Scholar] [CrossRef]
- Chen, T.; Liu, N.; Ren, P.; Xi, X.; Yang, L.; Sun, W.; Yu, B.; Ying, H.; Ouyang, P.; Liu, D.; et al. Efficient Biofilm-Based Fermentation Strategies for L-Threonine Production by Escherichia coli. Front. Microbiol. 2019, 10, 1773. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, Q.; Zhou, T.; Li, B.; Yao, S.; Li, A.; Wu, J.; Ying, H. Ethanol production by repeated batch and continuous fermentations by Saccharomyces cerevisiae immobilized in a fibrous bed bioreactor. J. Microbiol. Biotechnol. 2013, 23, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Chen, Y.; Ren, H.; Liu, D.; Zhao, T.; Zhao, N.; Ying, H. Economically enhanced succinic acid fermentation from cassava bagasse hydrolysate using Corynebacterium glutamicum immobilized in porous polyurethane filler. Bioresour. Technol. 2014, 174, 190–197. [Google Scholar] [CrossRef]
- Sun, W.; Yu, Y.; Chen, J.; Yu, B.; Chen, T.; Ying, H.; Zhou, S.; Ouyang, P.; Liu, D.; Chen, Y. Light Signaling Regulates Aspergillus niger Biofilm Formation by Affecting Melanin and Extracellular Polysaccharide Biosynthesis. Mol. Biol. Physiol. 2021, 12, e03434-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Chen, Y.; Li, A.; Ding, F.; Zhou, T.; He, Y.; Li, B.; Niu, H.; Lin, X.; Xie, J.; et al. Enhanced butanol production by modulation of electron flow in Clostridium acetobutylicum B3 immobilized by surface adsorption. Bioresour. Technol. 2013, 129, 321–328. [Google Scholar] [CrossRef]
- Ding, S.; Zhang, D.; Sha, Y.; Wang, F.; Liang, C.; Chen, T.; Sun, W.; Zhuang, W.; Yu, B.; Liu, D.; et al. Efficient preparation of phytase from genetically modified Pichia pastoris in immobilised fermentation biofilms adsorbed on surface-modified cotton fibres. Process Biochem. 2021, 111, 69–78. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, F.; Yu, Y.; Ding, S.; Chen, T.; Sun, W.; Liang, C.; Yu, B.; Ying, H.; Liu, D.; et al. Effect of quorum-sensing molecule 2-phenylethanol and ARO genes on Saccharomyces cerevisiae biofilm. Appl. Microbiol. Biotechnol. 2021, 105, 3635–3648. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Heterologous Protein Expression in Pichia pastoris: Latest Research Progress and Applications. Chembiochem 2018, 19, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.W.; Kim, K.; Chang, Y.J.; Kim, B.; Huh, W.K. The beta-1,3-glucanosyltransferase Gas1 regulates Sir2-mediated rDNA stability in Saccharomyces cerevisiae. Nucleic. Acids Res. 2014, 42, 8486–8499. [Google Scholar] [CrossRef] [Green Version]
- Lipke, P.N.; Mathelié-Guinlet, M.; Viljoen, A.; Dufréne, Y.F. A New Function for Amyloid-Like Interactions: Cross-Beta Aggregates of Adhesins form Cell-to-Cell Bonds. Pathogens 2021, 10, 1013. [Google Scholar] [CrossRef]
- Klis, F.M.; Mol, P.; Hellingwerf, K.; Brul, S. Dynamics of cell wall structure in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2002, 26, 239–256. [Google Scholar] [CrossRef]
- Schwartz, R. Enzymes in Industry Production and Application. Biochem. Educ. 1991, 19, 98. [Google Scholar] [CrossRef]
- Batra, N.; Singh, J.; Joshi, A.; Sobti, R.C. Improved Properties ofBacillus coagulans β-Galactosidase through Immobilization. Eng. Life Sci. 2005, 5, 581–584. [Google Scholar] [CrossRef]
- Matella, N.J.; Dolan, K.D.; Lee, Y.S. Comparison of Galactooligosaccharide Production in Free-Enzyme Ultrafiltration and in Immobilized-Enzyme Systems. J. Food Sci. 2006, 71, C363–C368. [Google Scholar] [CrossRef]
- Katrolia, P.; Zhang, M.; Yan, Q.; Jiang, Z.; Song, C.; Li, L. Characterisation of a thermostable family 42 β-galactosidase (BgalC) family from Thermotoga maritima showing efficient lactose hydrolysis. Food Chem. 2011, 125, 614–621. [Google Scholar] [CrossRef]
- Li, L.; Zhang, M.; Jiang, Z.; Tang, L.; Cong, Q. Characterisation of a thermostable family 42 β-galactosidase from Thermotoga maritima. Food Chem. 2009, 112, 844–850. [Google Scholar] [CrossRef]
- Gao, X.; Wu, J.; Wu, D. Rational design of the beta-galactosidase from Aspergillus oryzae to improve galactooligosaccharide production. Food Chem. 2019, 286, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, P.P.; Ryckaert, S.; Geysens, S.; De Vusser, K.; Callewaert, N.; Contreras, R. Pichia surface display: Display of proteins on the surface of glycoengineered Pichia pastoris strains. Biotechnol. Lett. 2008, 30, 2173–2181. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.-B.; Song, H.-T.; Gupta, N.; Ma, L.-X.; Wu, Z.-B. Cell surface display of functionally active lipases from Yarrowia lipolytica in Pichia pastoris. Protein Expr. Purif. 2007, 56, 35–39. [Google Scholar] [CrossRef]
- Wang, Q.; Li, L.; Chen, M.; Qi, Q.; Wang, P.G. Construction of a novel Pichia pastoris cell-surface display system based on the cell wall protein Pir1. Curr. Microbiol. 2008, 56, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Zhou, Q.; Yan, Y. Efficient gene disruption by posttransformational directed internal homologous recombination in Pichia pastoris. Anal. Biochem. 2019, 576, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zheng, C.; Chen, Y.; Ying, H. FLO Genes Family and Transcription Factor MIG1 Regulate Saccharomyces cerevisiae Biofilm Formation During Immobilized Fermentation. Front. Microbiol. 2018, 9, 1860. [Google Scholar] [CrossRef]
- Liang, C.; Ding, S.; Sun, W.; Liu, L.; Zhao, W.; Zhang, D.; Ying, H.; Liu, D.; Chen, Y. Biofilm-based fermentation: A novel immobilisation strategy for Saccharomyces cerevisiae cell cycle progression during ethanol production. Appl. Microbiol. Biotechnol. 2020, 104, 7495–7505. [Google Scholar] [CrossRef]
- Zhang, L.; Liang, S.; Zhou, X.; Jin, Z.; Jiang, F.; Han, S.; Zheng, S.; Lin, Y. Screening for glycosylphosphatidylinositol-modified cell wall proteins in Pichia pastoris and their recombinant expression on the cell surface. Appl. Environ. Microbiol. 2013, 79, 5519–5526. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yield of GOS (%) | Time-Space Yields of GOS (g/L/h) | |||||||
|---|---|---|---|---|---|---|---|---|
| Lactose | Glucose | Galactose | Transfer Disaccharide | Trisaccharide | Tetrasaccharides | GOS | ||
| Free surface-displayed enzyme | 34.2 ± 0.2 | 10.6 ± 0.3 | 6.6 ± 0.6 | 15 ± 0.2 | 24.6 ± 0.1 | 9 ± 0.2 | 48.6 ± 0.6 | 0.33 a |
| Immobilized surface-displayed enzyme | 33.8 ± 0.2 | 9.8 ± 0.2 | 6 ± 0.3 | 18.1 ± 0.1 | 23.6 ± 0.2 | 8.6 ± 0.1 | 50.3 ± 0.3 | 0.44 b |
| Strains or Plasmids | Relevant Characteristics | Sources |
|---|---|---|
| Strains | ||
| Escherichia coli | DH5α | Stored in our lab |
| GS115 | P. pastoris | Stored in our lab |
| GS115-Δ0226 | P. pastoris with the deletion of PAS_chr1-3_0226 | This study |
| GS115-lacA | lacA comes from Aspergillus oryzae BK03 | This study |
| Δ0226-Pir1p-lacA | Pir1p comes from S. cerevisiae S288c | This study |
| Δ0226-Aga2p-lacA | Aga2p comes from S. cerevisiae S288c | This study |
| Δ0226-Flo1p-lacA | Flo1p comes from S. cerevisiae S288c | This study |
| Plasmids | ||
| pPIC9K | Resistance to Ampicillin | Stored in our lab |
| pPICZαA | Resistance to Bleomycin | Stored in our lab |
| Primer Name | Primer Sequence | Source |
|---|---|---|
| Δ0226-UP-F | CCTAGTGATTCCTGTGATGTATTCACGGCTGCGCAAAACT | This work |
| Δ0226-UP-R | GCTATGGTGTGTGGGGGATCATTTTGATTATCTTTGTGAG | This work |
| Δ0226-DOWN-F | GCTCGAAGGCTTTAATTTGCGCGGTTCACATTAATTAAAG | This work |
| Δ0226-DOWN-R | AAATTTAAAGAGATGCGAAACTTGACAGCTTGAGCGTGAC | This work |
| BleoR-F | CTCACAAAGATAATCAAAATGATCCCCCACACACCATAGC | This work |
| BleoR-R | CTTTAATTAATGTGAACCGCGCAAATTAAAGCCTTCGAGC | This work |
| Pir1p-UP | AAAGAGAGGCTGAAGCTTACGTATATGCTCCAAAGGACCC | This work |
| Pir1p-DOWN | CCAGAACCACCACCACCGAATTCACAGTTGAGCAAATCGA | This work |
| lacA-UP | GCTCAACTGTGAATTCGGTGGTGGTGGTTCTGGTGGTGGTGGA TCTGGTGGTGGAGGTTCTTCTATTAAGCATAGA | This work |
| lacA-DOWN | TTAATTCGCGGCCGCCCTAGGTTACTTATCATCATCATCCTTGTA ATCGTAAGCACCCTTTCTT | This work |
| FS-UP | TACGTAGCCACAGAGGCGTGCTTACCAGCAGGCCAGAGGAAAA | This work |
| FS-DOWN | GAATTCAGAGCTGGTGATTTGTCCTGAAGATGATGATGAC | This work |
| Aga2p-UP | AAAGAGAGGCTGAAGCTTACGTACAGGAACTGACAACTAT | This work |
| Aga2p-DOWN | CCAGAACCACCACCACCGAATTCAAAAACATACTGTGTGT | This work |
| Δ0226-UP100 bp | TGAGACACATTTAACCATCGC | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Wang, S.; Niu, H.; Yang, G.; Wang, S.; Wang, Y.; Zhou, C.; Yu, B.; Yang, P.; Sun, W.; et al. Biofilm-Based Biocatalysis for Galactooligosaccharides Production by the Surface Display of β-Galactosidase in Pichia pastoris. Int. J. Mol. Sci. 2023, 24, 6507. https://doi.org/10.3390/ijms24076507
Chen T, Wang S, Niu H, Yang G, Wang S, Wang Y, Zhou C, Yu B, Yang P, Sun W, et al. Biofilm-Based Biocatalysis for Galactooligosaccharides Production by the Surface Display of β-Galactosidase in Pichia pastoris. International Journal of Molecular Sciences. 2023; 24(7):6507. https://doi.org/10.3390/ijms24076507
Chicago/Turabian StyleChen, Tianpeng, Shimeng Wang, Huanqing Niu, Guanjia Yang, Sinan Wang, Yuqi Wang, Chaowei Zhou, Bin Yu, Pengpeng Yang, Wenjun Sun, and et al. 2023. "Biofilm-Based Biocatalysis for Galactooligosaccharides Production by the Surface Display of β-Galactosidase in Pichia pastoris" International Journal of Molecular Sciences 24, no. 7: 6507. https://doi.org/10.3390/ijms24076507
APA StyleChen, T., Wang, S., Niu, H., Yang, G., Wang, S., Wang, Y., Zhou, C., Yu, B., Yang, P., Sun, W., Liu, D., Ying, H., & Chen, Y. (2023). Biofilm-Based Biocatalysis for Galactooligosaccharides Production by the Surface Display of β-Galactosidase in Pichia pastoris. International Journal of Molecular Sciences, 24(7), 6507. https://doi.org/10.3390/ijms24076507